
Академия педагогических и социальных наук московский психологосоциальный институт антропология хрестоматия
| Вид материала | Документы |
- Д. И. Фельдштейн Заместитель главного редактора, 6159.28kb.
- Л. М. Семенюк психологические особенности агрессивного поведения подростков и условия, 1581.61kb.
- Московский Центр непрерывного математического образования, 51.2kb.
- Педагогическое сопровождение подготовки по информационной безопасности в системе высшего, 143.17kb.
- Российская академия наук отделение общественных наук ран, 74.85kb.
- Московский Государственный Институт Электроники и Математики (Технический Университет), 10.69kb.
- Программа воспитания и обучения в детском саду, 3924.08kb.
- Программа воспитания и обучения в детском саду, 3936.51kb.
- Программа воспитания и обучения в детском саду, 3718.01kb.
- Программа воспитания и обучения в детском саду, 3919.5kb.
Схема расовой классификации по И. Е. Деникеру (с. 16)
Общее для всех классификаций XX в. проведение принципа иерархического соподчинения в группировке расовых типов. Нет ни одной схемы классификации, в которой выделенные типы, даже если их число невелико, выстраивались бы в ряд один подле другого. Любой исследователь непременно объединяет их в более крупные генетически связанные группы, исходя из имеющихся данных об их родстве и своих представлений в области расовой генеалогии. Таким образом, оба основных достоинства классификационной системы И.Е. Деникера были безоговорочно приняты исследователями позднейшего времени.
До сих пор активно дебатируется проблема числа основных рас. <...>
Существующие точки зрения варьируют от гипотезы двух основных расовых стволов, начиная с автора этой гипотезы Т. Гексли (Huxley, 1870), причем гипотеза сама выступает то в форме противопоставления европеоидов и монголоидов расам тропического пояса (Keith, 1949; Siasutti, 1959), то в форме противопоставления европеоидов и негроидов монголоидам (Рогинский, 1941; Алексеев, 1969), и до гипотезы 15 самостоятельных рас (Бунак, 1934).
В.В. Бунак в более поздней работе (1956) группирует расы, правда, в четыре ствола, но не отвергает свои прежние представления. Между этими крайними точками зрения лежит широкий спектр гипотез, постулирующих и три, и четыре, и пять основных расовых стволов» (с. 18).
Хрисанфова Е.Н., Перевозчиков И.В. «Антропология», 1991:
«В нашей стране наиболее известны три классификации. Одна приведена в учебнике «Антропология» Я.Я. Рогинского и М.Г. Левина, вторая предложена Г.Ф. Дебецем, третья разработана В.В. Бунаком.
Первая имеет довольно строгую структуру: три большие расы подразделяются на малые расы, промежутки между большими заполнены шестью промежуточными расами. В названиях соблюден географический принцип Символ схемы — круг».
В качестве примера приведем классификацию Я.Я. Рогинского и М.Г. Левина как одну из наиболее известных.
Рогинский Я.Я., Левин М.Г. «Антропология», 1978:
«При построении классификации учитывались: степень морфологического сходства рас, их географическое распространение и по возможности древность их формирования, причем последняя в ряде случаев за отсутствием материала устанавливалась и использовалась лишь предположительно.
| Большие расы | Расы |
| Экваториальная, или австралонегроидная | Австралийская |
| Веддоидная | |
| Меланезийская | |
| Негрская | |
| Негрилльская (центральноафриканская) | |
| Бушменская (южноафриканская) | |
| | Эфиопская (восточноафриканская) |
| Южноиндийская (дравидийская) |
| Евразийская, или европеоидная | Атлантобалтийская |
| Среднеевропейская | |
| Индосредиземноморская | |
| Беломорскобалтийская | |
| Балканокавказская | |
| | Южносибирская (туранская) |
| Уральская | |
| Азиатско-американская, или монголоидная | Североазиатская |
| Арктическая (эскимосская) | |
| Дальневосточная | |
| Южноазиатская | |
| Американская | |
| | Полинезийская |
| Курильская (айнская) |
На схеме видно, что промежуточное место между экваториальной и евразийской большими расами занимают две расы: эфиопская и южноиндийская; между евразийской и азиатско-американской размещаются также две расы — южносибирская и уральская; наконец, особое место по значительной части признаков промежуточное между американской и австралийской занимают полинезийская и курильская расы (рис. 183).
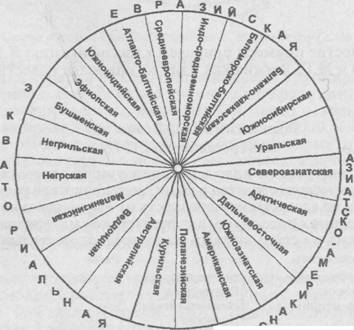
Рис. 183. Схема расовой классификации
Вопрос о происхождениях этих промежуточных форм разрешается различно для каждой из них» (с. 374).
Хрисанфова Е.Н., Перевозчиков И.В. «Антропология», 1991:
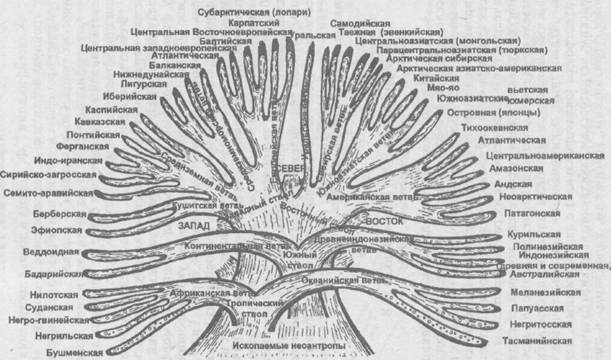
«Классификация Г.Ф. Дебеца в логике построения имеет тройную иерархию, а переплетения и слияния ветвей отражают процессы смешения при образовании антропологических типов. Символ схемы — дерево.
Классификация В.В. Бунака также имеет тройную иерархию, для нее характерен отказ от принципа дихотомии при ветвлении. Символ схемы — куст.
В свою классификацию В.В. Бунак ввел меру различий между иерархическими уровнями, которую он описывает следующим образом. «Для характеристики различий между формами современных гоминид английский морфолог Кизс (Keith, 1923) предложил термин, указывающий степень различимости («диакритичности») типов. Типы, различимые с «первого взгляда», т.е. состоящие из индивидуумов, несущих полный набор отличий группы, Кизс назвал «пандиакритическими». Если различимы не менее 80% индивидуумов, то тип называется макродиакритическим, при 50 % различимости тип определяется как мезодиакритический. Древние типы и типы, характерные для континентов, виды и стволы принадлежат к разряду пандиакритических, вести соответствуют макродиакритическим формам, подветви — мезодиакритическим, конечные разветвления — микродиакритическим» (с. 228—229).
Хрисанфова Е.Н., Перевозчиков И.В. «Антропология», 1991:
«...При большом количестве разных классификаций у исследователей существует некое «молчаливое» согласие по многим отдельным антропологическим типам и их группировке в более крупные подразделения. Практически во всех классификациях упоминаются следующие антропологические типы независимо от их ранга: европеоиды (евразийская, кавказоидная раса), среди которых обязательно выделяют светло и темнопигментированные, а также лопарский варианты; монголоиды (азиатскоамериканская) с американоидным, эскимоидным и дальневосточным типами; негроиды (экваториальная) с австралоидным, меланезийским, негритосским, собственно негрским, бушменским (южноафриканским, койсанским, капоидным) и пигмейским (негрильским, центральноафриканским) типами. В
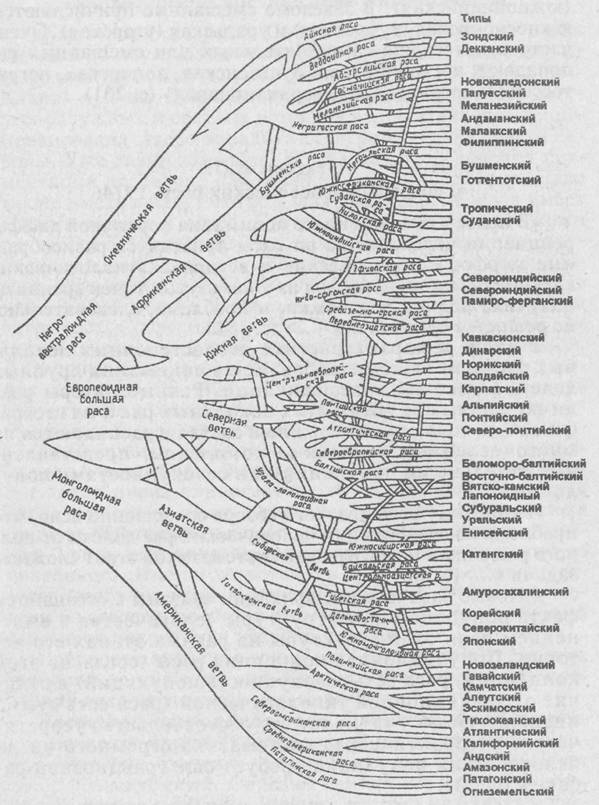
Рис. 99. Схема классификации человеческих рас по Г.Ф. Дебецу (1958). Переплетение и слияние ветвей отражают процессы смешения, имевшие место в различные периоды эволюции современного вида
самостоятельные варианты со спорным происхождением выделяются: полинезийская, айнская (курильская), эфиопская (восточноафриканская), веддоидная, дравидийская (южноиндийская). В заведомо смешанные причисляются: южносибирская (туранская) и уральская (угорская). Очень часто в категорию самостоятельных или смешанных рас попадают: австралийская, бушменская, лопарская, негритосская и негрильская, американоидная» (с. 231).
Алексеев В.П. «География человеческих рас», 1974:
«В пользу того или иного понимания первичной дифференциации человечества на расы приводятся разнообразные морфофизиологические и историко-этнологические аргументы, но пока ни одна из упомянутых точек зрения не получила полной поддержки, и проблема, следовательно, не решается однозначно <...> (с. 18).
В числе и характеристике второстепенных локальных рас не меньше разнобоя, чем в понимании крупных делений расовой классификации. Разные авторы различно понимают объем этих локальных рас и их географию. Они тоже имеют сложный состав и распадаются на многочисленные локальные комплексы, представленные отдельными популяциями и совокупностями популяций <...> (с. 20).
Из предыдущего краткого обзора совершенно ясно, что проблема классификации человеческих рас далека от Полного решения и перед расовой систематикой стоят сложные задачи <...>(с. 20).
...Новые задачи классификации связаны с осознанием факта популяционной структуры человечества и выяснением роли этой структуры на разных этапах его истории. Популяционная концепция расы (согласно этой концепции, раса есть совокупность популяций) в отличие от традиционной типологической (раса есть сумма индивидуумов) учитывает это обстоятельство теоретически, но практическая систематика огромного числа человеческих популяций требует еще грандиозной работы» (с. 20).
Хрисанфова Е.Н., Перевозчиков И.В. «Антропология», 1991:
Морфологическое описание «больших» рас
«Европеоидная раса. Цвет кожи варьирует от очень светлых до смугловатых оттенков. Популяции на север от широтной горной системы Пиренеи — Альпы — Балканы — Кавказ — Тянь-Шань имеют значительную частоту светлых радужных и светлых оттенков волос. Наиболее депигментирована северо-западная и северо-восточная части Европы. Уже давно замечено, что карта распределения пигментации по Европе напоминает карту оледенения. Надо также иметь в виду, что большая часть европеоидов имеет темные волосы и глаза. С этой точки зрения цвет кожи считается более важным признаком при объединении европеоидов в единую группу. Волосы чаще либо мягкие и прямые, либо волнистые. Курчавоволосость и узковолнистость чаще наблюдаются в южных районах, но спорадически встречаются повсеместно. Нос обычно со средним или высоким переносьем, узкий, прямой или с выпуклой спинкой. В северных популяциях частота вогнутых спинок носа может достигать 30—40%. Складка верхнего века развита значительно в северных группах и практически отсутствует в большинстве южных популяций. Разрез глаз горизонтальный (точнее, со слегка заметным поднятием наружного угла глаза). Горизонтальная профилировка от сильной (т.е. лицо очень клиногнатно) до умеренной. Последняя характеристика часто встречается в северо-восточных группах. Вертикальная профилировка лица ортомезогнатная. Третичный волосяной покров развит от сильного до умеренного. Иногда встречаются группы, где обволошенность (особенно тела) в среднем ослаблена. Они локализованы на севере и на юге, в промежуточной зоне третичный волосяной покров, как правило, сильный.
Групповые значения длины тела колеблются от высоких значений до нижесредних. Как уже говорилось, для пропорций тела характерна мезоморфия.
Австралонегроидная раса. Цвет кожи от очень темных до желтобурых (у бушменской расы) оттенков. Темные явно преобладают, но очень темных немного. Цвет волос и глаз темный. Описаны единичные случаи светлоглазых негров в Африке. Видимо, частота их близка к частоте мутаций. Есть популяции австралийских аборигенов, у которых с небольшой частотой встречаются светловоло
сые индивиды; в первую очередь дети (10—15%) и немногие молодые женщины. Небольшая тенденция к рыжеволосости описана у папуасов. Форма волос — от очень курчавых до широковолнистых (в австралоидной ветви). Нос со средневысоким или низким переносьем, часто очень широкий, маловыступающий (у папуасов бывает также сильное выступание с выпуклой спинкой). Губы часто с очень большой слизистой частью. Альвеолярная часть лица выступает вперед (прогнатна). Горизонтальная профилировка лица средняя или сильная <...> (с. 231—232).
Вариация в средних величинах длины тела очень значительна. В данной большой расе есть по крайней мере два пигмоидных типа (негрилли и негритосы). Есть и просто малорослые варианты (бушмены, папуасы и т.д.).
Монголоидная раса. Цвет кожи — от смуглого до светлого (в основном у североазиатских групп). Цвет волос темный, у некоторых вариантов очень темный (иссиня черный). Волосы, как правило, жесткие и прямые, но в Южной Азии есть группы со значительной частотой волнистых волос, в Северной Азии нередки мягкие волосы. Нос обычно довольно узкий, с малой или средней высотой переносья, выступает незначительно, но есть варианты с сильно выступающим носом. В целом нос по величине и форме значительно варьирует, в противоположность устоявшемуся стереотипу о «малоносности» монголоидов как характерной черте. Складка верхнего века развита значительно. Эпикантус может достигать частоты 90—95 %, но во многих группах (Америка, Южная Азия) он редок. Наружный угол глаза приподнят. Третичный волосяной покров развит слабо на лице и практически полностью отсутствует на теле.
Длина тела варьирует меньше, чем у негроидов, но по-настоящему высокорослых групп немного, как нет и очень малорослых» (с. 234).
Алексеев В.П. «Новые споры о старых проблемах», 1991:
Раса как морфофизиологическое понятие
«Основными критериями подразделения человечества на расы всегда были внешние особенности строения тела. Иными словами, с самого начала истории антропологии для выделения рас использовали морфологические признаки. Сами расы рассматривали как морфологические образова
ния. В рамках морфологического подхода было накоплено большое количество весьма ценных наблюдений над расовыми различиями.
Каковы характерные морфологические особенности монголоидов? Прямые жесткие черные волосы. Малое развитие бороды у мужчин и почти полное отсутствие волос на теле. Большое уплощенное лицо, слабо выступающий нос. Наличие эпикантуса, сильное развитие складки верхнего века. Значительный альвеолярный прогнатизм, т.е. выступающие вперед губы. По этому признаку монголоиды мало отличаются от негроидов. Правда, у них прогнатизм обусловлен только развитием мягких тканей, в то время как у представителей негроидной расы он зависит и от строения костной основы и зубов (что сильно заметно на негрских черепах).
Этот комплекс признаков варьирует в пределах разных локальных типов монголоидов, но в целом остается более или менее стабильным в границах ареала всей монголоидной расы, за одним исключением. Речь идет о так называемых американоидах, т.е. коренном населении Америки. У американских индейцев в отличие от азиатских монголоидов значительно выступает нос и отсутствует эпикантус. Эти два признака приближают их к европеоидам. Чтобы сохранить какую-то цельность морфологической характеристики монголоидной расы и придать ей какое-то единство, была предложена гипотеза, надо сказать, оправданная палеоантропологически. Предполагается, что в типе американоидов законсервирован древний первичный комплекс монголоидных особенностей, на основе которого и развились классические монголоиды.
Европеоидная раса отличается от монголоидной мягкими светлыми волосами, которые могут быть прямыми (как у северных народов Европы) или волнистыми (народы Южной Европы, Передней Азии и Северной Африки). У них светлее глаза и кожа, т.е. пигментация в целом. Резко профилированное и более узкое лицо, значительно выступающий нос с высоким переносьем. Именно у европеоидов наблюдается максимальное по лицевому масштабу выступание носа. Лицо у европеоидов ортогнатное, т.е. уплощено в вертикальной плоскости, у монголоидов — в горизонтальной, область рта не выступает вперед. Характеристика не абсолютна, ибо на Кавказе и Балканах проживают люди, ширина лица которых не намного меньше, чем у монголоидов. Во многих южных группах европеоидной расы заметны выступающие вперед губы. И в этих случаях также писали о сохранении протоморфных (первичных) вариан
тов в условиях изоляции, вызванной горной местностью, вариантов, исходных для европеоидной расы в целом.
Негроидная раса выделяется темной пигментацией — темно-коричневой, почти черной кожей, хотя цвет глаз и волос у негров, по-видимому, не темнее, чем у монголоидов. Курчавость свойственна всему волосяному покрову негров — волосы курчавятся не только на голове, но и на теле. Борода растет у мужчин, пожалуй, несколько сильнее, чем в пределах монголоидных групп, но значительно слабее, чем у европейцев. Общий лицевой и альвеолярный прогнатизм, очень широкий нос и толстые губы (толще, чем у монголоидов, а тем более европеоидов). Эти признаки дополняют своеобразие негроидного комплекса признаков.
Коренное население Австралии не уступает неграм Африки по развитию признаков всего этого комплекса. И конечно, обращает на себя внимание их волнистоволосость. Долгое время это рассматривалось как доказательство какого-то особого происхождения австралийцев. Однако затем было показано, что у папуасов и меланезийцев курчавоволосость появляется сравнительно поздно, на рубеже перехода от младенческого к детскому возрасту. Это наблюдение легко истолковать генетически как показатель общего происхождения волнистоволосых и курчавоволосых групп океанийской ветви негроидов.
Курчавоволосость папуасов и меланезийцев представляет собой, очевидно, позднее образование, результат вторичной адаптации. Позднее появление курчавоволосости африканских негров не наблюдали, но отсутствие наблюдений нельзя трактовать как свидетельство непременно разного происхождения африканских и океанийских негроидов. У тех и других светлая кожа на ладонях и подошвах, подмышками и в области промежности. У европейцев кожа подмышками и в промежности, наоборот, темнее, чем на всех остальных участках тела. Такое различие позволяет предполагать, что общие предки тех и других были нейтральны по пигментации, занимая промежуточное положение. По аналогии можно думать, что и волосы у них были ни курчавыми, ни прямыми, приближались по строению к волосам австралоидов. Интенсивная адаптация к условиям тропиков в Африке, по-видимому, способствовала более быстрой утере волнистоволосости, чем в Океании.
Морфологические характеристики рас не потеряли своего значения и в настоящее время <…>.
Сейчас углубляется собственно морфологический анализ. Постепенно накапливаются данные о генетике и воз
растной изменчивости расовых признаков. Морфология рас может быть исследована полнее и глубже, чем когда-либо. Однако бесспорно — вся совокупность накопленной к настоящему моменту информации стала недостаточной, ибо она не отражает уже и не исчерпывает всех наших знаний о человеческих расах <...> (с. 11—13).
В орбиту исследования были широко вовлечены генетические маркеры и определены их концентрации в разных популяциях <...> (с. 6).
Почти все коренное население Америки — носители группы О системы АВО и группы N системы MN. Коренное население Австралии в отличие от американских индейцев, наоборот, почти все носители группы М системы MN. Огромное число популяций Восточной Сибири фиксировано по гену D, т.е. у них отсутствует ген резусотрицательности. Таких примеров множество. <...>
Однако фиксированность того или иного гена (100%ное или нулевое присутствие его в популяции) — крайние случаи. Обычно на огромных территориях образуются и многочисленные расово-специфические концентрации тех или иных генетических маркеров. Классический пример — преимущественное накопление гена р (группа крови А) у населения Европы, относящегося к европеоидной расе, и гена q (группа крови В) у населения Азии, относящегося к монголоидной расе. Как оказалось, те или иные сочетания генов не менее типичны для разных рас, чем их морфологические признаки.
Накапливая сведения о вариациях генетических маркеров, мы столкнулись и с неожиданностями. Вскрылось немало случаев противоречия между собственно популяционно-генетическими и морфологическими данными, например, близость населения Индии и восточноазиатских монголоидов по преобладанию группы В над А. Для разумного объяснения всех подобных фактов потребуется большая дополнительная работа. Однако ее перспективы могут быть намечены уже сейчас. Она пойдет по линии усложнения существующей иерархии рас и выделения внутри известных сейчас расовых общностей дополнительных локальных вариантов. Увеличившееся богатство расовой характеристики, несомненно, обогатит и расовую классификацию человечества.
Наконец, нельзя не упомянуть и еще о ряде фактов (не связанных с популяционной генетикой), полученных буквально в последние годы. Они еще не полностью осмыслены, но чрезвычайно важны для нашей темы. Это факты, относящиеся к расовой физиологии в узком смысле слова. У ското
водов Восточной Африки — масаев, например, мясо и молоко, т.е. животные белки и жиры, составляют подавляющую часть рациона. Однако в крови у них меньше холестерина, чем у европейцев. Вероятно, этому способствовали какие-то пока не вскрытые адаптивные механизмы к пересыщенной животным белком и жиром диете. Появились сообщения о разном уровне проницаемости клеточных мембран у негроидов и европеоидов. Причем у негроидов она выше, что способствует, по-видимому, одноактным адаптивным сдвигам в обмене.
Вот так постепенно антропология наполнялась популяционно-генетическими и собственно физиологическими данными. Благодаря им усовершенствуются расовые характеристики и классификация человечества. Мы теперь знаем, что территориальная дифференциация человеческих популяций и их совокупностей — рас происходила не только по морфологическим, но и по физиологическим признакам. Иногда пишут, что расовые различия наблюдаются только в мелочах, в каких-то второстепенных внешних признаках. Однако это не так: Расовые различия охватывают и фундаментальные функциональные свойства. Человеческие расы — не узкоморфологические, а морфофизиологические категории» (с. 13—15).
Современное распространение человеческих рас
Области расселения современных человеческих рас
(по Рогинскому, Левину, 1978 г.):