
Селекция и генофонд
| Вид материала | Лекция |
- Селекция баклажана для аридной зоны нижнего поволжья 06. 01. 05 Селекция и семеноводство, 336.5kb.
- Методические рекомендации по преподаванию биологии в 11 классе. Планы уроков «Селекция, 216.83kb.
- Селекция на повышение требований к ростовым веществам, 52.08kb.
- План: Что такое селекция? Из истории развития селекции, 53.14kb.
- Князев Андрей Николаевич, кандидат биол наук, доцент; биотехнология растений Калашникова, 92.43kb.
- Рабочая программа генетика и селекция Код дисциплины по учебному плану опд ф 1 для, 292.62kb.
- Автореферат диссертации на соискание учёной степени, 828.49kb.
- Л. П. Трошин, 158.41kb.
- Генетика, селекция, гибридизация рыб. Тезисы докладов, 21.08kb.
- Задачи селекции : повышение урожайности сортов культурных растений, увеличение продуктивности, 105.58kb.
Анализ данных указывает на то, что на ранних стадиях развития плода тонина волоса меньше, чем у плодов позднего этапа развития.
В целом с увеличением возраста плодов толщина волоса повышается. Однако эта закономерность на стадии каракуля нарушается: у отдельных сортов каракуля наблюдается уменьшение тонины волоса. Это объясняется тем, что у каракуля появляются пуховые волокна и за счет этого уменьшается тонина. Это подтверждается изменчивостью признака.
Измерение длины волоса от голяка III сорта до каракуля - выпоротка (табл. 5) показало, что с увеличением возраста плодов увеличивается длина волоса. Если у голяка в 116-дневном возрасте длину волоса принять за 100%, то у 147-дневного плода она составляет 680%, а при рождении — 730%, т.е. волос в 7,3 раза длиннее.
Среднесуточный прирост волоса у плодов, начиная со 116 дн. и до 147-дневного возраста, составил 0,234 мм. На ранних стадиях развития плода (стадия голяка) среднесуточный рост волоса составил 0,09 мм, на стадии развития каракульчи от III сорта до I-го среднесуточный прирост был равен 0,17 мм. К моменту рождения рост волоса ускоряется и составил 0,35 мм в сутки.
Аналогичный результат отмечен в работах К. И. Мамина (1980), изучавшего рост волоса на каракульских плодах в утробный период. По его данным, к моменту рождения среднесуточный рост волоса составляет 0,35- 0,40 мм.
Плоды 130 — 132 дн., шкурки которых были признаны одним и тем же сортом (каракульча I), по длине волоса почти не отличались друг от друга. Но в то же время видовой ассортимент полученных в эти сроки шкурок был неодинаковым: 75% 130-дневных плодов соответствовали каракульче I и II сорта и 25% — каракулю. В 131-дневном возрасте плодов 25% шкурок были отнесены к сорту каракульча I, 25% — к каракуль-каракульче и 50% — к сорту каракульча II сорта. 80% шкурок 132-дневных плодов оцениваются как каракульча I сорта.
Таблица 5. Длина волоса у плодов в зависимости от срока
плодоношения
| Возраст плода, дн | Вид, сорт | Длина волоса, мм | Рост, % |
| 116 | Голяк III | 1,25 | 100 |
| 118 | Голяк II | 1,50 | 125 |
| 119 | Голяк I | 1,60 | 128 |
| 121 | Каракульча III | 2,70 | 216 |
| 128 | Каракульча II | 3,27 | 262 |
| 130 | Каракульча I | 4,60 | 368 |
| 131 | Каракульча I | 5,15 | 412 |
| 132 | Каракульча I | 4,60 | 368 |
| 133 | Каракуль-каракульча II | 5,31 | 425 |
| 136 | Каракуль-каракульча I | 5,50 | 440 |
| 141 | Каракуль плоский тонкий | 6,25 | 500 |
| 142 | - « - жакет московский | 7,00 | 560 |
| 144 | - « - ребристый толстый I | 7,95 | 636 |
| 145 | - « - жакет I | 7,39 | 591 |
| 147 | - « - жакет толстый | 8,50 | 680 |
| 149 | - « - жакет I (от живого ягненка однодневного возраста) | 9,12 | 730 |
Такое явление, очевидно, объясняется тем, что в одних случаях волос плодов, достигая длины 4,6 — 5,15 мм формирует завитки, а в других случаях, при той же длине волоса, — муаровый рисунок, а завитки появляются не на всей площади шкурки (еще не сформированы).
Видимо, при одинаковом кормлении овец качество волоса у плодов зависит от индивидуальной скороспелости.
Гистологическая структура кожного покрова плода. Н.А.Диомидова (1954), проводившая детальное описание формообразовательных процессов кожи и шерсти у овец отмечает, что плодный период является временем интенсивного роста эпидермиса и дифференцировки всех структур кожи.
Установлены различия в строении кожи у грубошерстных и тонкорунных овец, проявляющиеся в направлении корней волос, характере их расположения, строении и глубине залегания луковиц, толщине кожи и отдельных ее слоев (Диомидова, 1932).
Исследованиями О.И.Бригиса (1932), И.Н.Дьячкова, Ф.Н.Киямова (1959), М.Н.Кешеварца (1962), Н.П.Ралдугиной (1964, 1989), Э.М. Эрмана, М.А.Виноградовой (1966), А.К.Лебедева и др. (1973), Б.К.Чариева (1976), С.А.Асамова, Р.Г.Валиева (1977) и других авторов установлено, что от степени развития кожного покрова во многом зависят товарные свойства каракульских шкурок, так как от особенностей строения кожи, толщина отдельных слоев, ее формы волосяных луковиц, расположение их в коже и изогнутости волосянных фолликулов во многом зависят завитковые качества каракульских ягнят.
Исследованиями Т.М.Ахмеджанова (1962) установлено, что различия в гистологическом строении кожного покрова (толщина кожи и ее отдельных слоев, глубина залегания и густота корней волоса и так далее) можно установить не только в постэмбриональный, но и в эмбриональный период развития животных.
Результаты наших исследований по изучению гистологической структуры общей толщины кожи и ее отдельных слоев у плодов разных окрасок и происхождения представлены в таблице 6.
Таблица 6. Толщина кожи и ее слоев у плодов разных окрасок и происхождения
мкм
| Показатели | Окраска плодов | ||
| черная каракульская | черная метисная | белая метисная | |
| Учтено плодов | 7 | 7 | 7 |
| Общая толщина кожи | 185214,7 | 13739,7 | 150110,9 |
| В том числе толщина: | | | |
| эпидермиса | 224,4 | 171,7 | 182,1 |
| Пилярного слоя | 111013,5 | 100212,5 | 110817,3 |
| Ретикулярного слоя | 72016,1 | 35412,8 | 37513,4 |
Достоверно утолщенную кожу имеют каракульские плоды черной окраски, по сравнению с метисными плодами белой и черной окрасок (Р < 0,01).
Эпидермальный слой имеет важное значение в формировании шерстного волокна. На изученных гистопрепаратах четко просматривается нижняя граница субэпидермального слоя и верхняя рогового. Толщина эпидермального слоя у плодов колеблется в пределах 17,0 - 22,0 мкм, однако разница статистически не достоверна (Р > 0,1), что, видимо, связано с отсутствием существенного влияния происхождения и окраски на эпидермальный слой плодов.
При микроскопическом изучении отдельных слоев кожи выяснилось, что гистоархитектоника соединительно-тканных волокон составляет четко выраженное ячеистое строение, особенно четко оно выражено в ретикулярном слое. При этом плотность пространственного взаиморасположения соединительно-тканных волокон выше в образцах кожи каракульских плодов черной окраски. Следует также отметить, что ретикулярный слой значительно толще у каракульских плодов черной окраски (720 мкм), чем у метисных плодов черной (354 мкм) и белой (375 мкм) окрасок, разница статистически высокодостоверна (Р<0,001). Различия, отмеченные в ретикулярном слое, не отразились на толщине пилярного слоя, где эти показатели колеблются в пределах 1002-1110 мкм.
Таким образом, на общее развитие кожи плодов существенное значение оказывает его происхождение. При этом разница наблюдается лишь в ретикулярном слое, где наблюдается более интенсивный рост у каракульских плодов черной окраски.
Как известно, волосяной покров является производным волосяных филликулов. От количества первичных и развившихся вторичных волосяных фолликулов зависит густота волосяного покрова и качество каракулевого меха.
Проведенные нами исследования морфологии волосяных фолликулов показывают (табл.7), что наибольшее количество волосяных фолликулов имеют метисные плоды белой и черной окрасок (134,4-138,5 шт/мм2). Этот показатель у каракульских плодов черной окраски составил 58,3 на квадратный миллиметр площади кожи, разница статистически высокодостоверна (Р<0,001). Эта разница образуется за счет в два раза большего количества вторичных фолликулов на единицу площади кожи у метисных плодов белой (42,7) и черной (49,4) окрасок в сравнении с каракульскими плодами черной окраски. Кроме того, у метисных плодов почти в 6 раз больше зачаточных фолликулов на единице площади кожи.
Эти данные позволяют сделать заключение, что чистопородные каракульские плоды отличаются большей скороспелостью развития волосяных фолликулов в сравнении с метисными плодами черной и белой окраски. И к моменту рождения у каракульских ягнят волосяной покров окажется более густоволосым. Но в дальнейшем, при развитии зачаточных фолликулов, фолликулярный фонд помесей обеспечит лучшую густоту шерстного покрова и больший настриг шерсти, нежели у чистопородных каракульских овец.
Таблица 7. Густота волосяных фолликулов в коже плодов разных
окрасок и происхождения, шт/ мм2
| Показатели | Окраска плодов | ||
| черная каракульская | черная метисная | белая метисная | |
| Количество плодов | 7 | 7 | 7 |
| Общее количество фолликулов | 58,3 | 138,5 | 134,4 |
| В том числе: | | | |
| первичных (ПФ) | 22,8 | 26,9 | 28,7 |
| вторичных (ВФ) | 23,9 | 49,4 | 42,7 |
| зачаточных (ЗФ) | 11,6 | 62,2 | 63,2 |
На основании вышеизложенного, можно заключить, что в коже плодов в зависимости от происхождения наблюдается существенные различия. В коже метисных плодов больше вторичных и зачаточных волосяных фолликулов, что сказывается на суммарном количестве фолликулов.
М.А.Виноградовой (1966, 1973) установлено, что на гистологическое строение кожи оказывают большое влияние генетические особенности овец различных типов и паратипические факторы.
Проведенные нами исследования общей толщины кожи и ее отдельных слоев у плодов, полученных при различных вариантах спаривания родительских пар по смушковому типу показали, что у плодов различного срока плодоношения сохраняется общая тенденция в отношении толщины эпидермиса, пилярного и ретикулярного слоев, о чем можно свидетельствуют материалы таблиц 8 и 9.
Полученные нами данные соответствуют общеизвестным положениям о том, что абсолютные и относительные показатели толщины слоев кожи у одного и того же животного неодинаковые и зависят от биологической значимости слоя и его функции.
Развитие репродуктивного пилярного слоя кожи плодов обусловлено не только сроками плодоношения, но также и смушковыми типами родителей.
Использование в подборе баранов жакетного смушкового типа приводит к увеличению толщины пилярного слоя кожи у плодов, а баранов ребристого типа к уменьшению его толщины, не зависимо от смушкового типа маток и сроков плодоношения. Так, при спаривании маток и баранов жакетного смушкового типа толщина пилярного слоя кожи у плодов в возрасте 129-130 дней была больше на 10,9 % мкм, а в возрасте 134-135 дней больше на 17,9 %, чем у плодов такого же возраста, от спаривания баранов и маток ребристого смушкового типа. При подборе баранов жакетного типа к маткам ребристого показатели были больше соответственно на 7,8% и на 5%, а при подборе этих же баранов к маткам кавказского смушкового типа, толщина пилярного слоя у плодов оказалась больше на 8,4% (129-130 дн.) и на 17,3% (134-135 дн.), чем у одновозрастных плодов при подборе к маткам кавказского же смушкового типа баранов ребристого типа. Это наглядно видно из данных таблиц 7 и 8.
При использовании баранов ребристого типа эти показатели соответственно равны 909,214,6 и 1152,112,1 мкм; 906,421,4 и 1097,718,6; 934,836,8 и 1040,731,1 мкм. Отсюда можно заключить, что на толщину пилярного слоя кожи плодов оказывает большое влияние и смушковый тип матерей. От маток кавказского смушкового типа получались плоды с наибольшей толщиной пилярного слоя.
Аналогичные данные получены и по толщине ретикулярного слоя.
Одинаковое изменение толщины основных слоев кожи привело к тому, что общая толщина кожи тесно взаимосвязана со сроками плодоношения и смушковыми типами родителей. При этом использование баранов жакетного смушкового типа приводит к увеличению общей толщины кожи. использование маток кавказского смушкового типа также способствует увеличению общей толщины кожи.
При использовании баранов плоского смушкового типа на матках различных смушковых типов общая толщина кожи плодов была средней при сравнении с общей толщиной кожи, полученной от аналогичных маток при подборе баранов жакетного и ребристого смушковых типов.
Морфологические особенности волосяного покрова плодов. При оценке товарной ценности каракульских шкурок важное значение имеет как длина волоса, так и ее тонина.
Н.Ф.Никольский (1932), А.С.Канцепольский (1964), А.Г.Шамсутдинов (1970), М.Д.Закиров (1971), М.А.Виноградова (1973), Х.И.Укбаев, Р.Д. Шамекенова (1984), Н.П.Роддугина (1989), Х.И.Укбаев (1989) и др. провели широкие исследования по изучению морфологических особенностей волосяного покрова ягнят каракульских овец различных окрасок и смушковых типов.
По мнению М.Д.Закирова (1971), волос каракульских овец имеет неодинаковый диаметр на всем протяжении. А.Г.Шамсутдинов (1970) предлагает оставлять на племя тех каракульских баранчиков, у которых волос уравнен по тонине на всем протяжении всей длины.
Нами изучены морфологические особенности волосяного покрова плодов различных окрасок и происхождения.
В целом хорошо прослеживается влияние генотипа каракульских овец. интервал по тонине между фракциями волос приближен к чистопородным каракульским особям, но вместе с тем высоко содержание переходных волос во взятых пробах. Однако данный фактор положительно влияет на плотность и упругость волосяного покрова шкурки.
Для выяснения причин перевоплощения красивого муарового рисунка у плодов в малоценные завитки ягнят при рождении, у новорожденных помесных ягнят, в области крестца методом сбривания брали пробы волосяного покрова.
Исследования показали, что волосяной покров метисных ягнят на 44,3% состоит из ости, 24,4% - из переходного волоса и 31,3% - из пуха. По данным Б.Н.Васина (1941) у каракульских смушек первых и вторых сортов волосяной покров состоит из 63,3% ости и 36,7% пуха.
Толщина ости у основания волоса в среднем составляет 53,9 мкм, переходного волоса 38,7 мкм, пуха - 26,5 мкм. У каракульских ягнят, соответствующих по качеству смушка I - II сортам, толщина ости колеблется в пределах 32,2 – 39,7 мкм, а толщина пуха в среднем составляет 19,9 мкм при колебании от 16,9 до 20,5 мкм. Длина ости у чистопородных каракульских ягнят составляет 2,7-16,3 мм, а длина пуха 2,7-5,1 мм. Длина ости у помесных ягнят в среднем доходила до 10,9 мм, длина пуха -8,6мм.
Из сравниваемых данных видно, что между длиной ости у каракульских и помесных ягнят существенной разницы не наблюдается. Однако длина пуха у помесных ягнят в 2,2 раза больше, чем у каракульских. По-видимому, именно по этой причине шкурки метисных ягнят имеют более низкое качество. Длинный пух нарушает оптимальную структуру завитка, из-за чего он теряет упругость. Кроме того, нарушается необходимое соотношение волосков по толщине и длине.
На основе полученных материалов установлено, что кожа метисных плодов на 130 день утробного развития представляет собой вполне сформировавшийся орган с четко дифференцированными слоями и структурами и во многом схожа с гистологическим строением кожи чистопородного каракульского плода в том же возрасте.
Различия в качестве завитков помесных и чистопородных плодов отчетливо проявляется лишь к моменту рождения ягнят за счет резкого увеличения количества и длины пуха у метисных ягнят.
Основное отличие шкурок каракульских ягнят от шкурок ягнят других пород заключается в наличии у первых в преобладающем количестве своеобразных форм завитков – валька и боба.
Таблица 8
Толщина кожи и ее отдельных слоев у плодов в возрасте 129-130 дней
| Варианты подбора | Ед. изме-рения | Эпидермис | Пилярный | Ретикулярный | Общая толщина кожи |
| Жакетный х жакетный | мкм | 16,21,28 | 1017,914,8 | 317,211,6 | 1351,316,6 |
| | % | 1,2 | 75,3 | 23,5 | 100,0 |
| Жакетный х ребристый | мкм | 18,38,65 | 909,214,6 | 254,214,5 | 1181,717,0 |
| | % | 1,5 | 76,9 | 21,6 | 100,0 |
| Жакетный х плоский | мкм | 24,21,22 | 952,243,1 | 308,612,6 | 1285,033,1 |
| | % | 1,9 | 74,1 | 24,0 | 100,0 |
| Ребристый х жакетный | мкм | 17,21,07 | 980,628,9 | 294,117,6 | 1291,937,3 |
| | % | 1,2 | 75,0 | 22,9 | 100,0 |
| Ребристый х ребристый | мкм | 15,61,25 | 906,421,4 | 257,016,5 | 1179,014,6 |
| | % | 1,3 | 76,9 | 21,8 | 100,0 |
| Ребристый х плоский | мкм | 19,90,81 | 952,138,1 | 268,414,9 | 1240,441,4 |
| | % | 1,6 | 76,8 | 21,6 | 100,0 |
| Кавказский х жакетный | мкм | 17,81,40 | 1020,811,3 | 334,517,6 | 1373,122,8 |
| | % | 1,3 | 74,3 | 24,4 | 100,0 |
| Кавказский х ребристый | мкм | 17,21,33 | 934,836,8 | 308,621,7 | 1260,640,5 |
| | | | | | |
| Кавказский х плоский | мкм | 18,60,93 | 991,229,8 | 317,316,8 | 1327,138,8 |
| | % | 1,4 | 74,7 | 23,9 | 100,0 |
Таблица 9
Толщина кожи и ее отдельных слоев у плодов в возрасте 134-135 дней
| Варианты подбора | Ед. изме-рения | Эпидермис | Пилярный | Ретикулярный | Общая толщина кожи |
| Жакетный х жакетный | мкм | 18,61,11 | 1338,647,9 | 303,819,7 | 1661,142,5 |
| | % | 1,10 | 80,6 | 18,3 | 100,0 |
| Жакетный х ребристый | мкм | 16,79,58 | 1152,112,1 | 282,619,3 | 1451,424,3 |
| | % | 1,1 | 79,4 | 19,5 | 100,0 |
| Жакетный х плоский | мкм | 17,81,14 | 1221,122,6 | 297,125,7 | 1535,833,6 |
| | % | 1,2 | 79,5 | 19,3 | 100,0 |
| Ребристый х жакетный | мкм | 16,21,26 | 1210,419,1 | 279,411,5 | 1506,123,1 |
| | % | 1,1 | 80,4 | 18,5 | 100,0 |
| Ребристый х ребристый | мкм | 17,81,13 | 1097,718,6 | 277,011,4 | 1392,522,4 |
| | % | 1,3 | 78,8 | 19,9 | 100,0 |
| Ребристый х плоский | мкм | 23,21,16 | 1146,522,3 | 303,013,6 | 1472,624,7 |
| | % | 1,6 | 77,8 | 20,6 | 100,0 |
| Кавказский х жакетный | мкм | 19,40,87 | 1258,035,7 | 339,915,0 | 1617,343,4 |
| | % | 1,2 | 77,8 | 21,0 | 100,0 |
| Кавказский х ребристый | мкм | 15,61,28 | 1040,731,1 | 331,425,7 | 1387,749,6 |
| | % | 1,1 | 75,0 | 23,9 | 100,0 |
| Кавказский х плоский | мкм | 18,30,86 | 1226,527,7 | 388,723,0 | 1633,546,6 |
| | % | 1,1 | 75,1 | 23,8 | 100,0 |
В результате комплексных исследований гистоморфологических признаков кожноволосяного покрова разновозрастных плодов и новорожденных ягнят выявлены общие закономерности и особенности, присущие помесям и чистопородным каракульским овцам разных окрасок и смушковых типов; вскрыта причинная обусловленность процессов формообразования волосяного покрова в эмбриогенезе и шерстного покрова после рождения; установлена степень и характер модификации биолого-хозяйственно-ценных признаков кожноволосяного покрова плодов каракульских овец как результат взаимодействия организма суягных маток и факторов среды.
Современное состояние пастбищного овцеводства Казахстана ставит перед наукой и практикой все более сложные требования. Возникают такие проблемы, как быстрейшее восстановление численности поголовья овец; сохранение, размножение и рациональное использование оригинальных окрасок и расцветок каракульских овец. Эта проблема является центральной в селекции каракульских овец на повышение всех видов их продуктивности и улучшение качества. Поэтому, несмотря на достигнутые успехи науки и практики в разведении каракульских овец, научный поиск решения проблем продолжается, «в книге науки нет последней страницы.
ГЛАВА III
ГЕНЕТИЧЕСКАЯ СТРУКТУРА ПОПУЛЯЦИИ
КАРАКУЛЬСКИХ ОВЕЦ
Под популяцией в каракулеводстве понимают стадо каракульских овец
с определенной генной структурой, общностью обитания, приспособленное к
конкретным природно-климатическим условиям среды и размножающиеся
путем скрещивания особей между собой.
Термин популяция был предложен впервые в 1903 году Иоганссеном и
означает совокупность генетически различных особей одного вида,
обитающих совместно.
В 20-х годах прошлого столетия в результате синтеза
менделеевского и биометрического направлений генетики в качестве основы
современной селекции создавалась популяционная генетика. Популяционная
генетика изучает не отдельных, а группы особей, так называемые популяции.
Популяционной генетикой называют раздел генетики, изучающий
структуру и динамику генетического состава популяций, закономерности
наследования в популяциях.
Основной задачей популяционной генетики является исследование
генетического строения популяции в определенный период времени, путем
математико-статических методов и изменение строения этих популяций под
влиянием определенных факторов (отбора, мутации, миграции). Для
достижения определенной цели в селекции необходимо глубже изучить и
определить генетические параметры популяции.
Основы учения о генетической структуре популяций были заложены
выдающимся ученым-генетиком XX века С.С.Четвериковым. Его
исследования по генетической теории эволюции легли в основу науки,
получившей название - генетика популяции.
По мнению Ф.В.Ивлева (1984) с генетической точки зрения при
изучении популяции особый интерес представляет, во-первых, генетическая
структура данной популяции в определенный отрезок времени, во-вторых, ее
динамика, то есть изменение генетической структуры популяции в ряде
последующих поколений. При этом следует отметить, что под контролем
естественного отбора в популяциях происходит постоянное изменение
наследственности, т.е. одни генотипы, более приспособленные, спустя ряд
поколений приобретают преимущества, а менее приспособленные
относительно сокращаются. В результате, за счет смены частот генов
происходит длительная направленная перестройка генофонда.
Зная зависимость между генетическими параметрами популяции можно
описать все происходящие в ней селекционно-генетические процессы, а, зная характер и направление происходящих в структуре популяции можно судить
о количественных и качественных сдвигах в популяции за одно поколение и
о взаимосвязях этих процессов.
Генотипы каждого поколения возникают вследствие объединения гамет,
образованных предшествующим поколением. В свою очередь, гаметы
объединяются в процессы оплодотворения, образуя новый набор генотипов.
Тем самым, при скрещивании между членами популяции происходит
постоянный обмен генами. Следовательно, в каждом новом поколении
происходит реконструкция генофонда.
Изучение генетической структуры популяции поможет в практической
селекции выявить потенциальные генетические ресурсы селекционируемых
стад, прогнозировать их совершенствование.
Основным предметом популяционной генетики является анализ
популяции генов, то есть определение соотношения генов и генотипов в
одной популяции в определенный отрезок времени. Изучение любой
популяции начинается с изучения фенотипов особей.
Генетическая структура популяции более четко выявляется при анализе
качественных показателей, а значительно труднее провести генетический
анализ популяции по количественным признакам, которые контролируются
многими генами. Генетическая характеристика той или иной популяции по
количественным признакам дается не по соотношению отдельных генов и
генотипов, а по основным параметрам популяционной генетики
(наследуемость (h2), корреляция (r), средняя арифметическая (M), среднее
квадратическое отклонение (σ), коэффициент изменчивости (Сv) и другие).
Из всех этих показателей наибольшее значение с точки зрения генетической характеристики популяции имеет коэффициент наследуемости (h2).
Таким образом, изучение генетической структуры популяций помогает в
практической селекции выявить потенциальные генетические ресурсы
селекционируемых стад, прогнозировать их совершенствование.
Популяция должна характеризоваться генетическим гомеостазом,
способствующим ее сохранению и развитию, т.к. условия обитания стада
могут меняться, а популяция должна располагать генетическим резервом
изменчивости, позволяющим формировать новые свойства, которые в
процессе отбора будут закреплены или устранены.
Гомеостаз - это свойство популяции сохранять свою генетическую
структуру в ответ на условия внешней среды. По данным М.А.Ширинского
(1975) в госплемзаводе «Задарьинский» бараны-улучшатели с высоким
гомеостазом дали каракульских шкурок первых сортов больше на 7,2%, чем
производители с низким гомеостазом. Установлено, что отбор производителей с высоким гомеостазом по первым и жакетным сортам
каракульских шкурок снимает застой селекции и способствует повышению
качества каракуля.
В.С.Жилякова (1974) отметила, что отобранные бараны-производители -
улучшатели имели высокий гомеостаз независимо от пастбищно-кормовых
условий. Бараны-улучшатели 8692-8693 и 7673-7674 сур золотистый
полукруглого смушкового типа, класса - элита обладали высоким
гомеостазом наследуемости (0,75-0,86%), большой степенью посветления
волоса (0,82-0,86 и 0,75-0,81) в благоприятные и неблагоприятные по
кормовым условиям годы.
Если в популяции происходит свободное скрещивание особей любого
генотипа, с которыми не проводят планомерного отбора и подбора, не
наблюдается мутаций, популяция называется панмиксической или свободно
размножающейся. Это означает, что подбор пар происходит случайно, без
влияния и ограничения со стороны какого-то фактора. В результате
происходит случайное сочетание генотипов родителей у потомков. При этом
в популяции обнаруживается определенное постоянство в соотношении
генотипов и частоте генов равновесия, которое сохраняется до тех пор, пока
какой-либо фактор не выведет эту популяцию из генетического равновесия.
Если в популяции осуществляется отбор, происходит мутация, то ее
генетическая структура изменяется. При этом, прямые генные мутации
приводят к изменению доминантного аллеля (А) в рецессивный (а). Мутация
может быть и обратной, т.е. рецессивный аллель изменяется в доминантный.
Чем больше численность популяции, тем сильнее давление мутации на
структуру популяции.
Для определения доли гомозиготных доминантных (АА) и доли
гетерозиготных доминантных форм (Аа) в какой-либо популяции, не
отличающихся друг от друга по фенотипу, легко можно установить
соотношение всех трех генотипов в популяции по формуле Гарди-Вайнберга:
Р2 АА + 2*р*qAa + q2 aa, где
р и q – частота встречаемости аллелей А и а.
Сумма частот (р+q) равна 1.
По рецессивному фенотипу и формуле Гарди Вайнберга можно
получить информацию о соотношении генотипов в панмиктических
популяциях, выяснив сдвиги, происходящие в частоте генов под влиянием
различных факторов,
В каракулеводстве конкретным материалом применения этого метода
служат результаты гомогенного подбора родительских пар серых
каракульских овец. В стаде серых каракульских овец при гомогенном подборе серая овцематка х баран серой окраски из 100 родившихся ягнят
пало с признаками тимпанита 25 голов, что является результатом действия
рецессивного гена а, находящегося в гомозиготном состоянии. Поэтому,
для селекционера большое значение имеет определение, сколько среди
родившихся ягнят имеется особей с генотипом АА и Аа и установить частоту
доминантного (А) и рецессивного гена (а) в этой популяции.
Так, среди родившихся ягнят погибшие составляют 25%, которые имеют
гомозиготный генотип аа, что соответствует по формуле Гарди - Вайнберга
q2 aa = 0,25. Извлекая квадратный корень из этой величины, получаем частоту
рецессивного аллеля а:
q
 a = √0,25 = 0,447
a = √0,25 = 0,447Как было отмечено выше, сумма частот доминантного и рецессивного
аллеля равна единице (рА + qа = 1), то величина доминантного аллеля может
быть получена из выражения:
рА = 1 – qa
В нашем примере qа = 0,447, то
рА= 1- 0,447 = 0,553
По формуле Гарди-Вайнберга доля гомозиготных доминантных
генотипов АА будет равна:
р


 АА = √р2АА = √0,5532 = 0,3088
АА = √р2АА = √0,5532 = 0,3088Доля гетерозиготных доминантных генотипов Аа согласно той же
формуле составляет:
2p q Aa = 2 553 0,443 = 0,4944
Таким образом, популяция ягнят серой окраски при гомогенном подборе
родительских пар будет иметь следующее соотношение генотипов:
АА – 0,3058 или 30,58% или
Аа – 0,4944 или 49,44% или
аа – 0,20 или 20,00% или
∑ - 100% или 100 голов
Генная частота в популяции изменяется под влиянием мутации,
непрерывно совершающихся на уровне аллелей, хромосом.
Мутация - это внезапная, новая наследственная качественная
изменчивость. Под влиянием мутаций в популяции изменяется генная
частота непрерывно совершенствующих на уровне аллелей, хромосом.
Термин «мутация» был внесен голландским ботаником Гуго де Фриз,
одним из трех ученых вторично открывших законы Менделя. Он производил
опыты на многих растениях, в частности на ночной красавице, которая растет
в Голландии в диком состоянии. Пересаживая эти растения в свой сад и скрещивая их друг с другом обнаружил появление много необычных форм,
резко отличающихся от первоначального дикого типа. Самое главное,
впоследствии эти формы сохраняли свое своеобразие. Для этих форм,
возникающих в результате внезапных изменений, придумал название
мутации (К.Вилли, 1968).
Если в популяции осуществляется отбор в результате происходит
мутация, то ее генетическая структура изменяется.
К изменению доминантного аллеля (А) в рецессивный (а) приводят
прямые генные мутации. Реже встречается обратная мутация, когда
рецессивный аллель изменяется в доминантный. Установлено, что чем
больше численность популяции, тем сильнее давление мутаций на структуру
популяции.
Роль мутации в каракулеводстве огромна, т.к. использование ее
позволяет создать новые оригинальные расцветки каракульских шкурок.
Мутации приводят к созданию гетерозиготных организмов, которые
могут быть лучше или наоборот хуже приспособлены к условиям
существования, т.е. в зависимости от условий среды эффект мутаций бывает
разным (Е.К.Меркурьева, 1977).
Из популяции генные мутации исчезают сразу, если гетерозиготные
особи с новой аллельной мутацией не оставляют потомства. При этом,
рецессивные мутации, находясь у гетерозиготных особей в скрытом
состоянии, создают потенциальную генетическую изменчивость популяции.
В то же время следует отметить, что в большой популяции давление
мутационного процесса приводит к закреплению аллеля или к
сбалансированному равновесию.
Сохранение и дальнейшее развитие популяции происходит благодаря
получению нового поколения. Следует иметь в виду, что не все особи
участвуют в получении следующего поколения. Одни исключаются заранее в
результате естественного отхода, другие выбраковываются. Поэтому,
эффективная численность популяции определяется числом участвующих в
размножении особей.
Генетические параметры популяции
Коэффициент наследуемости. Селекция по тем признакам, фенотип
которых теснее связан с племенной ценностью, считается более успешной.
Эта связь выражается коэффициентом наследуемости (h2). Термин
наследуемость предложен американским генетиком Дж.Лашем (1939). Символ наследуемости h2 заимствован у С.Райта (1921), который обозначал
им детерминацию признаков и наследуемость. Наследуемость - это
отношение наследственно обусловленной генетической вариансы к общей наблюдаемой фенотипической вариансы. Большое значение в генетических
исследованиях фенотипической изменчивости количественных признаков
играет выявление коррелятивных связей между дочерями и матерями, между
полусестрами по отцу или матери, между группами родственных мужских
особей и т.д.
В целях выявления доли влияния генетического фактора на
количественный признак потомков С.Райттом было начато изучение связей
между родственными группами животных. В основе которой был положен
математический метод, позволивший разработать систему генетико-
математического анализа наследования и закономерностей изменчивости
количественных признаков, прослеживаемых в популяциях животных.
Выдающаяся заслуга С.Райтта заключается в том, что им была разработана
модель, схематически выражающаяся через цепь звеньев связи фенотипов и
генотипов между родственными особями, которая изображена следующим
образом (Е.К.Меркурьева, 1970):
σVpVn = 0,5


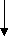 генотип родителей генотип потомка h= rVpФp h= rVnФn
генотип родителей генотип потомка h= rVpФp h= rVnФnфенотип родителей rФpФn фенотип потомка
Величина h – это коэффициент пути от генотипа к фенотипу;
Связь между генотипом и фенотипом выражена величиной h= rVФ
Связь между фенотипами родителей и потомков выражена rФpФn.
В каракулеводстве в основном используется термин коэффициент наследуемости (h2), который колеблется от 0 до 1. Чем выше величина
коэффициента наследуемости, тем больше фенотипическая изменчивость
признака. При высоком генетическом разнообразии в популяции величина h2
выше. Необходимо обратить внимание на то, что тесное и длительное
родственное спаривание приведет к снижению генетического разнообразия
популяции, т.е. характер подбора и структура стада оказывают существенное
влияние на величину коэффициента наследуемости. При этом следует иметь
в виду, что уменьшение генетического разнообразия приводит к малой
эффективности массовой селекции. В этом случае племенная работа должна основываться на индивидуальном отборе и подборе, анализе сочетаемости
пар, применении линейного разведения и т.д.
Уровень кормления, условия содержания, сезон года, возраст животного
также оказывают влияние на величину коэффициента наследуемости.
По М.Е.Лобашеву (1967) знание наследуемости необходимо для
потенциальной эффективности отбора, выбора максимально целесообразных
методов оценки признаков и научно обоснованного планирования роста
продуктивности животных. Коэффициент наследуемости h2 какого-либо
признака позволяет судить о том, в какой степени его можно увеличить
путем селекции.
Установлено, что при разработке вопросов селекции каракульских овец
необходимы знания о наследуемости основных хозяйственно-полезных
признаков. По мнению А.Н.Плохинского (1964), П.Ф.Рокицкого (1964),
Г.А.Стакана, А.А.Соскина (1965) наследуемость хозяйственно-полезных
признаков сельскохозяйственных животных варьирует в очень широких
пределах. Основным фактором, определяющим наследуемость хозяйственно-
полезных признаков является наследственное разнообразие родителей и
генетическая структура стада. Чем шире генетическое разнообразие стада,
тем естественно оно будет богаче представлено у потомков.
По данным Ш.А.Ахметова и Р.Е.Асылбекова (1985) коэффициент
наследуемости большинства хозяйственно-полезных признаков у
каракульских овец несколько выше и более достоверен как по серым, так и
по черным ягнятам в опытной группе, чем в контрольной (табл.10).
Коэффициент наследуемости в опытной группе черных ягнят был
достоверным по живой массе, толщине кожи и по основным промерам
экстерьера, а у серых также по ширине завитков, что свидетельствует об
эффективности отбора по этим признакам.
По данным Т.У.Умурзакова (1992) коэффициент наследуемости,
вычисленные различными методами, составили по типу рисунка 0,206-0,235,
по длине завитка 0,067-0,266, по удельному весу вальковатых завитков
(фигурность) 0,044-0,191. Им установлена высокая наследуемость и
селекционное значение экономически важного признака запаса кожи у ягнят.
Коэффициент наследуемости его составил у ягнят задарьинского заводского
типа 0,34, а у ягнят темирского заводского типа каракульских овец - 0,76.
Полученные данные подтверждают мнение Я.Л.Глембоцкого и
Г.Я.Копыловской (1972) о том, что значительная часть признаков
разводимых пород овец в том числе и смушковых характеризуется высокой
степенью наследуемости, т.е. на уровне достаточном для эффективного
массового отбора генотипическим разнообразием популяции отдельных стад.
При проведении племенной работы в стаде, линии или породе в целом,
имея данные о величине коэффициента наследуемости, можно решить ряд
вопросов.
Зоотехнику-селекционеру следует строго разграничивать понятия
«наследование», «наследственность» и «наследуемость».
Н.А.Плохинский (1969) считает, что наследование - процесс передачи
наследственной информации в поколениях, т.е. от родителей к детям и ее
реализации в форме признаков по определенным законам, а наследуемость -
это наследственная обусловленность разнообразия каждого признака,
проявление генетической информации, относящейся к данному признаку в
его фенотипическом разнообразии.
Таблица 10. Коэффициент наследуемости некоторых селекционно-
генетических признаков у каракульских овец
| Признаки | Степень наследуемости | Авторы | |||||
| серые | черные | ||||||
| опытная | контроль нал | опытная | контроль ная | | |||
| живая масса | 0,19- | 0,15 | 0,31- | 0,07 | Ш.А.Ахметов, РЕАсылбеков, 1985 | ||
| толщина кожи | 0,02 | 0,02 | 0,33- | 0,03 | |||
| длина черного волоса | 0,10 | 0,06 | 0,03 | 0,07 | |||
| длина белого волоса | 0,14 | 0,05 | | | |||
| ширина завитков | 0,76- | 0,10 | 0,12 | 0,06 | | ||
| длина завитков | 0,14 | 0,24 | 0,14 | 0,03 | |||
| высота в холке | 0,33- | 0,11 | 0,23- | 0,05 | |||
| косая длина туловища | 0,30- | 0,20 | 0,32- | 0,14 | |||
| обхват груди | 0,31- | 0,16 | 0,28- | 0,08 | |||
| обхват пясти | 0,20- | 0,22 | 0,13 | 0,06 | |||
| Примечание - Р<0,05 | |||||||
| длинный полукруглый валек | 0,28 | | | | В.С.Жилякова, 1978 | ||
| тонкая кожа | 0,37 | | | | |||
| короткий валек | 0,45 | | | | |||
| толстая кожа | 0,58 | | | | |||
| тип рисунка | 0,206- 0,235 | | | | Г.У.Умурзаков 1992 | ||
| длина завитка | 0,067- 0,266 | | | | |||
| фигурность | 0,044- 0,191 | | | | |||
По утверждению Ф.Е.Ильева (1984) очень важно понять, что
наследуемость - явление популяционное, оно изучается не по отдельным
особям, а на уровне популяции. Это такой показатель, который позволяет
решить задачу, поставленную М.Ф.Ивановым еще в 1949 году - «Ищите
лучшие генотипы среди лучших фенотипов». Отсюда следует, что тот, кто
уделяет все внимание лишь одному животному, теряет из виду всю
популяцию.
Наследственность - это передача потомкам генетической информации,
точнее генетических факторов (генов), которые обуславливают формирование у них родительских признаков и свойств или сходства родителей и их
потомков.
М.Д.Закиров (1989) считает, что если материнская форма имела генотип
Аа и, следовательно, образовала галоидные яйцеклетки А и а, то при
оплодотворении их мужской гаметой, у всех зигот будет проявляться
только доминантный аллель А, несмотря на то, что половина зигот будет
обладать генотипом аа. Это явление объясняется тем, что доминантный
аллель А у исходной материнской формы Аа еще до метода детермировал
свойства цитоплазмы яйца по типу А, т.е. в данном случае фенотип особи не
соответствует ее генотипу.
По данным В.С.Жиляковой (1981) отбор ягнят золотистой и серебристой
расцветок от гомогенного подбора увеличивает наследуемость серебристой
расцветки от 0,39 и 0,41 (у рядовых и улучшателей) до 0,83 (у линейных
производителей); наследуемость золотистой расцветки увеличивалась
соответственно от 0,41 и 0,74 до 0,88.
С увеличением ценности производителей возрастает наследуемость
большой степени посветления от 0,40 и 0,48 до 0,63, интенсивной расцветки
от 0,47 у рядовых до 0,85 у улучшателей и линейных производителей и
контрастного перехода темного основания к светлому концу от 0,39 и 0,69 до
0,86. Последнее определяет увеличение наследуемости желательного типа
животных от 0,33 и 0,69 у рядовых и улучшателей до 0,89 у линейных
производителей.
Наследуемость полукруглого смушкового типа колеблется от 0,42 у
рядовых и 0,44 у улучшателей до 0,46 у линейных производителей;
соответственно ребристого смушкового типа от 0,17 у рядовых до 0,15 у
улучшателеи и линейных производителей и плоского смушкового типа - от
0,13 и 0,11 до 0,8.
Наследуемость плоского и ребристого смушковых типов с увеличением
ценности производителей уменьшается. Наследуемость кавказского
смушкового типа не зависит от ценности производителей и составляет - 0,28.
Полная извитость полукруглого валька является желательной,
наследуемость ее колеблется от 0,28 до 0,34 (у рядовых и улучшателей) и до
0,43 (у линейных производителей).
С увеличением ценности производителей наследуемость малой и
неполной извитости уменьшается от 0,89 до 0,20.
Рядовые производители и улучшатели имеют низкую 0,09 и 0,14
наследуемость очень шелковистого и шелковистого волоса. Для усиления
наследуемости этих признаков к линейным относили производителей,
которые имели очень шелковистый волос, что позволило увеличить
наследуемость очень шелковистого волоса до 0,40 и шелковистого до 0,93 (у
линейных производителей). Отрицательным моментом племенной работы
является высокая наследуемость - 0,74 и 0,84 у рядовых и улучшателеи до
0,89 (у линейных производителей) недостаточной шелковистости путем
снижения наследуемости ее. Наследуемость сильного блеска не зависит от
качества производителей и составляет 0,44.
Достоверность Р изученных признаков колеблется от 0,95 до 0,99.
Для увеличения количества животных желательного типа необходимо
проводить селекцию на увеличение наследуемости желательных признаков
путем использования на всем стаде маток линейных производителей. После
проверки производителей по качеству потомства к улучшателям и линейным
производителям следует относить баранов, которые имеют высокую
наследуемость всех желательных смушковых признаков.
Кроме того, определение коэффициентов наследуемости находит
применение для оценки племенной препотентности производителей. Например,
при спаривании лучших производителей сур с уравненными по степени
посветления волоса матками оказалось, что при дифференцированном
анализе потомства от разных баранов коэффициенты наследуемости этого
показателя имели заметно различные величины.
Сопоставляя наследуемость конкретного показателя с индивидуальными
племенными качествами баранов, удалось обнаружить, что чем больше коэффициент наследуемости признака, тем производитель более устойчиво
передает потомству свои особенности.
Массовая селекция на среднюю и большую степень посветления волоса,
которая характеризовалась высоким коэффициентом наследуемости и имела
большой коэффициент корреляции с выраженностью расцветки,
способствовала созданию стада золотистых и серебристых суров с
константной наследственностью указанных свойств.
Коэффициент корреляции (r). Ф.В.Ильев (1984) считает, чтобы повысить
эффективность отбора прежде всего необходимо установить, в какой мере
тот или иной признак по которому ведется селекция зависит от
наследственных особенностей животного (от генотипа) и в какой - от среды
(от фенотипа). Поэтому, выявление степени зависимости между фенотипом и
генотипом является одной из важнейших задач селекции.
Наиболее распространенным параметром, выявляющим связь между
показателями и признаками каракульских овец, считается коэффициент
корреляции (r). Он выражает связь между признаками в относительных
величинах, а именно в виде дроби, которая находится в пределах от 0 до ±1.
Если коэффициент корреляции близок к нулю, то это указывает на
отсутствие связи. Принято считать связь малой, если коэффициент
корреляции имеет уровень 0,2-0,3; средняя величина близка к 0,5, если
коэффициент больше 0,7, то связь считается высокой.
Если коэффициент корреляции имеет знак плюс (+), то это означает, что
связь между признаками прямая или положительная, при которой с
изменением одного признака, другой изменяется в том же направлении. Знак
минус (-) свидетельствует о наличии обратной или отрицательной связи
между признаками.
Доказано, что путем племенного подбора можно резко изменить даже прочно сложившиеся связи и создавать взамен существующих
корреляций между признаками животных новые более удовлетворительные
соотношения. Следовательно, человек может управлять этими связями путем
соответствующего отбора и подбора родительских пар.
При положительной корреляции двух желательных признаков изменить
соотношение желательных качеств гораздо сложнее, так как при усилении
одного признака будет изменяться другой.
И.Я. Аверьяновым (1951) установлена положительная корреляция
конституции с размером завитка.
У потомства каракульских овец нежной конституции преобладает мелкий
размер завитка, у овец грубой конституции - крупный, крепкой конституции -
средний.
Использование в селекционной работе показателей размера завитка каракульских ягнят при рождении и конституции при подборе во взрослом
состоянии повышает качество каракуля.
Нами изучены корреляции между длиной волоса и другими
селекционируемыми признаками каракуля (табл.11).
Приведенные данные показывают, что некоторые признаки: смушковый
тип (r=0,498), размер завитка (r=0,789), живая масса (r=0,324),толщина кожи
(r=0,635) и срок эмбрионального развития ягнят (r=0,598) положительно и
статистически достоверно коррелируют с длиной волоса (r ≈ от 0,197 до
0,789). При этом по мере увеличения длины волоса каракуля жакетной
группы приобретают переходный характер в сторону перерослого
кавказского типа.
Длина волоса имела отрицательную, статистически достоверную
корреляцию с классностью (r= -0,563), длиной валька его плотностью (r=
-0,713), шелковистостью (r= -0,573 и блеском (r= -0,601), то есть удлинение
волоса способствовало снижению степени выраженности вышеуказанных
признаков.