
Транспорт веществ через биологические мембраны вопросы для подготовки к занятию
| Вид материала | Документы |
СодержаниеСодержание занятия |
- Щих процесс транспорта веществ и ионов через клеточные мембраны, несоизмеримо больше,, 76.65kb.
- Отделения заочного обучения, 25.05kb.
- Вопросы к занятию, 110.87kb.
- А. Л. Кекина г. Ростова Ярославской области Рассмотрена Утверждена На заседании кафедры, 140.75kb.
- Факультет биотехнологии и биотехники кафедра промышленной биотехнологии Реферат, 1153.97kb.
- Роль структуры в динамике протонного переноса через полимерные катионообменные мембраны, 416.5kb.
- 1. Основные положения клеточной теории. Вклад Пуркине, Шванна, Вихрова и др в учение, 679.89kb.
- Строение, свойства и функции наружной плазматической мембраны, 370.82kb.
- Методические указания по изучению курса «Страховое право», 149.11kb.
- Вопросы для подготовки к зачёту, 16.45kb.
ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
Вопросы для подготовки к занятию:
1. Структура, физико-химические свойства и функции биологических мембран.
2. Биофизические механизмы трансмембранного массопереноса. Особенности транспорта через клеточные мембраны гидрофобных и гидрофильных веществ.
3. Свободная диффузия.
4. Облегченная диффузия и ее отличительные свойства.
Роль переносчиков и каналов в пассивном транспорте гидрофильных веществ через биологические мембраны.
5. Активный транспорт веществ. Компоненты и свойства систем активного транспорта.
- Калий-натриевый насос как пример системы активного транспорта.
Литература: Медицинская биофизика, гл.5, §1-3, с.108-123, §6, с.141-152, гл.6,§1,2,с.153-166. Ремизов А.Н., гл.13, §13.1,13.3-13.5.с.228-242. Лекционные записи.
Содержание занятия:
Все клетки окружены мембранами, кроме того, во многих из них (прежде всего в эукариотических) имеется обширная сеть внутренних мембранных систем. Различают наружную клеточную мембрану (плазматическую мембрану или плазмалемму) и внутренние мембраны такие, как ядерная мембрана, мембраны миелиновых оболочек, митохондрий, эндоплазматического ретикулума и т.д. Основные функции, которые выполняют различные мембраны – барьерная, матричная, механическая, биоэлектрическая, рецепторная.
В состав биологических мембран входят в основном белки и фосфолипиды, весовое отношение которых колеблется в разных типах мембран в довольно широких пределах. Фосфолипиды – главная составная часть липидной фракции мембран – представляют собой амфипатические соединения, т.е. соединения, имеющие в своем составе две функционально различные части – полярную гидрофильную головку и гидрофобный хвост. В качестве полярной головки выступает остаток фосфорной кислоты и связанный с ним радикал, который может иметь различную природу. Чаще всего такими радикалами являются холин, этаноламин, серин, треонин, глицерин, инозин и олигосахариды. В качестве гидрофобной части выступают остатки жирных кислот (стеариновой, пальмитиновой, олеиновой, линолевой и др.), которые образуют сложный эфир с глицерином. В водных растворах фосфолипиды объединяются в сложную двухслойную структуру, которая представляет собой одну из разновидностей жидких кристаллов. В таком двойном слое, который является основой любой биомембраны, полярные головки направлены наружу, а хвосты жирных кислот погружены в толщу мембраны. Толщина мембран составляет в среднем величину 7 - 10 нм.
Было предложено несколько моделей строения биомембран, из которых наиболее адекватной считается жидкостно-мозаичная. Согласно этой модели мембрана – это подвижная мозаика, образованная фосфолипидами, в которую погружены белки и в которой они закреплены цитоскелетом. Такие белки называют интегральными мембранными белками. Некоторые из них пронизывают мембрану насквозь, часто многие интегральные белки объединяются в агрегаты. Эта модель предусматривает также, что молекулы фосфолипидов могут достаточно свободно перемещаться в плоскости мембраны. Такое перемещение называется латеральной диффузией. При этом перемещение молекулы из одного слоя в другой весьма редкое событие.
Так как мембрана отделяет внутреннее содержимое клеток от наружной среды, то у нее имеется наружная и внутренняя сторона, и, следовательно, можно говорить о наличии у мембраны асимметрии. С асимметрией строения связано и различие функций, выполняемых белками на внешней и внутренней поверхности мембраны.
Основные функции мембранных белков: во-первых, они выполняют роль структурного компонента, т.е. усиливают прочность липидного каркаса. Во-вторых, имеются белки, образующие так называемые мембранные каналы для транспорта в клетку необходимых веществ, и белки, играющие роль переносчиков. В-третьих, в мембраны встроены белки-ферменты и белки-рецепторы.
Жесткость и проницаемость мембраны зависят от типа жирных кислот, входящих в состав фосфолипидов. Чем больше в мембране ненасыщенных жирных кислот, тем меньше ее жесткость и выше проницаемость. Одним из мембранных липидов является холестерин. Мембраны, содержащие повышенное количество холестерина, отличаются повышенной жесткостью и более низкой проницаемостью.
Понятие транспорта веществ через биомембраны объединяет механизмы поступления в клетку и выделения из клетки различных веществ. При этом различают два основных типа транспорта, а именно пассивный и активный. Пассивным транспортом называют перенос вещества из области с высокой его концентрацией в область с низкой концентрацией. На совершение работы по переносу в этом случае тратится свободная энергия, связанная с неравновесным распределением вещества в системе. Если переносимое вещество имеет электрический заряд, нужно учитывать и потенциальную энергию в электрическом поле.
Движущей силой переноса ионов является обобщенный градиент, являющийся суммой концентрационного и электрического. Для переноса воды через мембраны клеток существенными являются также градиенты осмотического и гидростатического давлений. Результатом пассивного транспорта является выравнивание концентраций. Массоперенос, происходящий в результате суммарного действия осмотического и фильтрационного градиентов, называют конвективным потоком. Это трансмембранный поток воды вместе с растворенными в ней веществами. Конвективный поток обеспечивает обмен веществ в капиллярах между кровью и межклеточной жидкостью, образование первичной мочи и др. К пассивному транспорту относят три основных механизма прохождения веществ через мембраны. Это, во-первых, диффузия жирорастворимых веществ через липидную фазу, во-вторых, диффузия через специальные каналы (поры) и, так называемая, облегченная диффузия - перенос ионов через мембрану специальными молекулами-переносчиками.
Активным транспортом, называется перенос вещества в направлении, противоположном тому, которое определено термодинамическим сопряжением всех градиентов. Этот процесс требует затраты свободой энергии, запасаемой в молекулах АТФ. Системы активного транспорта называют также биологическими насосами.
Диффузией называется взаимное проникновение соприкасающихся веществ друг в друга вследствие теплового движения частиц вещества. Диффузия вещества происходит в направлении уменьшения концентрации и ведет к его равномерному распределению в занимаемом объеме. Величина плотности потока j вещества диффундирующего в направлении оси Х пропорциональна градиенту концентрации dC/dx, это закон Фика: j = -DdC/dx. Плотность потока вещества j = (dm/dt)/S, С – концентрация вещества. Коэффициент пропорциональности D называется коэффициентом диффузии. Для вычисления плотности потока вещества при диффузии через биомембрану используют также уравнение: j = - Dk (Ci – Ce)/l = Dk (Ce – Ci)/l или j = P(Ce – Ci), где Ci и Ce -концентрации частиц вблизи клеточной мембраны внутри и снаружи клетки соответственно, l – толщина мембраны. Отношение P = Dk/l называют коэффициентом проницаемости, k – коэффициент распределения вещества между между липидной и водной фазой.
Через биомембраны идут потоки не только электрически нейтральных веществ, но и ионов, в первую очередь катионов натрия и калия. На процесс переноса заряженных частиц существенное влияние оказывает наличие электрического поля внутри мембраны, обусловленного разностью потенциалов между внутри- и межклеточной средой. Это значит, что перенос ионов определяется в первую очередь двумя градиентами – градиентом концентрации и градиентом потенциала.
При наличии обоих градиентов, процесс диффузии ионов описывается электродиффузионным уравнением Нернста-Планка: j = - DdC/dx – СuzFd/dx, где u – подвижность частиц, z – валентность частиц, F – постоянная Фарадея, φ – электрический потенциал.
Пути и способы прохождения различных веществ через биомембраны зависят от химической природы и размеров молекул. Жирорастворимые вещества способны проникать через билипидный слой, просто растворяясь и диффундируя в нем. Гидрофильные вещества проходят через специальные каналы или с помощью молекул-переносчиков. Каналы представляют собой образованный белками проход в мембране, через который могут проходить гидратированные ионы. Они обладают специфичностью в пропускании ионов. Скорость транспорта вещества по каналу на 5-6 порядков медленнее, чем в результате обычной диффузии того же вещества в водном растворе, но в 100-1000 раз выше, чем при транспорте с переносчиком. Важно отметить, что транспорт ионов через каналы подчиняется уравнениям диффузии и не требует затрат энергии АТФ.
Облегченной диффузией называют разновидность пассивного транспорта веществ через мембраны, который происходит в том же направлении, что и свободная диффузия, но с помощью специальных белков-переносчиков. Облегченная диффузия обеспечивает значительно более высокую скорость переноса вещества, чем свободная диффузия, но в отличие от нее нуждается в энергии АТФ. Эта энергия расходуется на создание и поддержание благоприятных градиентов, при наличии которых, собственно, и возможна облегченная диффузия. Процесс облегченной диффузии описывается уравнением Михаэлиса-Ментен: j
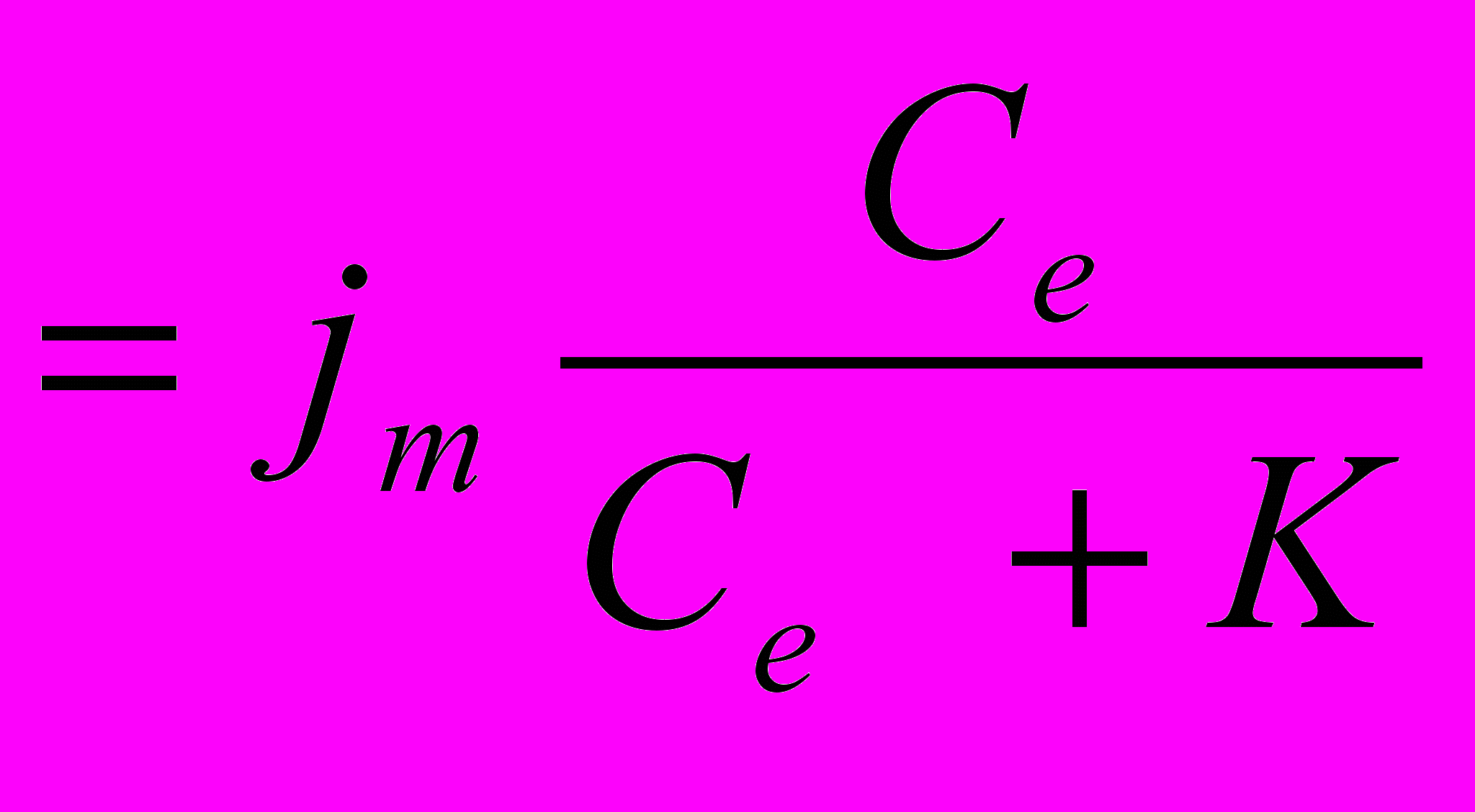 . В этом уравнении j – плотность потока вещества через мембрану, j m – максимальное значение этой величины, Ce – концентрация диффундирующего вещества вне клетки, K – константа, определяющая способность данного вещества проникать в клетку (величина 1/К определяет сродство переносчика к переносимому веществу). Эта константа численно равняется концентрации вещества вне клетки Сe, при которой плотность потока равна половине максимальной, т.е. j = 0,5jm. Из этой формулы следует, что при значениях концентрации вещества вне клетки Сe, значительно меньших величины К (Се << K), плотность потока диффундирующего в клетку вещества j линейно зависит от Се, а при Се >> K она достигает своего максимального значения и не изменяется более с ростом Се.
. В этом уравнении j – плотность потока вещества через мембрану, j m – максимальное значение этой величины, Ce – концентрация диффундирующего вещества вне клетки, K – константа, определяющая способность данного вещества проникать в клетку (величина 1/К определяет сродство переносчика к переносимому веществу). Эта константа численно равняется концентрации вещества вне клетки Сe, при которой плотность потока равна половине максимальной, т.е. j = 0,5jm. Из этой формулы следует, что при значениях концентрации вещества вне клетки Сe, значительно меньших величины К (Се << K), плотность потока диффундирующего в клетку вещества j линейно зависит от Се, а при Се >> K она достигает своего максимального значения и не изменяется более с ростом Се.Перенос вещества против действия физико-химических градиентов является кардинальным отличием активного транспорта от переноса вещества в рассмотренных выше процессах пассивного транспорта. Очевидно, что такой перенос оказывается возможным благодаря термодинамическому сопряжению биологических процессов в клетке. Заметим, что у человека в покое (30 40)% всей энергии, получаемой в ходе метаболических процессов, расходуется на активный транспорт, т.е. на поддержание концентрационного и электрического градиентов на клеточных мембранах. Расход энергии на активный транспорт молей вещества можно оценить через изменение электрохимического потенциала переносимых ионов: W = RTlnC2/C1 + ZFΔφ.
Характерной особенностью активного транспорта является специфичность. Каждая из систем активного транспорта обеспечивает перенос через мембрану только данного вещества (или группы веществ) и не переносит другие. Однако, возможно сопряжение активного транспорта одного вещества с пассивным транспортом другого (Na+ - глюкоза).
Таким образом, основными компонентами систем активного транспорта веществ через мембраны являются: источник энергии – АТФ, специфический переносчик – молекула белка, входящая в состав мембраны, фактор сопряжения работы переносчика с источником энергии (фермент - АТФ-аза).
Известно, что для нормальной жизни клетки необходимо поддержание определенного градиента концентрации калия и натрия между клеткой и внешней средой. А именно, концентрация калия внутри клетки выше, нежели вне ее. При этом концентрация натрия вне клетки выше, чем внутри. Калий-натриевым насосом называют систему, которая постоянно осуществляет транспорт калия в клетку и выведение натрия из нее, т.е. поддерживает концентрационные градиенты этих катионов на клеточной мембране, противодействуя выравниванию их концентраций вследствие пассивного транспорта.
При изменении концентрации ионов калия или натрия в цитоплазме в неблагоприятную сторону (т.е. при возрастании концентрации натрия или понижении концентрации калия) активируется калий-натрий-зависимая АТФ-аза (иначе называемая калий-натриевая-АТФ-аза). Этот фермент содержится в плазмалемме.
Далее, активированная АТФ-аза катализирует отщепление от АТФ фосфатной группы HPO3, которая присоединяется к переносчику, роль которого в данной системе выполняет сама АТФ-аза. Иначе говоря, АТФ-аза фосфорилируется.
Фосфорилированная АТФ-аза приобретает сродство к ионам натрия и связывает три иона Na+ из цитоплазмы. Связывание этих ионов приводит к изменению конформациии фермента и его ориентации в мембране, в результате чего фосфатная группа с ионами натрия оказывается на внешней стороне мембраны. Для отрыва ионов Na+ от переносчика необходима затрата энергии. Эта энергия получается за счет переноса фосфатной группы от АТФ-азы к молекуле воды (HPO3 + H2O = H3PO4). Освободившиеся ионы натрия переходят в межклеточную среду.
Дефосфорилированный переносчик приобретает сродство к ионам калия и связывает два иона К+. Это связывание приводит к изменению конформации переносчика и очередному изменению его ориентации в мембране, в результате чего связанные ферментом ионы калия переносятся на внутреннюю поверхность мембраны, т.е. внутрь клетки. Затем начинается новый цикл.
Таким образом, за один цикл калий-натриевый насос перекачивает три иона Na+ из клетки наружу и два иона К+ извне внутрь клетки. На это затрачивается энергия одной молекулы АТФ. Отметим, что калий-натриевый насос имеет очень высокий коэффициент полезного действия.