
Экология зоопланктона разнотипных водоемов бассейна Верхней Волги
| Вид материала | Автореферат диссертации |
- Зоопланктон разнотипных водных экосистем бассейна верхней оби, 420.52kb.
- Республика Марий Эл расположена в центре европейской части Российской Федерации,, 938.98kb.
- Партийно-государственная политика по формированию инженерно-технической интеллигенции, 390.4kb.
- Экология водоемов зоны техногенной радиационной аномалии на южном урале 03. 00., 1110.67kb.
- Славяне Введение, 467.33kb.
- Эколого-гидрологические исследования водоемов, 425.38kb.
- Культура Руси в период, 138.66kb.
- Промышленная экология методы и способы защиты воздушного бассейна, 721.61kb.
- Тульский Государственный Университет кафедра"Истории и культурологии" Контрольно-курсовая, 141.72kb.
- Эколого-фаунистический анализ паразитофауны рыб вислинского залива, рек преголи и прохладной, 419.14kb.
Общая характеристика водоемов. Бассейн Верхней Волги занимает территорию между 55о и 61о с.ш. в средней части Восточно-Европейской равнины в подзонах средней и южной тайги. Нижней границей считают место впадения в Волгу р. Шексна или плотину Рыбинской ГЭС (Волга и ее жизнь, 1978), в последнее время к Верхней Волге относят и бассейн Горьковского водохранилища (Экологические проблемы.., 2001). Бассейн включает ряд крупных водохранилищ (Верхневолжское, Иваньковское, Угличское, Рыбинское и Шекснинское) и значительное количество небольших озер, основная часть которых сосредоточена в западной части Вологодской области. Озерность варьирует от 2% на юге до 6-10% на севере (Волга и ее жизнь, 1978; Экологические проблемы.., 2001).
Таблица 1. Общая характеристика исследованных озер.
| Группа водоемов | Площадь, км2 | Hmax, м | Проточность | рН | РОВ, мгС/л |
| Озера Молого-Шекснинской низины (Дарвинский заповедник, Вологодская обл., Череповецкий р-н), 23 озера | 0.005-6.4 | 0.8-3.5 | Замкнутые 52% Сточные 40% | 4.4-10.1 | 0.9-64.2 |
| Озера верховьев р. Суда (Вологодская обл., Бабаевский р-н), 10 озер | 0.1-10.0 | 1.0-8.0 | Сточные 50% Проточные 30% | 4.1-7.5 | 8.2-34.4 |
| Озера верховьев р. Шуя (Карелия, Суоярвский р-н), 6 озер | 1.0-13.7 | 1.8-6.0 | Проточные 83% | 4.6-6.7 | 9.6-62.7 |
| Вендюрско-Вохтозерские озера (Карелия, Кондопожский р-н), 11 озер | 0.3-10.0 | 3.0-13.0 | Замкнутые 55% Проточные 36% | 4.1-8.9 | 1.9-8.4 |
Примечание: Hmax – максимальная глубина, РОВ – содержание растворенного органического вещества.
Основная часть работ выполнена на Рыбинском – наиболее крупном среди водохранилищ Верхней Волги, площадь его водосбора 150500 км2, зеркала 4550 км2, средняя глубина 5.6 м, коэффициент условного водообмена 1.9 год–1, средний годовой приток воды 35.4 км3, его воды относятся к маломинерализованным гидрокарбонатным группы кальция, среднегодовая сумма ионов в Главном плесе составляет 180 мг/л (Экологические проблемы.., 2001). Водоем в современный период считают умеренно эвтрофным (Минеева, 2004). Величина рН воды летом 7.0-8.9 с максимумом в центре водохранилища, цветность 30-140 град. Pt-Co шкалы, наиболее гумифицированы воды Моложского и Шекснинского плесов (Экологические проблемы.., 2001).
Малые озера исследовали в рамках программы по оценке влияния антропогенного и природного закисления вод на их экосистемы. В качестве реперных выбраны небольшие (<10 км2) мелководные маломинерализованные водоемы, расположенные вблизи водоразделов крупных речных бассейнов, где хозяйственная деятельность и локальное антропогенное воздействие минимальны. Исследованы две группы озер (33 водоема) в бассейне Рыбинского водохранилища (Вологодская обл.) и, для сравнения, две группы озер (17 водоемов) в бассейне Онежского озера в Карелии (табл. 1). Сравнительно крупные исследованные озера ледникового происхождения (Антипов, 1975), большинство малых представляют вторичные водоемы, образовавшиеся в процессе неравномерного роста торфяника (Иванов, 1957; Лесненко, Абросов, 1973). Их трофический статус варьирует от олиго- и ультраолиготрофного до эвтрофного и зависит от особенностей водного питания (Лазарева и др., 1998; Лазарева, Комов, 1998). По степени закисления вод озера подразделяли на: нейтральные (рН >6.5), переходные (рН 5.4-6.4) и ацидные (рН <5.3) (Комов, Лазарева, 1997). По уровню гумификации выделяли светловодные (олигогумозные) водоемы – (РОВ <10 мгС/л, цветность 20-50 град.), переходные (мезогумозные) – (РОВ 10-14 мгС/л, цветность 50-100 град.) и темноводные (полигумозные) – (РОВ >15 мгС/л, цветность 100-250 град.). Среди последних выделяли подгруппу сильно гумифицированных водоемов (РОВ >40 мгС/л, цветность >250 град.), наиболее богатых аллохтонным органическим веществом (Лазарева, 1991; Комов, Лазарева, 1994; Лазарева и др., 1998). С учетом гидрологического режима выделены 4 типа болотных водоемов (табл. 2).
Материалы. В основу работы положены материалы, полученные при проведении комплексных экспедиционных работ на Рыбинском водохранилище в 1956-2005 гг. и малых озерах его бассейна в 1983-2006 гг. В них вошли:
- сезонные наблюдения (архивные материалы), выполненные с мая по октябрь на 6 станциях Рыбинского водохранилища (1956-1995 гг. каждые 2 нед.)
- круглогодичные наблюдения, выполненные на 5 станциях Рыбинского водохранилища (1983-1988 гг. ежемесячно зимой и два раза в месяц летом), на 8 болотных озерах Дарвинского заповедника (1983-1996 гг. ежемесячно зимой и с интервалом в 1-2 нед. летом).
- крупномасштабные пространственные съемки, выполненные в период максимального прогрева воды в Рыбинском водохранилище (июнь-август 1997-2001 гг. 14-16 станций, июль-август 2003-2005 гг. 15-25 станций), то же в озерах (май-август 1983-1985 32 озера, июль 1988 г. 20 озер, август 1990 27 озер, август 1991 г. 17 озер)
- съемки мелководных участков Рыбинского водохранилища (май-октябрь 1983-1988 гг. 5 станций, август 2002 г. 22 станции) и 4-5 озер Дарвинского заповедника (май и июль 1983, май-октябрь 1987, июль и октябрь 1993).
- отдельные съемки на озерах бассейна водохранилища зимой (март) и в течение вегетационного периода (1988-2006 гг.).
Таблица 2. Типы болотных озер, обусловленные гидрологическим режимом водосборов (по: Лазарева и др., 1998).
| Стоково-нейтральные вблизи вершины болотного массива (F<5) | Стоково-приточные на краю болотного массива (F 7-25) | ||
| Светловодные (олигогумозные) | Темноводные (полигумозные) | ||
| РОВ <10 мгС/л | РОВ 15-39 мгС/л | РОВ>40 мгС/л | |
| В составе анионов доминирует SO42–, Ca+Mg2+ <200 мкг-экв./л, железо не входит в число главных ионов (Feобщ <0.5 мг/л) | В составе анионов доминирует HCO3–, Ca+Mg2+ до 1400 мкг-экв./л, железо входит в число главных ионов (Feобщ >0.4 мг/л) | ||
| Ацидные, рН<5.3 | Нейтральные, рН>6.5 | ||
| Эпитермические | Эпи- или метатермические | Эпитермические | Метатермические |
| Олиготрофные Хл а 1.1-2.1 мкг/л | Мезотрофные Хл а 5.0-8.1 мкг/л | Эвтрофные Хл а 28.2-77.3 мкг/л | |
| Средняя (V-X) численность зоопланктона, тыс. экз./м3 | |||
| 42-253 | 24-630 | 127-1353 | 787-2483 |
| Тип 1. В питании преобладают атмосферные осадки и поверхностные воды верхового болота | Тип 2. В питании преобладают поверхностные воды верхового болота | Тип 3. В питании преобладают воды переходного болота | Тип 4. В питании преобладают грунтовые воды торфяника |
| Мотыкино Дорожив | Дубровское Змеиное Утешково | Хотавец Мороцкое | Кривое Высоцкое |
Примечание: F – коэффициент удельного водосбора, Хл а – (по: Минеева, 1994).
Методы сбора и обработки данных. Для сбора зоопланктона на Рыбинском водохранилище до 1962 г. использовали планкточерпатель Богорова 50 л, в 1962-2001 гг. – планктобатометр Дъяченко-Кожевникова 10 л, с 2002 г. большинство проб собрано малой сетью Джеди с ситом №70 (диагональ ячеи 120 мкм). Основная часть проб на озерах (1-2 станции в центре) собрана малой сетью Джеди с диаметром входного отверстия 12 см и ситом №42 (диагональ ячеи 213 мкм). В них учитывали ракообразных и коловраток Asplanchna. Прочие виды коловраток и науплиусов копепод подсчитывали в 0.5 л нефильтрованной воды после отстаивания. Пробы фиксировали 4% формалином, камеральную обработку проводили по принятой в гидробиологии методике (Киселев, 1969). Биомассу зоопланктеров определяли по таблицам зависимости массы организмов от длины тела (Балушкина, Винберг, 1979). К доминантным относили виды рачков с плотностью 5% от суммарной для ракообразных и виды коловраток с плотностью 5% от суммарной для данной таксономической группы (Лазарева и др., 2001; Lazareva, 1995).
Для анализа структуры зоопланктона рассчитывали 7 групп показателей. Как характеристики видового богатства анализировали общее количество видов в списке (Sобщ), число видов в пробе (Sпр) или найденных в водоеме за год, индекс Маргалефа (S – 1)/ln Nобщ (Песенко, 1982). Уровень доминирования в сообществе измеряли с использованием индексов Симпсона (ISimp = ∑pi2) и Бергера-Паркера IB/P = pmax) (Песенко, 1982; Мэгарран, 1992), по количеству доминантов (Андроникова. 1996), а также графическим способом (Уиттекер, 1980; Одум, 1975). Видовое разнообразие зоопланктона определяли по индексу Шеннона –∑pi log2pi, который рассчитывали по биомассе (Hb) или численности (Hn) (Гиляров, 1972; Песенко, 1982). В ряде случаев использовали показатели, выводимые из индекса Симпсона 1/∑pi2, 1 – ∑pi2 (Песенко, 1982).
Для оценки изменения состава видов и трофического статуса водоемов по зоопланктону использовали фаунистический индекс трофности (Е) (Мяэметс, 1980):
Е = К(х + 1)/(А+У)*(у + 1)
где: К, А, У – число видов коловраток, копепод и кладоцер соответственно, х – число видов-индикаторов мезо- и эвтрофии, у – число видов-индикаторов олиго- и мезотрофии. Соотношение обилия ракообразных и коловраток (Bcr/Brot, Ncr/Nrot) рассчитывали для оценки изменчивости размерной структуры сообщества и кормового значения зоопланктона для рыб-планктофагов. Динамику размерно-массовой структуры сообщества исследовали по показателю средней индивидуальной массы особей (wср = Вобщ/Nобщ) (Крючкова, 1987; Андроникова, 1996). В большинстве случаев, если это не оговорено отдельно, в работе анализировали средние за сезон (май-октябрь) величины общей численности (Nобщ) и биомассы (Вобщ) зоопланктона.
Всего собрано и обработано автором >300 проб зоопланктона из пелагиали и столько же из литорали водохранилищ, >600 проб из озер, систематизированы и проанализированы материалы почти 3000 архивных проб. На всех станциях проводили определение фоновых характеристик. Кроме того, автором по материалам комплексных экспедиций проанализированы водный баланс и годовой бюджет основных ионов сложного водосбора оз. Хотавец (Лазарева, Комов, 1998), особенности круговоротов органического вещества, азота, фосфора и серы в болотных водоемах (Лазарева и др., 1999, 2000 а,б).
По данным 1983-1995 гг. для озер различных трофических типов с модальным уровнем рН воды от 4.5 до 7.2 сделана оценка динамичности фауны зоопланктона по методу (Титлянова, 1993) в модификации автора. На основе летних (июль-август) сборов 1984-1993 гг. выполнена классификация сообществ зоопланктона разнотипных озер и ординация по градиентам закисления и гумификации по методике (Миркин, 1974). Схема анализа временных рядов принята (по: Пановский, Брайер, 1972). Анализ трофической структуры зоопланктона проводили (по: Pimm et al., 1991). При обработке данных использованы стандартные статистические программы для персонального компьютера.
Глава 3. Факторы, определяющие динамику и сукцессию зоопланктона водоемов бассейна Верхней Волги
К основным факторам, заметно влияющим на динамику и сукцессии водных сообществ в регионе можно отнести три: динамику климата, сукцессии экосистем водоемов и закисление поверхностных вод. Закисление поверхностных вод определяет сукцессию экосистем и сообществ малых болотных водоемов бассейна Верхней Волги. Для водохранилищ антропогенное закисление не представляет большого значения в силу нечувствительности большей части их водосборов к поступлению кислот из атмосферы (Комов и др., 1997). Однако трофический статус водохранилищ в значительной степени зависит от притока гумифицированных вод, динамика которого тесно связана с динамикой региональных климатических процессов и заболачивания водосборов (Экология фитопланктона…, 1999; Минеева, 2004; Лазарева, 2005).
Из известных причин закисления для озер региона наиболее важны две: антропогенная, связанная с поступлением окислов серы и азота с атмосферными осадками, и природная, которую определяет вынос органических кислот из болот (Комов, Лазарева, 1994). Закисление атмосферных осадков вызвано антропогенными выбросами кислотообразующих соединений (SOx, NOx). Закисление поверхностных вод возможно при поступлении сульфатов из атмосферы >1.5 г/м2 в год (The Stockholm Conference..., 1982). В начале 90-х годов ХХ в. в бассейне Верхней Волги кислотная нагрузка составила 2-3 г/м2 сульфатов (Комов, Лазарева, 1994; Комов и др., 1997). В 1988-1990 гг. среднегодовой уровень рН атмосферных осадков на побережье Рыбинского водохранилища составил 4.5-4.8 (Комов, Лазарева, 1994).
Максимально чувствительны к кислотному загрязнению воды с концентрацией Ca+Mg <400 мкгЭкв/л и общей минерализацией <50 мг/л (Manual for Chemical..., 1987). Из особенностей гидрологического режима важны доля атмосферных осадков в питании водосборов, горизонт почв, через который перемещается основной поток воды, и скорость водоотдачи водосбора (Miegroet, 1994; Landers et al., 1994). В западной половине бассейна Верхней Волги сильно закислены до 10% малых приводораздельных преимущественно болотных водоемов (Комов и др., 1997). Среди исследованных антропогенно закислены озера типов 1 и 2, которые питаются атмосферными осадками и поверхностными водами верховых олиготрофных болот (Комов, Лазарева, 1994; Лазарева, Комов, 1998). Мало чувствительны к современному уровню поступления кислотообразующих соединений из атмосферы болотные озера, которые питаются грунтовыми водами торфяников (тип 4), а также водоемы, которые питаются смешанными водами, поступающими из ландшафтов разного типа (тип 3). Эти озера в основном испытывают влияние природного закисления и даже при 4-5-ти кратном увеличении сульфатной атмосферной нагрузки не будут закислены ниже рН 5.5 (Комов, Лазарева, 1994).
Д
 ругая сторона проблемы закисления поверхностных вод – увеличение подвижности тяжелых металлов в водной фазе в ацидных озерах, что приводит к усилению их биоаккумуляции. Особенно опасна ртуть, основной источник поступления в атмосферу которой тот же (сжигание топлива), что и кислотообразующих соединений (Шипунов и др., 1981). Снижение рН повышает скорость метилирования ртути (Сафонова, 1989). Высокий уровень накопления ртути в биоте часто наблюдается именно в озерах с низкими значениями рН воды и характерен для водоемов Молого-Шекснинского водораздела (Комов, 1999). При рН <5 наибольший уровень накопления ртути наблюдается в организмах бентоса и рыбах, тогда как в зоопланктоне аккумуляция ртути мала и сопоставима с таковой в донных отложениях (Степанова, Комов, 1996).
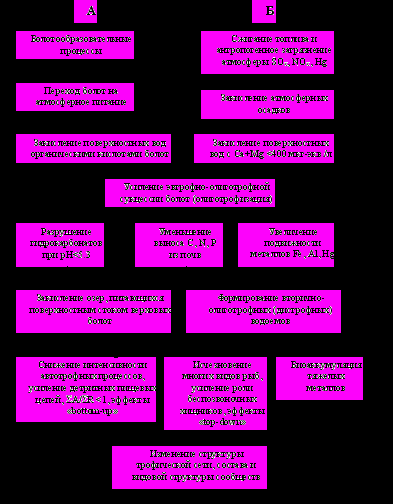
ругая сторона проблемы закисления поверхностных вод – увеличение подвижности тяжелых металлов в водной фазе в ацидных озерах, что приводит к усилению их биоаккумуляции. Особенно опасна ртуть, основной источник поступления в атмосферу которой тот же (сжигание топлива), что и кислотообразующих соединений (Шипунов и др., 1981). Снижение рН повышает скорость метилирования ртути (Сафонова, 1989). Высокий уровень накопления ртути в биоте часто наблюдается именно в озерах с низкими значениями рН воды и характерен для водоемов Молого-Шекснинского водораздела (Комов, 1999). При рН <5 наибольший уровень накопления ртути наблюдается в организмах бентоса и рыбах, тогда как в зоопланктоне аккумуляция ртути мала и сопоставима с таковой в донных отложениях (Степанова, Комов, 1996).Рис. 1. Схема развития процесса закисления в экосистемах болотных озер.
А - природная ацидификация, Б - антропогенная.
Загрязнение атмосферных осадков окислами серы в настоящее время характерно для большей части Европейской России (Dedkova et al., 1993) и представляет собой крупно-масштабный региональный процесс (рис. 1, ряд Б). Вызванное им закисление поверхностных вод наблюдается не во всех, а только в чувствительных к кислотным осадкам ландшафтах, для которых антропогенная сульфатная нагрузка превышает критическую (Комов, Лазарева, 1994; Комов и др., 1997; Лазарева и др., 1998; Моисеенко, 2003), что характеризует ацидификацию поверхностных вод как мозаичный региональный процесс. По мощности воздействия на водные экосистемы его можно сравнивать с эвтрофированием. Некоторые наблюдения свидетельствуют о противоположном направлении сукцессий экосистем озер, индуцированных эвтрофированием и ацидификацией (Grahn et al., 1974; Wetzel, 1983; Лазарева и др, 1998, 2000а).
Эвтрофно-олиготрофная сукцессия торфяных болот (рис. 1, ряд А), связана со сменой гидрологического режима при переходе торфяника по мере его роста на атмосферное питание (Иванов, 1957; Глазовская, 1981). Усиление зависимости ионного состава болотных вод от состава атмосферных осадков сказывается на гидрохимическом режиме болотных озер (Лазарева и др., 1998; Лазарева, Комов, 1998). Природное закисление вод озер мы рассматриваем как начальную стадию, а антропогенное воздействие кислотных атмосферных осадков – как последующую, заключительную стадию ацидификации. Такой подход совпадает со схемой взаимодействия природной и антропогенной ацидификации, принятой в Европе и Америке (Emmett et al., 1994).
Важнейшим следствием объединенного (природное + антропогенное) влияния закисления на экосистемы озер служат разрушение гидрокарбонатов и смена химического класса вод при рН <5.3 (Комов, Лазарева, 1994; Лазарева и др., 1998, 1999, 2000б). Вследствие этого еще более снижается и без того небольшое поступление в озера из болот азота, фосфора, а также углерода, минерального (Глазовская, 1981; Лазарева и др., 1999, 2000б; Grahn et al., 1974) и растворенного органического (Krug, Frink, 1983). Подобная картина характерна также для озер, водосборы которых расположены на минеральных почвах (Schindler, 1994), что позволяет говорить о глубоком изменении круговоротов азота, фосфора и углерода в водных экосистемах под влиянием атмосферного антропогенного закисления (Schindler, 1994; Sherman, Fairchield, 1994; Лазарева и др., 1999, 2000б).
Снижение притока биогенных элементов с водосбора при неизменной скорости водообмена приводит к уменьшению их запаса в донных отложениях озер вследствие выноса с водой за пределы экосистемы. Этот процесс называют «деэвтрофированием» (Мартынова. 1984; Лазарева и др., 2001; Лазарева, 2005а) или «олиготрофизацией» (Rhode, 1969; Grahn et al., 1974). Его темп может быть очень высоким из-за формирования «каскадных» эффектов: снижение притока C,N,P с водосбора вызывает обеднение илов C,N,P, снижение фотосинтеза и бактериальной продукции, уменьшение количества ежегодно образующегося ОВ и еще большее обеднение илов C,N,P (см. также Wetzel, 1983). По-видимому, именно такие эффекты определяют значимые корреляции характеристик кислотного загрязнения c показателями трофического статуса (Комов, Лазарева, 1994; Komov et al., 1997; Siegfried et al., 1989). Результатом подобных трансформаций в болотных водосборах служат вторично-олиготрофные озера, подобные рассмотренным в настоящей работе (типы 1 и 2).
Механизмы влияния ацидификации на сообщества озер не тождественны таковым при олиготрофизации, во-первых: из-за наличия прямого токсического действия тяжелых металлов и протонов. Во-вторых, при ацидификации эффекты «bottom-up» и «top-down» в трофической сети формируются фактически одновременно, тогда как при олиготрофизации воздействия «сверху», вызванные исчезновением отдельных видов рыб, возникают лишь при дефиците пищи для них. В большинстве американских озер не зарегистрировано снижение продукции фитопланктона при закислении, этот факт лежит в основе возражений против гипотезы олиготрофизации (Schindler, 1994). Напротив, в Скандинавии и Европейской России озера с рН <5.5 отличаются низкой биомассой водорослей, содержанием хлорофилла а и скростью фотосинтеза (Минеева, 1994; Корнева, 1994; Бульон, 1997; Никулина, 1997; Grahn et al., 1974).
Изменение регионального климата определяет гидрологический, химический и термический режим вод и наиболее отчетливо проявляется в откликах сообществ водохранилищ – инертных экосистем большого размера, влияние локальных возмущений в которых не так заметно по сравнению с региональными, влияющими на водосбор в целом (Романенко, 1985; Ривьер, 1988; Поддубная, 1988; Лазарева, 1997,1999, 2005; Пырина, 2000; Девяткин и др., 2000; Литвинов и др., 2005). Квазициклические колебания биотических характеристик экосистемы водохранилища тесно связаны с динамикой гелиофизических параметров и глобальных климатических характеристик (Смирнов и др., 1993; Девяткин и др., 1996; Литвинов и др., 2005). Они определяют климат в регионе, влияют на гидрологический и термический режимы водохранилища, а также на уровень продуктивности его экосистемы.
За время существования водохранилища выделяют несколько фаз водности (Литвинов и др., 2005): 1941-1950 гг., 1963-1976 гг. и с 1996 г. маловодные, 1951-1962 гг. и 1977-1995 гг. многоводные. Колебания стока рек включают квази-циклы с периодами 2-4, 8-12 и 19-33 года (Литвинов, Рощупко, 2000; Литвинов и др., 2005). Многолетний ход температуры воздуха имеет четкий 7-11-летний ритм, зависящий от изменения солнечной активности (Литвинов и др., 2002). Колебания гидрологических характеристик воздействуют на зоопланктон внутрикаскадного водохранилища двумя путями (рис. 2). Во-первых, непосредственно через изменение состава, структуры и объема биостока из рек и, особенно, из лежащих выше по каскаду водохранилищ. Во-вторых, косвенно через вариации скорости водообмена, уровенного и гидротермического режимов, условий нереста рыб, стартовых условий развития короткоцикловых видов водорослей, бактерий и беспозвоночных, количества и качества поступающего со стоком рек ОВ и степени его аккумуляции в донных отложениях (Лазарева, 1999).
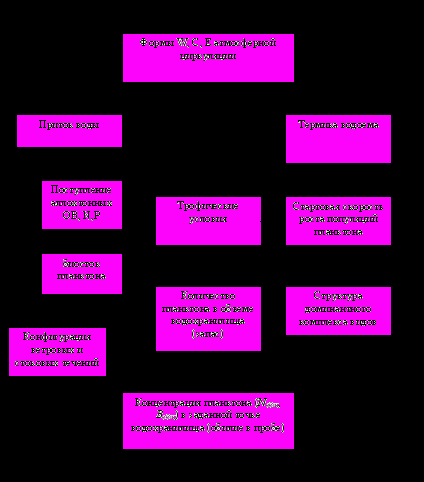
Рис. 2. Схема влияния динамики климатических характеристик на запас и концентрацию планктона во внутрикаскадном водохра-нилище.
Большинство авторов указывают на невозможность исследования сукцессий без учета квази-периодических колебаний праметров сообществ и экосистемы водохранилища (Романенко, 1985; Лазарева, 1997; Пырина, 2000; Лазарева и др., 2001; Литвинов и др., 2005). Наибольший размах вариаций характеристик наблюдается в сообществах, образованных короткоцикловыми видами. Минимумы и максимумы обилия бактерий, фито- и зоопланктона различаются в 4-6 раз (Рыбинское водохранилище, 1972; Романенко, 1985; Экология фитопланктона.., 1999; Лазарева и др., 2001).
Третьим важным для динамики сообществ фактором служит сукцессия экосистем водоемов, интенсивность которой определяется колебаниями климата – с одной стороны и антропогенным (локальным и фоновым) воздействием на водосборы и водоемы – с другой. В развитии экосистемы Рыбинского водохранилища выделяют 4 периода с различным уровнем продуктивности, границы между которыми размыты вследствие асинхронности процессов в разных компонентах экосистемы. 1 – первичное эвтрофирование от начала заполнения водохранилища (1941 г.) до 1955-1956 гг. или “мотылевая” стадия (по: Мордухай-Болтовской, Дзюбан, 1966; Баканов, Митропольский, 1982), в планктоне она прослеживалась до начала 60-х годов прошлого века (Лазарева, 2005). Первичное эвтрофирование было выражено слабо, вследствие перегруженности затопленных почв торфом (Сорокин, 1958). Формирование донных и планктонных сообществ тормозилось недостатком покоящихся стадий гидробионтов и небольшим поступлением легкоусвояемого ОВ со стоком рек и с берегов (Гусева, 1958; Поддубная, 1958; Мордухай-Болтовской, 1958; Воронина, 1959; Ривьер, 1998).
2 – период низкой продуктивности экосистемы (деэвтрофирование) с середины 50-х до конца 60-х годов ХХ в. (до середины второй маловодной фазы) (Баканов, Митропольский, 1982; Терещенко, Стрельников, 1997; Ривьер, 1998). В это время выпала основная часть затопленных лесов, приостановился размыв берегов, ложа водохранилища и торфяных сплавин (Зиминова, Курдин, 1972).
3 – вторичное эвтрофирование началось в маловодные 70-е годы и продолжалось до середины 90-х годов ХХ века. Эту стадию отмечают по изменениям фитопланктона, зоопланктона и зообентоса (Баканов, Митропольский, 1982; Ривьер, 1998; Экология фитопланктона.., 1999; Лазарева и др., 2001; Экологические проблемы.., 2001), дестабилизации в сообществе рыб (Терещенко, Стрельников, 1997), расширению зарослей воздушно-водных растений (Рыбинское водохранилище, 1972). Чаще всего вторичное эвтрофирование опеделяют как антропогенное (Баканов, Митропольский, 1982; Ривьер, 1998; Экология фитопланктона.., 1999). Однако, на мой взгляд (Лазарева, 2005), основной его причиной стало расширение площади высокопродуктивных серых илов с 8% площади водохранилища в 1955 г. до 40% в 1978 г. (Законнов, 1981), а также усиление аутогенных процессов в экосистеме в маловодные 70-е годы.
В многоводные 80-е годы ХХ в. площадь серых илов сократилась с 40 до 17% площади водохранилища (Законнов, 1995). Это сказалось на развитии бентоса (олигохеты, хирономиды), до 1990 г. его биомасса оставалась на уровне 70-х годов ХХ века (Перова, 1999; Экологические проблемы.., 2001). Это же, вероятно, способствовало снижению выноса в толщу воды легкоусвояемого ОВ, что вместе с усилением аллогенных процессов в экосистеме определило рост численности длинноцикловых видов зоопланктона (рачки) и появление в его структуре признаков, указывающих на деэвтрофирование экосистемы (Лазарева и др., 2001). Значительный вклад в изменение направления сукцессии экосистемы внесло уменьшение антропогенной нагрузки на водосбор (снижение площади пахотных земель, поголовья скота), начавшееся в 90-х годах ХХ в (Лазарева, 2007а). Во второй половине 90-х годов констатировано снижение трофического статуса водохранилища по содержанию хлорофилла а от эвтрофного до умеренно эвтрофного (Минеева, 2004).
В конце ХХ в. Отмечено усиление биоинвазионных процессов в водохранилищах Волги, которое связывают с особенностями глобальных климатических процессов в бассейнах Балтийского, Азовского и Каспийского морей (Яковлев, 2005). В 80-90-х годах в Рыбинском водохранилище колонии дрейссены Dreissena polymorpha (Pallas) сформировали новый высокопродуктивный биотоп заиленного ракушечника (Перова, Щербина, 1998). На рубеже веков отмечено распространение второго вида D. bugensis (Andrusov) (Экологические проблемы.., 2001). Численность велигеров (до150 тыс. экз./м3) во второй половине лета стала сравнима с количеством зоопланктона (Лазарева. 2005б). Велигеры дрейссены конкурируют за пищу с пелагическими простейшими и коловратками, для успешного развития которых необходима высокая концентрация пищи (Гутельмахер и др., 1988). Снижение в 1.5-2 раза численности коловраток, вызванное распространением дрейссен, стало заметно с 1992 г. (Лазарева и др., 2001; Лазарева, 2005а,в). В конце 90-х годов ХХ в. произошло вселение в водохранилище нового планктофага тюльки Clupeonella cultriventris (Nordmann) (Экологические проблемы.., 2001). С 2000 г. тюлька образует до 97% численности рыб пелагического комплекса (Герасимов, 2005). Успешной ее натурализации способствовала высокая численность крупных форм зоопланктона (роды Leptodora, Bythotrephes, Heterocope), предпочитаемого корма для этого вида рыб (Лазарева, 2005в; Кияшко и др., 2007). Таким образом, в 90-х годах прошлого века начался новый период сукцессии экосистемы, связанный с активизацией непланктонных инвазионных видов, способных формировать каскадные эффекты в трофической сети (Лазарева, 2005б).