
На правах рукописи Болдырева Лидия Валерьевна Влияние гена
| Вид материала | Автореферат |
- На правах рукописи, 848.13kb.
- На правах рукописи тамбовцев константин Александрович влияние синтетических феромонов, 1060.64kb.
- Печатная или на правах рукописи, 21.09kb.
- Чистякова Елена Андреевна Открытый урок, 128.48kb.
- Удк 796/799: 378 , 770.24kb.
- Золотухина лидия Валерьевна особенности психического здоровья врачей-психиатров с синдромом, 309.24kb.
- 1. 1 Природа генів, 282.45kb.
- Вопросы к экзамену по курсу «Молекулярная биология» (вечернее отделение), 36.64kb.
- На правах рукописи, 726.26kb.
- На правах рукописи, 1025.8kb.
На правах рукописи
Болдырева
Лидия Валерьевна
Влияние гена SuUR на эффект положения
и локализацию белков гетерохроматина
в политенных хромосомах Drosophila melanogaster
Генетика – 03.00.15
АВТОРЕФЕРАТ
ДИССЕРТАЦИИ НА СОИСКАНИЕ УЧЕНОЙ СТЕПЕНИ
КАНДИДАТА БИОЛОГИЧЕСКИХ НАУК
Новосибирск – 2007
Работа выполнена в лаборатории молекулярной цитогенетики Института цитологии и генетики СО РАН, г. Новосибирск
Научный руководитель: доктор биологических наук,
профессор Беляева Елена Сергеевна
Институт цитологии и генетики
СО РАН, г. Новосибирск
Официальные оппоненты: доктор биологических наук,
профессор Высоцкая Людмила Васильевна
Новосибирский государственный
университет, г. Новосибирск
доктор биологических наук,
Колесников Николай Николаевич
Институт цитологии и генетики
СО РАН, г. Новосибирск
Ведущее учреждение: кафедра генетики,
Томский государственный
университет, г. Томск
Защита состоится «____» ___________2007г. на утреннем заседании диссертационного совета по защите диссертаций на соискание ученой степени доктора наук (Д-003.011.01) в Институте цитологии и генетики СО РАН в конференц-зале Института по адресу : 630090, г. Новосибирск, проспект академика Лаврентьева, 10. e-mail: dissov@bionet.nsc.ru, т/ф (383)3331278.
С диссертацией можно ознакомиться в библиотеке Института цитологии и генетики СО РАН.
Автореферат разослан «___» __________ 2007г.
Ученый секретарь
диссертационного совета
Доктор биологических наук А.Д.Груздев
ВВЕДЕНИЕ
Актуальность проблемы
Проблема регуляции активности генов – одна из важнейших в современной биологии. В настоящее время в центре исследований этого направления стоит феномен глубокой инактивации генов (silencing, сайленсинг - молчание). Это состояние устанавливается в раннем онтогенезе и затем наследуется в многочисленных циклах деления клетки. Эпигенетическая репрессия генной активности осуществляется мультимерными белковыми комплексами. ДНК-специфичное связывание репрессоров транскрипции с промоторными областями генов инициирует сборку комплексов, модифицирующих гистоны. Определенные комбинации модификаций гистонов, в свою очередь, обеспечивают установление и поддержание репрессированного состояния (Craig, 2005; Hediger, Gasser, 2006; Richards, Elgin, 2002; Ringrose, Paro, 2004).
Общими свойствами молчащих районов являются репрессия транскрипции, плотная упаковка и поздняя репликация ДНК в S-фазе клеточного цикла (Schubeler et al., 2002, обзор Zhimulev, Belyaeva, 2003). В политенных хромосомах слюнных желез D. melanogaster к молчащим районам относится область хромоцентра, содержащая прицентромерный гетерохроматин, и многочисленные районы интеркалярного гетерохроматина, дискретно распределенные по плечам хромосом. Прицентромерный гетерохроматин состоит, в основном, из повторенной ДНК - он насыщен мобильными элементами и сателлитными повторами. Кроме того, он содержит гены, для правильной экспрессии которых необходимо гетерохроматиновое окружение (Yasuhara, Wakimoto, 2006). В районах интеркалярного гетерохроматина лежат кластеры уникальных генов, имеющие сходные профили репликации и транскрипции (Belyakin et al., 2005).
У Drosophila на данный момент хорошо описаны два белковых комплекса, обеспечивающих репрессию. Один из них включает в себя белок HP1 (Heterochromatin protein 1) и гистон-метилтрансферазу SU(VAR)3-9 (Schotta et al., 2002). Этот комплекс собирается в прицентромерном гетерохроматине, репрессированное состояние которого может распространяться на перемещённые к нему эухроматиновые гены при эффекте положения мозаичного типа (ЭПМ). Другой белковый комплекс, в сборке которого основная роль принадлежит белкам группы PC-G (Polycomb Group), осуществляет репрессию эухроматиновых генов. Несмотря на значительные различия этих комплексов, механизмы их действия объединяет ряд общих черт, главной из которых является метилирование гистонов. Состав и функции отдельных компонентов репрессирующих комплексов служат предметом интенсивного изучения.
Белок SUUR, продукт гена SuUR (Suppressor of Underreplication), является уникальным гетерохроматиновым компонентом, так как он присутствует в обоих типах районов – интеркалярном и прицентромерном гетерохроматине. Фенотип мутации SuUR - подавление недорепликации в гетерохроматиновых районах политенных хромосом – обусловлен тем, что все районы интеркалярного гетерохроматина и некоторые прицентромерные районы заканчивают репликацию существенно раньше, чем в норме. Механизмы влияния гена SuUR на репликацию ДНК пока неизвестны. Он может действовать на уровне упаковки хроматина, делая его менее доступным для репликативной машины, однако, возможно также влияние на саму систему репликации.
ЭПМ возникает вследствие распространения репрессированного состояния гетерохроматина на перемещенные к нему гены. Поэтому об участии того или иного фактора в репрессии гетерохроматина судят, прежде всего, по его влиянию на степень проявления эффекта положения. Так, гены, кодирующие структурные белки гетерохроматина – НР1 и SU(VAR)3-9, SU(VAR)3-7, являются сильнейшими модификаторами эффекта положения.
Цель и задачи исследования
Природа факторов, определяющих локализацию белка SUUR в гетерохроматиновых районах, до настоящего времени не ясна. Неизвестно также, как ген SuUR влияет на организацию гетерохроматиновых доменов и взаимодействует ли белок SUUR с другими структурными белками гетерохроматина, такими как PС и HP1.
Целью настоящей работы было получение ответов на эти вопросы, а именно:
- Выяснить, является ли ген SuUR модификатором эпигенетического сайленсинга, возникающего при мозаичном эффекте положения гена.
- Определить взаимоотношения SUUR с другими хорошо известными белками гетерохроматина.
Для выяснения этих вопросов необходимо было выполнить следующие конкретные исследования:
- Изучить влияние мутации гена SuUR, дополнительных доз гена SuUR и его оверэкспрессии на ЭПМ в ряде эу-гетерохроматиновых перестроек.
- Выяснить, обнаруживается ли белок SUUR в районах хромосом, подверженных ЭПМ.
- Изучить влияние гена SuUR на политенизацию ДНК в районах хромосом, подверженных эффекту положения.
- Проверить взаимозависимость локализации белков SUUR, HP1, SU(VAR)3-7, НР2 и PC на политенных хромосомах методом непрямой иммунофлуоресценции:
а) изучить влияние мутаций генов, кодирующих SUUR и HP1, на связь гетерохроматиновых белков с хромосомами.
б) проверить, как влияет оверэкспрессия этих генов на их локализацию в хромосомах.
Научная новизна
Впервые выявлена функция гена SuUR как супрессора мозаичной экспрессии и недорепликации генетического материала при эффекте положения. Впервые показано, что белок SUUR появляется в эухроматиновых участках политенных хромосом, гетерохроматизированных в результате эффекта положения. Открыта взаимозависимость хромосомной локализации белка SUUR с белком HP1 – важнейшим компонентом респрессированных гетерохроматиновых доменов. Показано, что изменение в локализации и количестве этих белков приводят к сложной цепи нарушений в распределении белковых компонентов гетерохроматина.
Практическая ценность
Обнаруженное в работе сложное взаимодействие между гетерохроматиновыми белками необходимо учитывать во всех исследованиях, где используются линии, мутантные по генам, кодирующим данные белки, и оверэкспрессия этих генов. В особенности, это важно для работы с модификаторами эффекта положения и интерпретации результатов при изучении репликации ДНК.
Апробация работы
Результаты работы представлены на международных конференциях в докладах и стендовых сообщениях: на 5-ой международной конференции по гетерохроматину (Кортона, Италия, 2001), XIV Всероссийском симпозиуме "Структура и функции клеточного ядра" (Санкт-Петербург, 2002), III съезде ВОГиС «Генетика в XXI веке: современное состояние и перспективы развития» (Москва, 2004), 7-ой международной конференции по гетерохроматину (Губбио, Италия, 2005), 8-ой международной конференции по гетерохроматину (Губбио, Италия, 2007) и др.
Объем работы
Диссертация состоит из введения, обзора литературы, описания материалов и методов, результатов и обсуждения, а также выводов и списка цитируемой литературы, в который входит 217 ссылок. Работа изложена на 124 страницах машинописного текста, содержит 13 таблиц и 18 рисунков.
Публикации
По теме диссертации опубликовано 6 работ.
Вклад автора
Основные результаты получены автором самостоятельно. Работа по выявлению домена SUUR, необходимого для перераспределения белка НР1, выполнена совместно с Т.Д. Колесниковой. Картирование сайтов локализации белка SUUR на политенных хромосомах линии дикого типа Oregon-R проведено И.Ф. Жимулевым. Саузерн гибридизация фрагментов из интеркалярного гетерохроматина района 89Е выполнялась совместно с С.А. Демаковым.
Благодарности
Автор выражает глубокую признательность д.б.н. Е. С. Беляевой, академику РАН И.Ф. Жимулеву, к.б.н. Д. Е. Корякову, к.б.н В.В. Шломе, к.б.н. Т.Д. Колесниковой, А.В. Пиндюрину за помощь в работе и обсуждение результатов. Автор также благодарен всем сотрудникам отдела молекулярной цитогенетики за постоянную поддержку.
МАТЕРИАЛЫ И МЕТОДЫ
Линии D. melanogaster описаны в справочнике (Lindsley, Zimm, 1992), базе данных FlyBase (ссылка скрыта), а также, в статьях (Belyaeva et al., 1998, Belyaeva et al., 2003, Makunin et al. 2002; Zhimulev et al. 2003, Kolesnikova et al., 2005).
Экстракцию красного глазного пигмента мух проводили по методу Ephrussi and Herold (1944) с некоторыми модификациями, описанными в статье (Belyaeva et al., 2003).
Подсчет доли щетинок yellow, scute и Stubble вели по методу, описанному в статье (Belyaeva et al., 2003).
Статистическую обработку результатов проводили по схеме однофакторного дисперсионного комплекса с использованием критерия Фишера для анализа альтернативных (качественных) признаков (Васильева, 1999).
Непрямое иимунофлуоресцентное окрашивание политенных хромосом проводили по методике описанной в статье (Belyaeva et al., 2003).
Антитела Для детекции белка SUUR и его фрагментов использовали антитела Е45 против центральной части белка SUUR (Makunin et al. 2002). Антитела С1А9 против белка НР1 (НР1а) и антитела против белка НР2 были любезно предоставлены С. Элгин (США), против белка РС В. Пирротой (Швейцария), против белка SU(VAR)3-7 П. Спирером (Швейцария). В качестве вторых антител использовали коньюгаты иммуноглобулина G: полученные в козле против антигена кролика c флюорохромом FITC и полученные в козле против антигена кролика c родамином (Sigma).
Вестерн-блот анализ проводили согласно стандартной методике (Sambrook, Fritsch, Maniatis, 2004). Белковые экстракты получали из слюнных желез 4-х часовых предкуколок.
Количественный Саузерн-блот анализ проводили по методу, описанному в статье (Belyaeva et al., 2003).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Влияние гена SuUR на ЭПМ
Целью первой части работы было охарактеризовать ген SuUR как возможный модификатор ЭПМ. Исследовали влияние мутации SuUR на репрессию гена white+ в инверсии In(1)wm4h, гена yellow+ в дупликации Dp(1;f)1187, мутации Stubble в транслокации T(2;3)SbV и гена scute+ в инверсии In(1)sc8.
При введении мутации SuUR в линию, несущую In(1)wm4h, мы обнаружили супрессирующее действие мутации SuUR на ЭПМ гена white. Измерение величины оптической плотности (OD480) экстрактов пигмента глаз у мух изогенных линий wm4h;ru,h,SuUR+ и wm4h;ru,h,SuUR- подтвердило повышение количества пигмента глаз у мутантов SuUR-(Таблица 1).
Таблица 1. Влияние мутации гена SuUR на ЭПМ гена white+ в изогенных линиях wm4h;ru,h, SuUR+ и wm4h;ru,h,SuUR-
| | OD480 | |
| Генотип | ♀ | ♂ |
| wm4h;ru,h,SuUR + | 0,092 ± 0,005 | 0,049 ± 0,005 |
| wm4h;ru,h,SuUR - | 0,138 ± 0,005 | 0,068 ± 0,006 |
р0,99 р0,95
Полученные данные позволяют заключить, что мутация SuUR является модификатором ЭПМ. Однако влияние мутации SuUR на ЭПМ в перестройке In(I)wm4h оказалось, хотя и достоверным, но слабо выраженным.
Для изучения влияния мутации гена SuUR на ЭПМ гена yellow+ в "минихромосоме" Dp(1;f)1187 получили потомков от скрещиваний самок y,f; SuUR- с самцами y;Dp(1;f)1187,y+;SuUR+ и контрольного скрещивания самок y,f; SuUR+ с самцами y;Dp(1;f)1187,y+;SuUR+. Полученные потомки были гетерозиготами по мутации SuUR, либо гомозиготами по нормальному аллелю гена SuUR и несли Dp(1;f)1187. Для определения степени модификации ЭПМ у гомозигот по мутации гена SuUR была синтезирована линия y,w;Dp(1;f)1187,y+;SuUR-. Подсчитывали долю желтых щетинок, расположенных по краю крыла (Таблица 2).
Таблица 2. Влияние мутации гена SuUR на ЭПМ гена yellow+ в «минихромосоме» Dp(1;f)1187
| | Частоты щетинок yellow,% | |||
| | 25оC | 18oC | ||
| Генотип F1 | ♀ | ♂ | ♀ | ♂ |
| y; Dp(1;f)1187, y+;SuUR + | 4,7 | 4,4 | 8,3 | 8,4 |
| y; Dp(1;f)1187, y+;SuUR-/ SuUR + | 1,3 | 2,1 | 1,1 | 1,3 |
| Линия y; Dp(1;f)1187, y+;SuUR- | 0,44 | 0,46 | -- | -- |
| y; Dp(1;f)1187, y+; ru,h,SuUR+/ SuUR+ | -- | -- | -- | 9,0 |
| y; Dp(1;f)1187, y+; ru,h,SuUR-/SuUR + | -- | -- | -- | 1,3 |
р0,99 р0,99 р0,99 р0,99
У гомозигот по мутации SuUR мы наблюдали уменьшение числа жёлтых щетинок, что говорило об ослаблении ЭПМ по сравнению с контрольными классами. При температуре 18ºС, усиливающей ЭПМ гена yellow+, супрессирующее влияние мутации SuUR на ЭПМ становилось более заметным. Мы показали эффект мутации SuUR не только в гомозиготе, но и в гетерозиготе, что позволяет сделать заключение о полудоминантности эффекта SuUR.
Аналогичные результаты получены для супрессии ЭПМ гена scute в инверсии In(1)sc8.
В целом, полученные данные характеризуют ген SuUR как гапло-супрессор ЭПМ. Многие гены гапло-супрессоры ЭПМ проявляют себя как энхансеры ЭПМ в дополнительных дозах, поэтому следующей задачей было изучить влияние дополнительных доз и оверэкспрессии SuUR на ЭПМ.
Для оценки степени влияния на ЭПМ оверэкспрессии SuUR мы использовали линию, содержащую транспозон H7, несущий полноразмерный ген SuUR под контролем промотора теплового шока (Makunin et al., 2002). Измерили оптическую плотность экстрактов глазного пигмента потомков от скрещивания самок y,w;H7-X;H7-3 с самцами wm4h;ru,h,SuUR+. Оверэкспрессия SuUR достигалась с помощью серийного теплового шока. Для контрольных измерений использовали потомков этого же скрещивания, но прошедших всё развитие при температуре 25oC. Как следует из данных в Таблице 3 оверэкспрессия SuUR усиливает ЭПМ гена white: количество красного пигмента уменьшается.
Таблица 3. Влияние оверэкспрессии SuUR на ЭПМ гена white+ в In(1)wm4h
-
Температура развития
OD480
Генотип F1
♀
y,w,H7-X / wm4h;H7-3/ru,h,SuUR+
25oC
0,388 ± 0,020
тепловой шок
каждые 12 часов
0,138 ± 0,010
р0,99
Усиление репрессии генов при ЭПМ наблюдалось и при введении дополнительных доз гена SuUR в линии с дупликацией Dp(1;f)1187
Полученные данные позволяют заключить, что ген SuUR является модификатором ЭПМ и его влияние на ЭПМ зависит от дозы гена.
Корреляция локализации белка SUUR с изменением морфологической структуры района, подверженного ЭПМ. Поскольку для белка SUUR характерна локализация в гетерохроматиновых районах, мы хотели выяснить, появляется ли белок SUUR в районах, претерпевающих гетерохроматизацию при ЭПМ. Для этого мы использовали перестройку Dp(1;1)pn2b, которая наилучшим образом демонстрирует различную степень гетерохроматизации района, подверженного ЭПМ (Belyaeva, Zhimulev, 1991).
На цитологических препаратах степень компактизации исследуемого района варьировала от слабой, когда компактизация касалась только одного диска, до гетерохроматизации всего района (Рисунок 1).
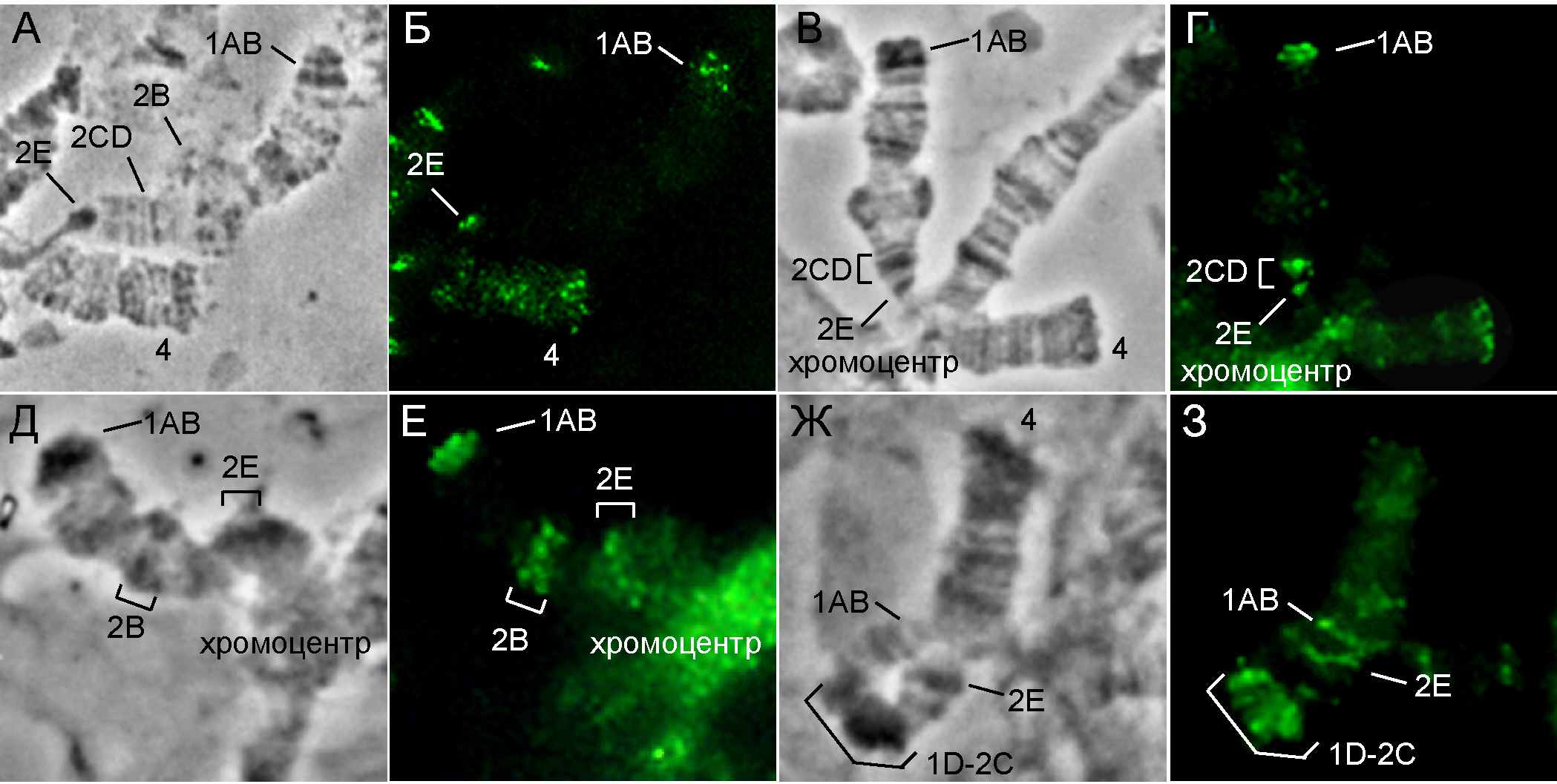
Рис.1. Локализация белка SUUR на политенных хромосомах слюнных желез линии Dp(1;1)pn2b. А,Б – район дупликации без признаков гетерохроматизации; В,Г – компактизация района 2CD; Д,Е – компактизация района 2В; Ж,З – гетерохроматизация участка 1D-2C. Район компактизации отмечен скобкой. А, В,Д,Ж – фазовый контраст; Б,Г,Е,З – иммунолокализация белка SUUR – белый сигнал на черном фоне.
Полученные результаты показали, что SUUR появляется в районах, подверженных гетерохроматизации при ЭПМ, так как его присутствие совпадает с паттерном компактизации эухроматиновых районов.
Влияние мутации SuUR на степень политенизации последовательностей в районах, подверженных ЭПМ. Известно, что эухроматиновые районы политенных хромосом, подверженные ЭПМ, в ряде случаев становятся недореплицированными. Мы оценили степень влияния мутации SuUR на политенизацию в области гена white в In(1)wm4h и в области гена scute в In(1)sc8.
Последовательность w1-4 из гена white полностью политенизируется у личинок дикого типа Oregon-R. В инверсии In(1)ww4h степень её политенизации составляет менее 5% по сравнению с диким типом. У гомозиготных мутантов SuUR степень представленности фрагмента w1-4 в перестройке In(1)ww4h увеличивается до 69% (Рисунок 2,А).
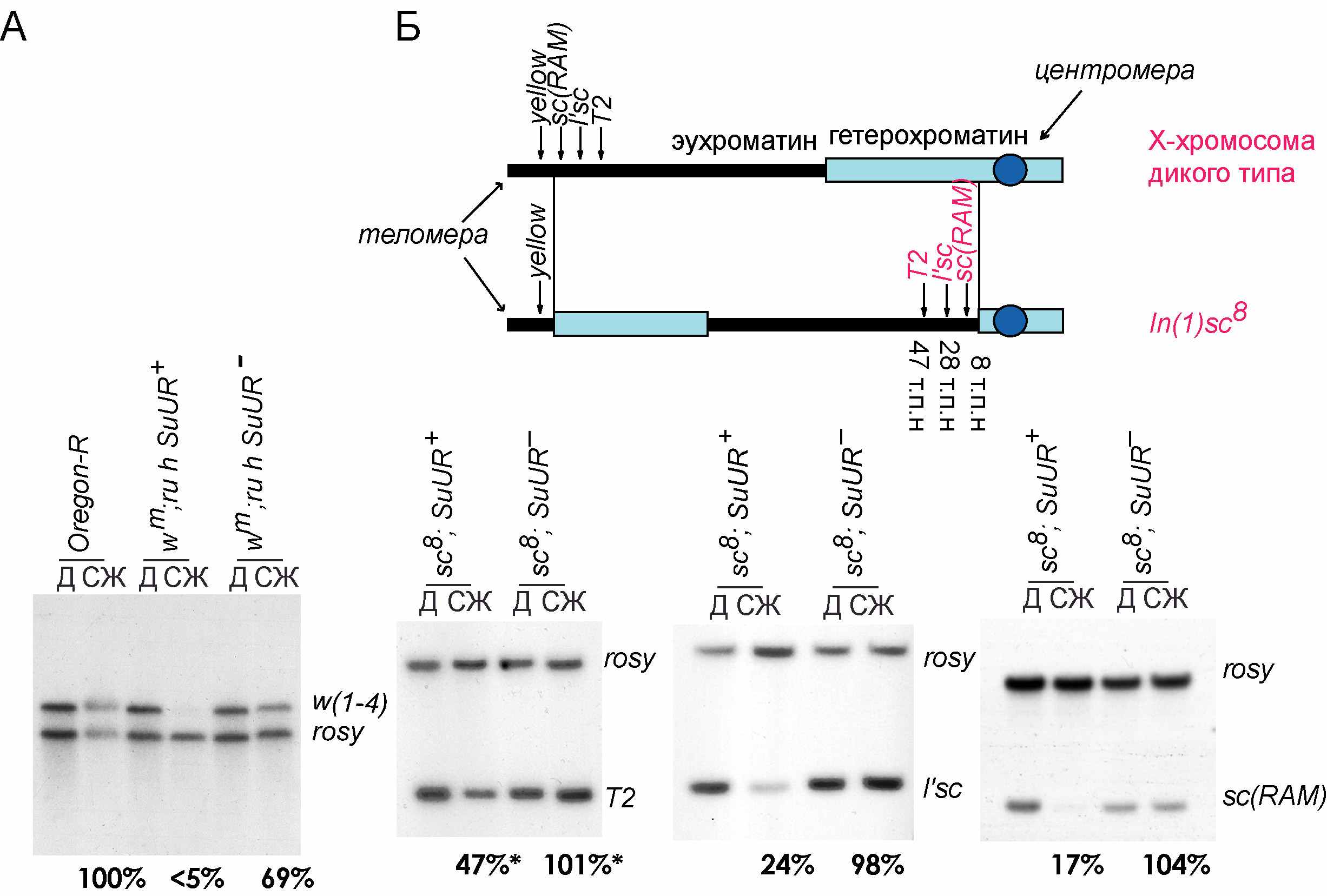 –
–Рис. 2. А. Саузерн гибридизация зонда w 1-4 с геномной ДНК линии Oregon-R и изогенных линий wm4h;ru,h,SuUR+ и wm4h;ru,h,SuUR-. Б. Саузерн гибридизация зондов T2, l'sc, sc(RAM) с геномной ДНК линий sc8;SuUR+ и sc8;SuUR-. Д – диплоидная ткань (нейробласты). СЖ – слюнные железы. Интенсивность сигналов исследуемых зондов соотнесена с таковой полностью политенизированного фрагмента rosy.
Мы также определили степень представленности последовательностей, лежащих на разном расстоянии от точки разрыва в инверсии In(1)sc8 в линиях sc8;SuUR+ и sc8;SuUR-. Самая ближайшая к точке разрыва последовательность sc(RAM) в слюнных железах (8 т.п.н. от точки разрыва) политенизируется всего на 17% (Рисунок 2, Б).
Фрагмент l’sc, гомологичный участку, лежащему в 28 т.п.н. от точки разрыва, политенизируется на 24%. Даже на расстоянии 50 т.п.н. от точки разрыва In(1)sc8 политенизация участка, соответствующего фрагменту Т2, составила всего 47%. При введении мутации SuUR в линию все три последовательности становятся полностью представленными. Таким образом, мутация SuUR приводит к полной супрессии недорепликации в In(1)sc8 (Рисунок 2, Б).
Можно полагать, что функции белка SUUR при ЭПМ связаны с его влиянием на процесс репликации в гетерохроматиновых районах. Кроме того, влияние гена SuUR на ЭПМ может быть обусловлено его взаимодействием с продуктами генов, кодирующих другие белки гетерохроматина.
Локализация SUUR и гетрохроматинового белка HP1 взаимозависима. Белок НР1 играет центральную роль в формировании гетерохроматина. В диких линиях он локализован в хромоцентре и немногочисленных сайтах в плечах хромосом (Рисунок 3, А). Белок SUUR в норме колокализуется с НР1 в хромоцентре и обнаруживается в многочисленных сайтах интеркалярного гетерохроматина (Рисунок 3, Б). Чтобы определить, зависит ли способность белка SUUR связываться с хромосомами от белка НР1, провели иммуноокрашивание препаратов политенных хромосом трансгетерозигот Su(var)20503/Su(var)20505 антителами к белку SUUR. У таких мутантов белок HP1 (продукт гена Su(var)205) не выявляется на хромосомах у личинок третьего возраста. Оказалось, что при этом полностью отсутствует и сигнал локализации белка SUUR (Рисунок 3, В).
Это говорит о зависимости связывания SUUR от HP1. При оверэкспрессии HP1, индуцированной тепловым шоком в линии hs-HP1, HP1 связывается с множеством районов (Рисунок 3, Г). При этом сигнал белка SUUR целиком исчезает с хромосом как из районов интеркалярного гетерохроматина, так и из хромоцентра (Рисунок 3, Д). Однако, при одновременной оверэкспрессии HP1 и SUUR тепловым шоком картина связывания SUUR с хромосомами восстанавливается (см. Таблицу 4).
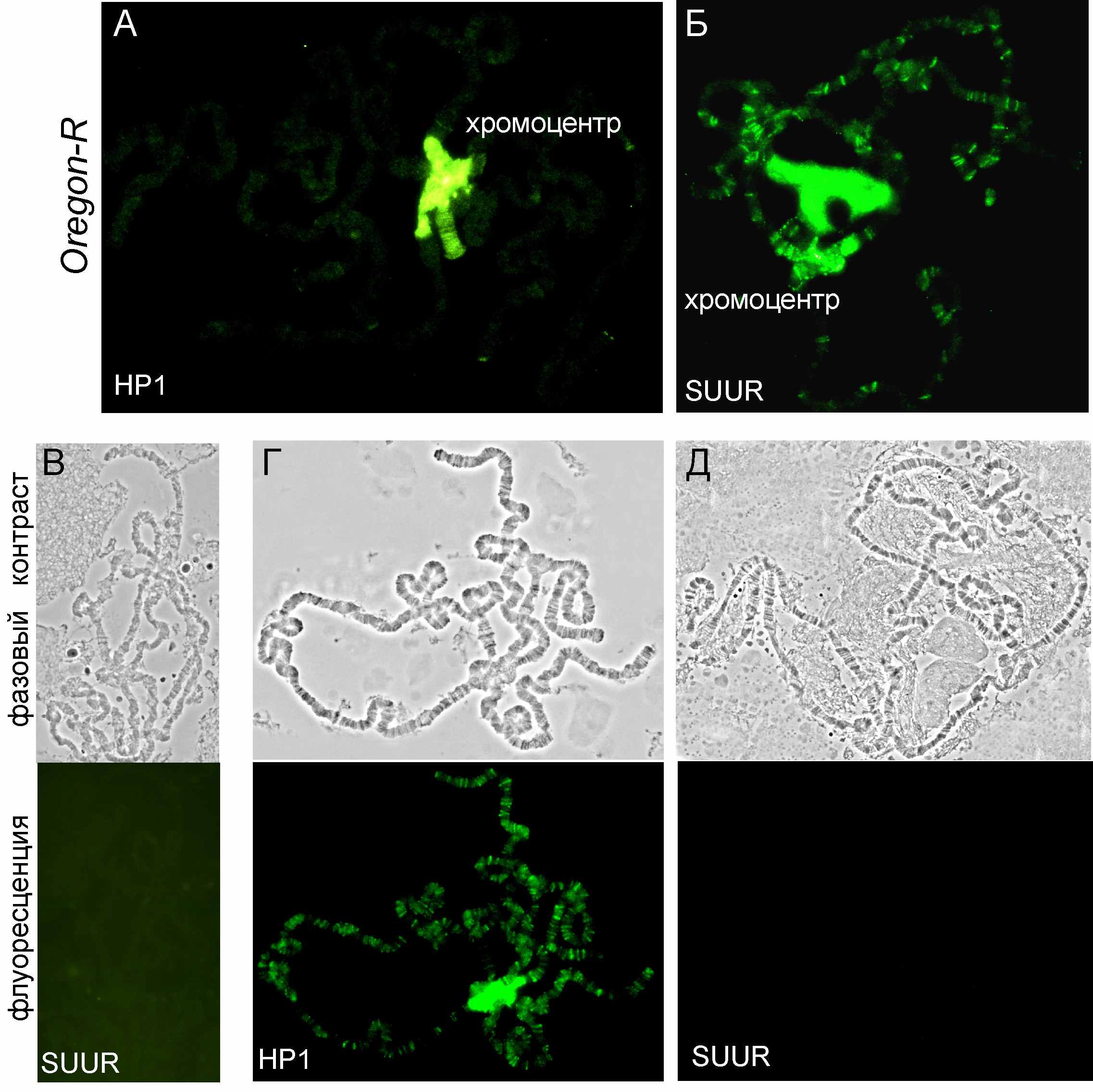
Рис. 3. Локализация на политенных хромосомах линии дикого типа Oregon-R белков HP1(А) и SUUR(Б). Отсутствие белка SUUR на хромосомах у 0-мутантов НР1(B). Локализация белка НР1(Г) и отсутствие белка SUUR(Д) при оверэкспрессии НР1.
Чтобы определить, сказывается ли делокализация белка SUUR при оверэкспресии НР1 на его функции, мы проверили, происходит ли подавление недорепликации ДНК в интеркалярном гетерохроматине, что характерно для мутантов SuUR. Для этого выбрали фрагмент Abd-A3 из максимально недореплицированной части района 89Е (Рисунок 4,А) и обнаружили, что при овреэкспреcсии НР1 политенизация этого фрагмента повышается до 20%, что говорит о нарушении функции белка SUUR (Рисунок 4,Б).
Таким образом, нормальное количество белка НР1 необходимо для нормального функционирования и локализации белка SUUR.

Рис. 4. А. Профиль недорепликации района 89Е1-2 в дикой линии OregonR и у мутанта SuUR (Moshkin et al., 2001). Б. Саузерн гибридизация зонда AbdA3 с геномной ДНК линий y,w (контроль) и hs-HP1 после воздействия серийного теплового шока. Д – диплоидная ткань (нейробласты). СЖ – слюнные железы. Интенсивность сигналов исследуемых зондов соотнесена с таковой полностью политенизированного зонда rosy.
Отсутствие белка SUUR не влияет на распределение HP1, у мутантов SuUR картина локализации HP1 на хромосомах не отличается от нормальной. Однако, при оверэкспрессии SuUR, индуцированной тепловым шоком, когда резко возрастает число сигналов SUUR, множество сигналов локализации HP1 появляется в эухроматиновых плечах. В результате этого практически все районы связаны как с SUUR, так и с HP1. В возникающих эктопических сайтах SUUR и HP1 колокализуются (Рисунок 5). Аналогичная картина наблюдается при оверэкспрессии SUUR в системе GAL4>UAS после индукции Sgs3 драйвером.
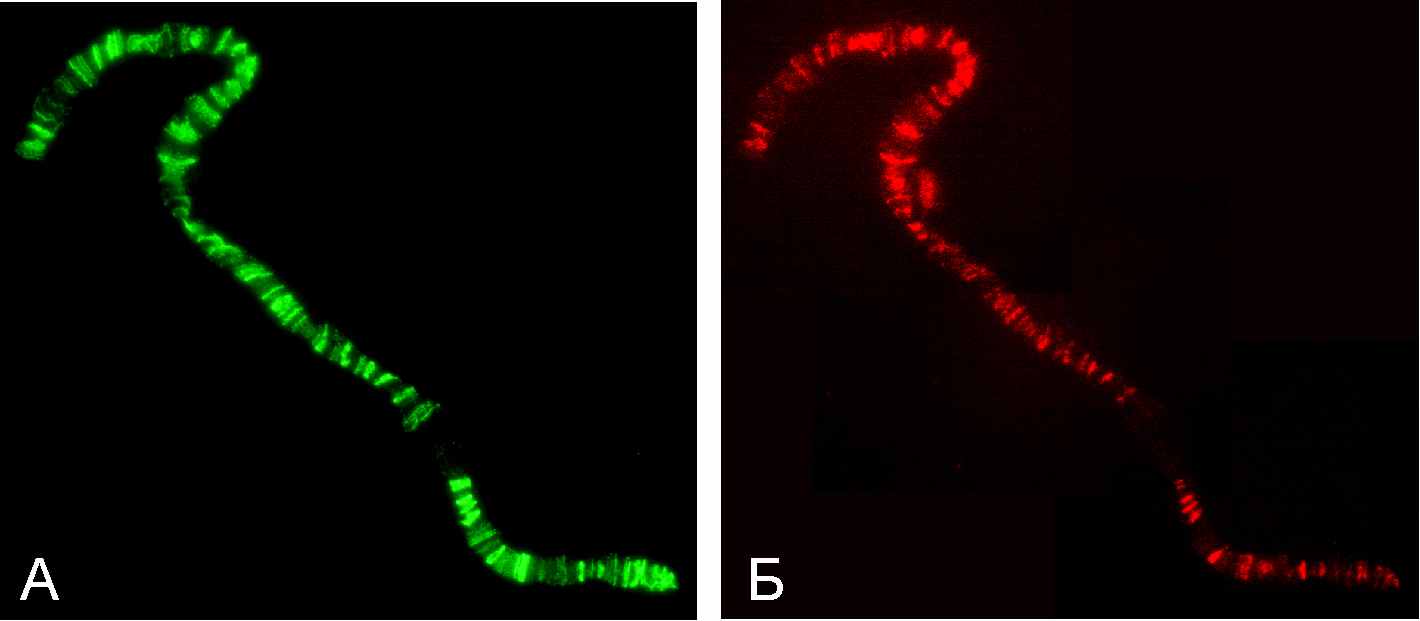
Рис.5. Локализация белков SUUR (А) и НР1 (Б) при оверэкспрессии SuUR, индуцированной тепловым шоком.
Возникновение дополнительных сайтов локализации НР1 на хромосомах при оверэкспресии SuUR может быть связано с усилением экспрессии гена Su(var)205, кодирующего НР1. Чтобы проверить это предположение, мы провели Вестерн гибридизацию антител против НР1 с белковыми экстрактами слюнных желез и показали, что при оверэкспрессии SuUR количество НР1 не возрастает, то есть, наблюдается именно перераспределение белка НР1, а не дополнительная экспрессия.
Для того, чтобы выяснить, какая часть белка SUUR является определяющей для перераспределения HP1, мы изучили распределение НР1 при экспрессии разных фрагментов белка SUUR на фоне мутации SuUR. Два из них, SUUR1-599 и SUUR1-799, - N-концевые фрагменты длиной с 1-й по 599-ю и с 1-й по 799-ю аминокислоты белка SUUR соответственно. SUUR495-962 - С-концевой фрагмент белка SUUR длиной с 495-й по 962-ю аминокислоты (Kolesnikova et al,, 2005). Полученные результаты показали, что оверэкспрессия N-концевых фрагментов SUUR1-599 и SUUR1-799 перераспределяет HP1 так же, как оверэкспрессия целого белка SUUR. Оверэкспрессия C-концевого фрагмента, SUUR495-962 не приводит к перераспределению НР1, и он остаётся в своих обычных сайтах - хромоцентре, 4 хромосоме, теломерах и 31 районе.
Таким образом, оверэкспрессия SuUR приводит к перераспределению НР1 на политенных хромосомах. Наличие неизмененной N-концевой части белка SUUR необходимо для наблюдаемого изменения паттерна локализации НР1.
Оверэкспрессия SUUR не приводит к изменению паттерна локализации белка HP2. На данный момент известно большое количество патрнёров НР1, имеющих различные функции и принимающих участие как в процессах генной репрессии, так и в репликации и репарации ДНК, а также в пространственной организации ядра. (Li et al., 2002; Hediger, Gasser, 2006). Можно ожидать, что обнаруженное нами перераспределение НР1 при оверэкспрессии SUUR отражается на паттерне локализации других белков – партнеров НР1. Хорошо изученным партнером НР1 является НР2, выявленный в дигибридной дрожжевой системе (Schaffer et al., 2002) и являющийся модификатором ЭПМ. Мы исследовали локализацию этого белка при перераспределении НР1 в результате оверэкспрессии SuUR. Оказалось, что оверэкспрессия SuUR, приводя к перераспределению НР1 на политенных хромосомах, не влияет на поведение НР2, и этот белок сохряняет нормальную локализацию (см. Таблицу 4).
Таблица 4. Локализация белков гетерохроматина в политенных хромосомах в зависимости от генотипа и экспериментальных условий.
| | SUUR | HP1 | HP2 | PC |
| Дикий тип OregonR | В хромоцентре, теломерах и в районах интрекалярно го гетерохроматина | В хромоцентре, теломерах и нескольких районах в плечах хромосом | В хромоцентре, теломерах и нескольких районах в плечах хромосом | Преимущест венно в районах интеркаляр ного гетерохрома тина |
| 0-мутация НР1 | Полностью отсутствует | Полностью отсутствует | Исчезает из теломер (Schaffer et al., 2006) | НД |
| Овер экспрессия НР1 | Полностью отсутствует | В нативных и 200 дополнитель ных сайтах в плечах хромосом | Следует за химерным белком НР1-PС (Shaffer et al., 2002) | НД |
| Мутация SuUR | Полностью отсутствует | Локализуется как в норме | Локализуется как в норме | Локализуется как в норме |
| Овер экспрессия SuUR | В хромоцентре и во всех дисках | В хромоцентре и во всех дисках | Локализуется как в норме | Локализуется как в норме |
| Одновременная овер экспрессия SuUR и НР1 | В хромоцентре и во всех дисках | В хромоцентре и во всех дисках | Локализуется как в норме | НД |
НД – нет данных о локализации белка
Белки SUUR и PС распределяются на хромосомах независимо. Помимо прицентромерного гетерохроматина, белок SUUR локализуется в районах интеркалярного гетерохроматина и обеспечивает недорепликацию этих районов в политенных хромосомах слюнных желез. Репрессия генов во многих районах интеркалярного гетерохроматина определяется комплексами, содержащими белки Pc-G. На политенных хромосомах слюнных желез эти белки в значительной степени колокализуются (Zhimulev et al., 2003). Мы проверили возможность взаимного влияния SUUR и PC. Ранее было показано, что мутация SuUR не влияет на локализацию сигналов связывания PC с хромосомами (Zhimulev et al., 2003). Мы обнаружили, что оверэкпрессия SuUR также не влияет на связывание белка PC с хромосомами и он локализован в своих обычных сайтах (см. Таблицу 4).
Мы исследовали также влияние оверэкспрессии Pс, индуцированной тепловым шоком в линии hs-Pc, на степень политенизации последовательностей в районе 19Е интеркалярного гетерохроматина. Методом количественной Саузерн гибридизации определили степень политенизации последовательности R(230-240) из района 19Е, в норме максимально недореплицированной (Рисунок 6, А). При оверэкспресии Pc степень политенизации этого фрагмента не изменялась, а у мутантов SuUR этот фрагмент оставался полностью политенизированным (Рисунок 6, Б). Таким образом, оверэкспрессия Pc не сказывается ни на недорепликации последовательностей интеркалярного гетерохроматина района 19Е, ни на влиянии мутации SuUR на этот процесс.
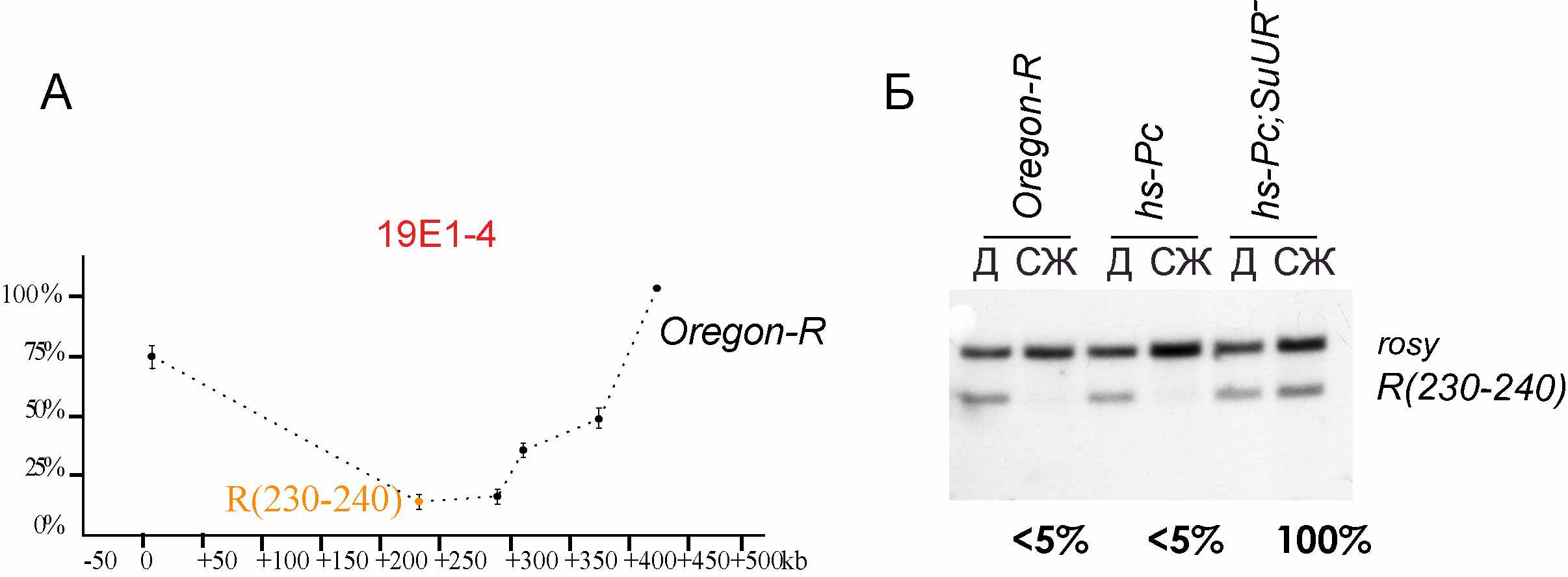
Рис. 6. А. Профиль недорепликации района 19Е1-4 в дикой линии OregonR. Б. Саузерн гибридизация зонда R(230-240) с геномной ДНК линий OregonR (контроль), hs-Pc и hs-Pc; SuUR- после воздействия серийного теплового шока. Д – диплоидная ткань (нейробласты). СЖ – слюнные железы. Интенсивность сигналов исследуемых зондов соотнесена стаковой полностью политенизированного зонда rosy.
Выводы
Ген SuUR является модификатором эффекта положения мозаичного типа: мутация SuUR супрессирует мозаичную инактивацию гена white+ в инверсии In(1)wm4h, гена yellow+ в дупликации Dp(1;f)1187 и гена scute+ в инверсии In(1)sc8. Увеличение доз гена SuUR и его оверэкспрессия усиливают репрессию этих генов при эффекте положения.
Белок SUUR появляется в эухроматиновых районах, гетерохроматизированных в результате эффекта положения, и является, таким образом, универсальным компонентом гетерохроматина в хромосомах Drosophila.
-
Мутация SuUR супрессирует недорепликацию в районах, гетерохроматизированных при эффекте положения.
Локализация белков НР1 и SUUR на политенных хромосомах взаимозависима:
а) в отсутствие НР1 SUUR не связывается с хромосомами
б) оверэкспрессия НР1 также препятствует связыванию SUUR с хромосомами; в результате этого в интеркалярном гетерохроматине супрессируется недорепликация ДНК.
в) оверэкпрессия SuUR приводит к перераспределению белка НР1 в ядре – он появляется в многочисленных эктопических сайтах и колокализуется с белком SUUR.
Участок, необходимый для перераспределения НР1 находится в пределах N-концевой части белка SUUR с 1 по 599 а.к.
Локализация белков НР2 и PC не зависит от степени экспресcии и локализации SUUR.
Публикации
- Zhimulev I.F., Belyaeva E.S., Makunin I.V., Pirrotta V., Volkova E.I., Alekseyenko A.A., Andreyeva E.N., Makarevich G.F., Boldyreva L.V., Nanayev R.A., Demakova O.V. Influence of the SuUR gene on intercalary heterochromatin in Drosophila melanogaster polytene chromosomes // Chromosoma. 2003. V.111(6). P. 377-398.
- Zhimulev I.F., Belyaeva E.S., Makunin I.V., Pirrotta V., Semeshin V.F., Alekseyenko A.A., Belyakin S.N., Volkova E.I., Koryakov D.E., Andreyeva E.N., Demakova O.V., Kotlikova I.V., Kolesnikova T.D., Boldyreva L.V., Nanayev R.A. Intercalary heterochromatin in Drosophila melanogaster polytene chromosomes and the problem of genetic silencing // Genetica. 2003. V.117(2-3). P. 259-270.
- Belyaeva E.S., Boldyreva L.V., Volkova E.I., Nanayev R.A., Alekseyenko A.A., Zhimulev I.F.. Effect of the Suppressor of Underreplication (SuUR) gene on position-effect variegation silencing in Drosophila melanogaster// Genetics. 2003.V.165(3). P. 1209-1220.
- Alekseyenko A.A., Makarevich G.F., Boldyreva L.V., Belyaeva E.S., Zhimulev I.F. (2001). Genetic control of DNA underreplication of intercalary heterochromatin regions in Drosophila melanogaster // Chrom. Res. 2001. V.9 (suppl. 1). P. 119.
- Нанаев Р. А., Болдырева Л. В., Алексеенко А. А., Жимулев И. Ф. Особенности политенизации района интеркалярного гетерохроматина 19Е1-4 политенных хромосом Drosophila melanogaster // Цитология. 2002. Т. 44(9) С. 895.
- Л.В. Болдырева, Т.Д. Колесникова, А.А. Юрлова, Г.В. Похолкова, Е.С. Беляева, И.Ф. Жимулев. Белки НР1 и SUUR взаимодействуют в районах интеркалярного гетерохроматина политенных хромосом Drosophila melanogaster // Цитология. 2005. Т. 47(9). С. 795.