
Строение, свойства и функции наружной плазматической мембраны
| Вид материала | Документы |
- Биологические мембраны. Строение, свойства, функции. Ю. А. Владимиров, 157.33kb.
- Урок по теме: «Опорно-двигательная система. Строение, состав и свойства костей», 98.27kb.
- 1. Основные положения клеточной теории. Вклад Пуркине, Шванна, Вихрова и др в учение, 679.89kb.
- Строение и свойства металлических материалов лекция 2 Строение и свойства металлов, 103.5kb.
- Строение, свойства, получение. Цель: изучить строение, свойства и способы получения, 283.21kb.
- Тест по предмету «Физиология цнс» (1 курс дневного отделения), 79.8kb.
- Определители и системы линейных уравнений. Матричная алгебра. Векторная алгебра. Аналитическая, 8.15kb.
- Урок алгебры в 10 классе Тема: «Логарифмы, логарифмическая функция, её свойства и график», 66.6kb.
- Простейшие функции. Квадратные корни, 326.99kb.
- Тема урока: «Фосфор. Строение атома, аллотропия, свойства и применение фосфора», 43.99kb.
1 2
Строение, свойства и функции наружной плазматической мембраны.
Наружная плазматическая мембрана является обязательно составной частью любой клетки. Она служит барьером, который обособляет содержимое клетки от внешней среды и обеспечивает ее целостность. Она обеспечивает взаимодействие клетки с другими клетками организма. Особенности химического состава, строения и функционирования наружной плазматической мембраны создают для клетки возможность существования как единой целостной структуры, открытой системы, обменивающейся с окружающей средой веществом, энергией и информацией. Целостность самой мембраны поддерживается структурными компонентами цитоплазмы клетки.
Основные понятия.
Клеточная мембрана Избирательная проницаемость Экзоцитоз
Липиды Самосборка мембраны Трансцитоз
Липидные слои Пассивный транспорт Ионный канал
Интегральные белки Активный транспорт Осмос
Поверхностные белки Диффузия Микроворсинки
Глицерофосфолипиды Облегченная диффузия Гликокаликс
Межклеточные контакты Эндоцитоз Белок-переносчик
Жидкостно-мозаичная модель Пиноцитоз Адгезия
Асимметрия мембраны Фагоцитоз Холестерин
Опосредованный рецепторами эндоцитоз Липосома Амфифильность
Рецепторный белок Сигнальный путь Лиганд
Амфипатичные молекулы Транспортный белок
- Химический состав, строение и свойства наружной плазматической мембраны.
- Липиды и углеводы мембраны.
Основу любой клеточной мембраны составляет двойной липидный слой. Липидные слои не являются плоскими, они всегда изгибаются, замыкаясь сами на себя, устраняя свободные края и образуя полые структуры – пузырьки, вакуоли, вытянутые мешкообразные структуры и т.п. Эти особенности строения мембраны обусловлены особенностями строения молекул липидов и их взаимодействием с полярными молекулами воды.
Липиды представляют собой обширную группу химических соединений, которая объединяет разные по строению вещества, но обладающие одним общим свойством – гидрофобностью. Характерным примером гидрофобной молекулы является триацилглицерин, или триацилглицерид – производное трехатомного спирта глицерина и трех жирных кислот (рис.1).
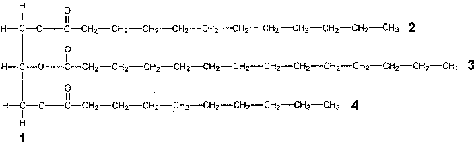
Рис.1 Триацилглицерин.
1 – остаток глицерина
2-4 – остатки трех насыщенных жирных кислот
Молекулы триацилглицерола не растворяются в воде; молекулы воды, взаимодействуя друг с другом многочисленными водородными связями, оттесняют молекулы липидов и те оказываются вместе друг с другом, формируя скопление гидрофобных молекул, окруженное молекулами воды. Создается впечатление, что они взаимодействуют друг с другом особыми – гидрофобными – связями. Таких связей не существует в природе; этим термином обозначают поведение липидов в окружении молекул воды: полярные молекулы воды, взаимодействуя друг с другом, ''подталкивают'' неполярные молекулы друг к другу.
В группу липидов включают следующие химические соединения:
1) Жирные кислоты: насыщенные (олеиновая, пальмитолеиновая) и ненасыщенные, содержащие двойные связи (линолевая, арахидоновая).
2) Мыла - соли жирных кислот
3) Триацилглицерины, или нейтральные жиры (рис.1), которые являются важными компонентами пищи челевека. В состав природных жиров обычно входят жирные кислоты с четным числом атомов углерода.
4) Фосфолипиды – липиды, содержащие не только жирные кислотиы и спирт, но и остаток фосфорной кислоты, с которым часто связаны азотсодержащие соединения и моносахариды.
а) Фосфоглицериды – похожие на триацилглицерины вещества, у которых вместо одной жирной кислоты расположен остаток фосфорной кислоты, связанный с остатком низкомолекулярного химического соединения, таким как этаноламин, холин, серин, инозит и т.п. (рис.2а)
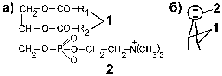
Рис.2. Фосфатидилхолин (лецитин)– один из фосфолипидов мембраны.
1 – гидрофобные остатки жирных кислот
2 – полярная гидрофильная химическая группировка, способная взаимодействовать с полярными молекулами воды.
Обычно одна из жирных кислот фосфоглицеридов является насыщенной, а другая – ненасыщенной, то есть содержащей двойные связи.
б) сфингофосфолипиды – производные многоатомного аминоспирта сфингозина, с которым связаны остатки фосфорной кислоты и жирной кислот, а также остатки холина в сфингомиелине и галактозы или глюкозы - в цереброзиде. Миелин оболочек, окружающих и изолирующих многие нервные волокна, состоит, в основном, из сфингомиелина.
Молекулы фосфолипидов имеют две группы участков: гидрофобные и гидрофильные. Вещества, обладающие одновременно и гидрофобными, и гидрофильными химическими группами, называются амфипатическими, а свойство молекулы иметь в своем составе как гидрофильные, так и гидрофобные группы, называется амфифильностью. При описании строения мембраны фосфоглицериды принято изображать в виде полярной ''головки'' и двух неполярных ''хвостов'' (рис.2б). Полярные ''головки'' фосфоглицеридов взаимодействуют с молекулами воды, а неполярные ''хвосты'' подталкиваются молекулами воды друг к другу. В результате этого фосфолипиды располагаются в два слоя, в которых их полярные головки обращены к водной среде – межклеточной жидкости и цитоплазме, а неполярные хвосты оттесняются в центральную область мембраны (рис.3, 4).
 1
1
 6
6 
 7
7
 3 5
3 5
 2
2 
 4
4 Рис.3. Подвижность липидов наружной плазматической мембраны.
1 – перемещение вдоль слоя
2 – вращение вокруг своей оси
3 – колебательные движения гидрофобных участков
4 – полярные молекулы воды
5 – двойной липидный слой
6 – полярная головка фосфолипида
7 – гидрофобные хвосты фосфолипида
Липиды одного слоя обнаруживают высокую подвижность (рис.3): их гидрофобные участки совершают колебательные движения, а сами фосфолипиды очень быстро вращаются вокруг своей оси и перемещаются вдоль слоя, в котором они находятся. Благодаря этому липидные слои ведут себя как густая маслянистая жидкость, то есть обладают определенной текучестью. Чрезвычайно редко липиды переходят из слоя в слой.
Межклеточная среда
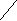 2 2
2 2 








 3 4 5 3 5
3 4 5 3 5 



 1 6
1 6



11
10


 12
12 

 9
9  7 8 13 7 8 14
7 8 13 7 8 14цитоплазма
Рис.4. Схема строения наружной плазматической мембраны животной клетки.
1 – интегральный белок наружного слоя
2 – олигосахарид, ковалентно связанный с белком
3 – периферический белок наружного слоя
4 – интегральный белок, пронизывающий мембрану насквозь
5 – олигосахарид, ковалентно связанный с липидом
6 - полярная головка фосфолипида
7 – периферический белок внутреннего слоя
8 – интегральный белок внутреннего слоя мембраны
9 – гидрофобные хвосты фосфолипида
10 – двойной липидный слой
11 – наружный липидный слой
12 – внутренний липидный слой
13 – холестерин
14 – молекулы воды
5)Гликолипиды – производные сфингозина, с которым связана жирная кислота и разветвленные олигосахариды. В эту группу входит сиаловая кислота и химические соединения, обусловливающие принадлежность человека к той или иной группе крови системы АВО. Гликолипиды всегда располагаются во внешнем слое плазматической мембраны и их углеводная часть находится во внешней для клетки среде (рис.4).
6) Стероиды – производные фенантрена,с которым соединено циклопентановое кольцо. Эта группа включает в себя холестерин (рис.5), желчные кислоты, гормоны коры надпочечников,половые гормоны, витамины группы D и другие важные для организма вещества.
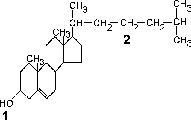
Рис.5. Холестерин.
1 – гидрофильный участок молекулы
2 – гидрофобный участок молекулы
Холестерин является одним из главных компонентов плазматической мембраны (рис.3,6). Гидрофильная группа ОН- холестерина взаимодействует с полярной головкой мембранного фосфолипида , а его гидрофобная углеводородная цепь располагается между гидрофобными хвостами фосфолипидов. Холестерин скрепляет фосфолипидный слой и препятствует плотной упаковке гидрофобных хвостов, повышая тем самым текучесть мембраны.
Рис.6. Взаимодействие холестерина с фосфолипидами мембраны.
1 – гидрофильный участок фосфолипида
2 – остаток ненасыщенной жирной кислоты
3 – гидрофобный участок холестерина
4 – гидрофильный участок холестерина
5 – остаток насыщенной жирной кислоты
Для наружной мембраны всех организмов характерна асимметрия химического состава липидов. Так, гликолипиды всегда расположены во внешнем слое; здесь же расположена большая часть сфингомиелина и фосфатидилхолина. Фосфатидилсерин, несущий суммарный отрицательный заряд, расположен, в основном, во внутреннем слое мембраны; здесь же находится фосфатидилинозитол, участвующий в трансмембранной передачи внешнего сигнала в цитоплазму клетки. Асимметричное расположение разных липидов в каждом слое мембраны очень важно для функции клетки и поддерживается специальными механизмами, в которых участвуют транспортные белки и ферменты.
Двойной липидный слой подобен двумерной жидкости, в которой молекулы липидов перемещаются в пределах своего слоя. Липидный бислой обычно устраняет свободные края, замыкаясь сам на себя. По этой причине двойные липидные слои способны спонтанно формироваться, самопроизвольно восстанавливаться при повреждениях, сливаться при тесном контакте. Благодаря такой способности липидных слоев происходит слияние клеток, образование и слияние транспортных пузырьков при эндо- и пиноцитозе.
Важнейшим свойством липидного бислоя является текучесть, вязкость. Текучесть двойного липидного слоя характеризуется следующим образом:
а) текучесть повышается при увеличении числа двойных связей в жирных кислотах мембранных липидов: в месте расположения двойной связи жирная кислота формирует излом, что препятствует плотной упаковке молекул
б) текучесть возрастает при уменьшении числа углеводородных групп (СН2) в жирных кислотах
в) при увеличении количества холестерина текучесть сначала уменьшается, а затем – увеличивается
г) текучесть меньше в гидрофильной области липидного бислоя и больше – в гидрофобной области
д) текучессть увеличивается при повышении температуры.
Липидные слои обладают достаточной прочностью и гибкостью, что особенно важно при изменении формы и при движении клеток, при делении тела клетки во время митоза и мейоза, при формировании многих межклеточных контактов.
Липиды встраиваются в мембрану определенным образом, объединяются в группы, формируя участки с различными свойствами и функциональными возможностями. Такие области могут возникать только в одном из двух слоев липидного бислоя. Поэтому мембрана содержит не однородный липидный бислой, а представляет собой своеобразную мозаику из областей разного состава липидов с разными свойствами, позволяющим им выполнять разные функции.
Липиды мембраны служат средой, в которой расположены и функционируют мембранные белки. Липиды мембраны участвуют в регуляции активности мембранных белков.
Липидные слои мембраны практически непроницаемы для сильно полярных молекул, которых много в цитоплазме. Это позволяет липидному бислою осуществлять свою главную функцию – служить барьером, препятствующим утечке компонентов цитоплазмы.
Некоторые мембранные липиды участвуют в передаче сигналов: фосфатидилинозитол, эйкозаноиды – производные всех жирных кислот, содержащих 20 атомов углерода ( простагландины и лейкотриены)
Эйкозаноиды участвуют в болевых и температурных реакциях, регуляции артериального давления, сокращения матки при родах, регуляции цикла сна и бодрствования, в воспалительных реакциях, в том числе при таких патологических процессах как ревматоидный артрит и псориаз. Существуют заболевания, обусловленные нарушением образования мембранных липидов. Например, при болезни Тея-Сакса и Гоше нарушается обмен гликосфинголипидов и накопление продуктов их частичного расщепления вызывает тяжелое поражение нервной системы.
2.2. Белки мембран.
Большая часть мембранных белков погружена в липидный бислой почти перпендикулярно и расположена в ней мозаично (рис.4). В мембране различают два типа белков:
а) периферические белки
б) интегральные белки.
Периферические белки расположены на поверхности мембраны и связаны с ней в основном ионными связями. Эти белки легко отделяются от мембраны. У большинства клеток, за исключением клеток крови, снаружи расположен белок фибронектин. На внутренней поверхности большинства клеточных мембран, особенно в эритроцитах, расположен белок спектрин.
Интегральные белки или погружены в толщу липидного бислоя, или пронизывают его насквозь (трансмембранные белки). Все интегральные белки можно выделить из мембраны, только разрушив ее. Интегральные белки амфипатичны: в их молекулах четко выделяются гидрофобные и гидрофильные участки. Гидрофобные участки расположены в толще мембраны, а гидрофильные – обычно на внешней и внутренней поверхностях мембраны. Расположение мембранных белков соответствует максимально эффективному их взаимодействию с липидныи бислоем. Участки белков, расположенные в гидрофобной среде внутренней части мембраны, имеют вид -спирали. Обычно N-конец белков находится во внешней для клетке среде, а С-конец – в цитоплазме (рис.7). Многие интегральные белки перемещаются в плоскости мембраны. Это имеет место при группировке рецепторов, при эндоцитозе.
 1
1

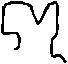
 2
2 NH3+-

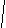 3
3
4
-COO- 5
Рис.7. Расположение участков интегрального белка в мембране.
1 - углевод
2 - N-концевой гидрофильный участок белка
3 –липидный бислой
4 – внутримембранный гидрофобный участок белка в виде -спирали
5 –С- концевой гидрофильный участок белка
Многие трансмембранные белки только один раз пересекают мембрану (рис.7). Многие из них являются рецепторами. Они обычно имеют три участка:
а) наружный, или внеклеточный участок, который узнает сигнальную молекулу, например, молекулу гормона;
б) внутримембранный участок;
в) цитоплазматический, или внутриклеточный, участок, который содержит каталитический центр; активация этого участка сопровождается его фосфорилированием.
Многие трансмембранные белки несколько раз пересекают мембрану. Большинство из них относится к рецепторам, участвующим в передаче сигнала с поверхности клетки внутрь клетки. Многие из них 7 раз пересекают мембрану. К таким белкам относятся 2-адреноэргический рецептор, родопсин палочек сетчатки глаза. Важная особенность этих белков – использование специальных G-белков для трансмембранной передачи сигнала. G-белки используют в качестве источника энергии ГТФ.
Часть периферических и интегральных белков присутствуют в мембранах всех клеток и являются компонентом мембранного цитоскелета.
В многоклеточном организме разные стороны одной и той же клетки различаются по составу мембранных белков. Клетки разных тканей также различаются по составу мембранных белков.
Для мембраны характерна асимметрия расположения белков: одни белки расположены в наружном слое, а другие во внутреннем. Функциональный свойства мембран обусловлены белками, входящими в ее состав. По биологической роли мембранные белки разделяют на три группы:
а) ферменты
б) рецепторные белки – трансмембранные белки, которые способны специфически связываться с информационными молекулами и передавать сигналы внутрь клетки
в) структурные белки.
Некоторые рецепторные белки и ферменты осуществляют перенос ионов, химических соединений или осуществляют трансмембранный перенос сигнала, приходящего к клетке.
Со многими белками ковалентно связаны небольшие разветвленные олигосахариды, участвующие в молекулярном узнавании и межклеточных взаимодействиях. Олигосахариды, связанные с липидами и белками, являются основным компонентом примембранной области клетки, называемой гликокаликсом.
Со стороны цитоплазмы расположена другая примембранная область – кортикальный слой, содержащий фибриллярные элементы цитоплазмы – микрофиламенты и микротрубочки, а также вспомогательные белки, которые связывают эти структуры с наружной мембраной.
Высокая подвижность липидного бислоя и мозаичность расположения в нем белков дали название модели, хорошо описывающей строение, свойства и функции мембраны – жидкостно-мозаичная модель (рис.8).
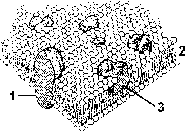
Рис.8. Мозаичное расположение белков в липидном бислое мембраны.
1 – интегральный белок, пронизывающий мембрану
2 – липидный бислой
3 – интегральный белок наружного слоя
Липиды, белки и олигосахариды наружной мембраны придают ей ряд важных свойств:
- эластичность
- гибкость
- упругость
- способность к самосборке и росту
- избирательная проницаемость.
- асимметричность химического состава
- полярность мембраны: имеются разноименные электрические заряды на разных сторонах мембраны
- способность к молекулярному узнаванию химических соединений.