Роль водорода и металлов в становлении и эволюции метаболических систем
| Вид материала | Документы |
- Роль Великого Новгорода в становлении российской государственности Роль Великого Новгорода, 210.52kb.
- Пятый физические основы сваривания металлов, 781.92kb.
- Концепция эволюции в биологии, 91.47kb.
- Реферат по теме: «Металлы. Свойства металлов.», 196.2kb.
- Автореферат диссертации на соискание ученой степени, 2303.74kb.
- Патофизиология эндокринной системы, 226.95kb.
- Алгоритмы и примеры решения клинических задач по патофизиологии нарушений кислотно-основного, 674.6kb.
- Контроля и управления процессом получения водорода, 27.17kb.
- Коррозия металлов, 201.83kb.
- Происхождение человека, 109.18kb.
Роль водорода и металлов в становлении и эволюции метаболических систем
М.А.Федонкин*
*Институт геохимии и аналитической химии им. В. И. Вернадского РАН
mfedon@paleo.ru
Предлагается гипотеза о ведущей роли водорода (источник протонов и электронов) и ионов металлов переходной группы (первичные катализаторы) в формировании энергетической основы жизни в условиях ранней Земли. Реконструированы доорганические стадии эволюции метаболических систем, вовлекавшие в качестве акцептора электрона неметаллы относительно большого ионного радиуса. Становление и ранняя эволюция метаболических систем живой клетки рассматривается как бутсрэпинг-процесс, компенсирующий необратимые изменения геохимических особенностей биосферы, в особенности, уменьшение доступности водорода и некоторых металлов переходной группы.
Ссылка: Федонкин М.А. Роль водорода и металлов в становлении и эволюции метаболических систем. В кн.: Проблемы зарождения и эволюции биосферы (Под ред. Э.М.Галимова). М.: Книжный дом «Либроком», 2008, с. 417-437.
1. Ведение
Идея о том, что неравновесность - поток энергии или вещества – формирует упорядоченные системы, по праву обретает статус научной парадигмы современного естествознания (Галимов, 2001; Пригожин, Стенгерс, 2003). Применительно к проблеме происхождения жизни эта идея направляет исследование в область процессов, которые описываются не столько языком химии, сколько в терминах физики. Универсальной физической характеристикой живого является формирование ионных градиентов и перенос электрона от донора к акцептору. Эта физическая основа энергетики метаболизма клетки допускает предположение о том, что процесс переноса энергии и был основным фактором упорядочения, приведшим к формированию динамически устойчивых молекулярных систем – предшественников биоорганических соединений. Существование таких систем в условиях ранней Земли было не только возможным, но, по-видимому, термодинамически выгодным в аспекте стабильности. Организующая (структурирующая) способность потока энергии проявлялась на всех стадиях становления биологических систем в той же мере, как она действует в современных явлениях жизни.
Физический подход к проблеме происхождения жизни с акцентом на энергетику обменных процессов (Morowitz, 1992; Hengeveld, 2007) позволяет использовать единый язык описании процессов неживой и живой природы. Историческая непрерывность жизни в части электрохимической природы метаболической активности клетки допускает возможность использования подхода от начальных условий (снизу вверх). Этот подход описывает развитие метаболизма и факторов среды как дополнительные и взаимозависимые процессы. Физический подход допускает проверку достоверности моделей данными биохимии, геохимии и палеобиологии. Все это указывает на ряд преимуществ физического подхода над химическим подходом, который до сих пор доминирует в области исследований проблемы происхождения жизни.
2. Подходы к проблеме происхождения жизни
Большая часть современных подходов к проблеме происхождения жизни ориентируется на процессы синтеза макромолекул, которые могли служить предшественниками биоорганических соединений. Наиболее детально разработаны модели, объединяемые в группу так называемых «минеральных гипотез», в значительной степени опирающихся на лабораторный эксперимент по синтезу макромолекул на минеральной подложке (Cairns-Smith et al., 1992; Ferris et al., 1996). Действительно, многие минералы обладают каталитическими свойствами, кристаллические решетки ряда минеральных видов могут служить фактором концентрации и сортировки (по размеру или симметрии) простых молекул и шаблоном для синтеза макромолекул органических соединений.
Критики указанной группы гипотез справедливо указывают на их слабые стороны: 1) минеральный кристалл – относительно стабильная, а значит, химически пассивная форма соединений; 2) в явлениях современной жизни минералы не играют ни одной из тех ролей, которые им приписываются в процессе становления биологических систем; 3) все органические молекулы синтезируются самой жизнью (Э. М. Галимов, 2007, устное сообщение). Дополнительную сложность представляет собой проблема исторической последовательности формирования липидов, белков, РНК, ДНК и других «строительных блоков» живого – реакции синтеза этих веществ в живой клетке настолько тесно связаны между собой, что формирование их порознь и самосборка в функционирующие цепи представляется маловероятной. Вообще, в рассуждениях об абиогенном синтезе биологически значимых макромолекул нередко упускается из виду весьма важное обстоятельство – динамическую стабильность живых систем, в том числе и на молекулярном уровне. При этом не придается значения тому, что все «строительные блоки» живого (белки, жиры, углеводы, РНК и ДНК) синтезируются самой живой клеткой. Существование «первичного бульона», насыщенного абиогенными органическими соединениями, которые сформировали в итоге живую клетку путем самосборки, представляется маловероятным прежде всего с термодинамической точки зрения, - подобная система неизбежно (и очень быстро) приходит в равновесие.
Несмотря на указанные недостатки, некоторые из «минеральных гипотез» происхождения жизни заслуживают дальнейшей разработки хотя бы потому, что минералы с их способностью структурировать пространство химических реакций были важнейшим фактором среды происхождения жизни (Юшкин, 2000), а также и потому, что химические элементы, составляющие эти минералы, могли в виде более простых соединений формировать системы накопления, сохранения и освобождения энергии. Представляется, что такие соединения должны были формироваться в области резких электрохимических градиентов. Именно в этом отношении интересны гидротермальные системы раннего океана.
Одна из наиболее популярных ныне гипотез происхождения жизни в среде океанических гидротерм развивается М. Расселлом и его соавторами (см. Russell, 2007 и ссылки в этой работе). Щелочные флюиды древнейших гидротерм выносили водород, сульфид и аммиак, а также ионы таких металлов, как железо и никель, сульфиды которых в изобилии осаждались вокруг источников. Микроскопические поры в минеральной корке таких сульфидов, как греигит, предоставляли полуизолированное пространство для синтеза простых органических молекул, которые формировали более сложные пептиды. Эти пептиды покрывали внутреннюю поверхность пор, что могло быть первым шагом к автономии клетки. Привлекательным аспектом этой гипотезы представляются отнюдь не микроскопические поры сульфидов – гипотетические предшественники клеток, но, прежде всего, - среда на переходе от гидротермальной системы к придонным водам океана, геохимические градиенты и высокая концентрация свободных ионов и электронов. Именно в такой среде могли формироваться молекулярные системы, как бы встраивающиеся в естественный поток энергии, концентрирующие его и способные к временному сохранению и освобождению энергии. Мы говорим о неорганических предшественниках ферментов. В этом отношении симптоматичным представляется сходство молекулярной структуры пирита и Fe-S кластеров в активных центрах многих ферментов или же сходство молекулярной структуры другого сульфидного минерала, греигита (Fe5NiS8), и тиокубана в составе молекулы белка ферредоксина (Russell, 2006). Это сходство можно интерпретировать как свидетельство происхождения жизни в среде гидротермальных систем, и вместе с тем, как намек на то, что первичными катализаторами были не белки, а их неорганические предшественники (Федонкин, 2007; Hengeveld, Fedonkin, 2007).
В поддержку гипотезы гидротермального происхождения жизни ( Forterre et al., 2002) нередко приводят факт базального положения гипертермофильных групп Archaea и Bacteria на молекулярном древе жизни (Stetter, 1996), хотя возможность позднейшей адаптации некоторых мезофильных прокариотных таксонов к агрессивной среде гидротерм, подобных черным курильщикам, исключить нельзя (Brochier et al., 2005).
3. Методология
Принцип исторической непрерывности эволюции материи от неживых объектов к живым (Опарин, 1924) дает нам основание рассматривать формирование энергетической основы жизни, связанной, прежде всего, с переносом электрона, на добиологической (и даже на доорганической!) стадии эволюции земного вещества в условиях хадейской Земли (Федонкин, 2007; Hengeveld, Fedonkin, 2007). С физической точки зрения происхождение жизни в среде гидротермальных систем или вблизи от места их разгрузки представляется плодотворной гипотезой в виду таких факторов, как насыщенность среды свободными электронами, резкий электрохимический градиент, обилие ионов металлов и молекулярного водорода.
В основе методологии исследований проблемы происхождения жизни, развиваемой в этой статье, лежит принцип сохранения энергетической преемственности и исторической непрерывности жизни от самых ранних стадий ее становления. Этот подход преодолевает кажущуюся пропасть между неживым (косным, по выражению В.И.Вернадского) и живым, которая неизбежно присутствует в доминирующих моделях, описанных в терминах химии. В настоящей работе мы следуем некоторым постулатам (см. Hengeveld, Fedonkin, 2003, 2007; Fedonkin, 2008): 1) происхождение жизни и ее эволюция определяется физическими и химическими параметрами среды; 2) начальная биохимия была теснейшим образом связана с геохимией внешних геосфер ранней Земли; 3) жизнь началась как процесс, не требующий большой энергии, в среде, богатой свободными электронами и протонами; 4) неорганическая стадия эволюции метаболизма предшествовала органической стадии истории живого; 5) неорганические катализаторы биохимических реакций были предшественниками белковых молекул ферментов; 6) металлы как основной источник электронов играли ключевую роль катализатора древнейших биохимических реакций; 7) физико-химические параметры биосферы необратимо удалялись от начальных условий происхождения жизни; 8) сохранение функциональности архаичных метаболических модулей клетки в изменяющихся условиях среды осуществлялось путем надстраивания метаболических каскадов дополнительными модулями, функционально сопряженными с новыми факторами среды; 9) структура метаболических сетей клетки и структура молекул ферментов допускает причинно-историческую интерпретацию; 10) метаболические каскады живой клетки рекапитулируют биохимическую эволюцию и отражают исторические изменения геохимических факторов среды; 11) если последнее справедливо, то анализ особенностей передачи энергии по метаболическим цепям открывает возможность реконструкции происхождения жизни (прежде всего в физическом и историческом аспектах).
Основу жизнедеятельности клетки составляют реакции двух типов: а) окислительно-восстановительные реакции, связанные с переносом электрона от донора к акцептору и б) кислотно-щелочные реакции фосфатов, связанные с переносом протона. В живой клетке сигнал (импульс энергии) усиливается каскадами биохимических реакций, в которых один фермент, будучи продуктом предшествующей каталитической реакции, в свою очередь катализирует формирование другого фермента, который катализирует синтез последующего и т.д. Исследуя природу энергетической основы метаболизма клетки, мы должны принимать во внимание важное различие живого и неживого в отношении энергии: в неживых системах энергия используется прямо, тогда как в живых системах энергия временно сохраняется (в виде энергии химических связей или ионных градиентов) в промежутке времени между моментом ее поступления и освобождения (Ho, 1995). Сохранение энергии представляется первым процессом после ее генерирования, прежде чем начать использование этой энергии в таких процессах, как рост, поддержка постоянства внутренней среды, размножение, движение и др. (Kooijman, Hengeveld, 2005). Эта важнейшая и универсальная особенность живого направляет наше внимание к обратимым, циклическим реакциям, протекавшим в геохимических обстановках ранней Земли. Такие универсальные механизмы жизнедеятельности клетки, как перенос электрона и протона, накопление, сохранение и освобождение энергии, по-видимому, сформировались очень рано, возможно, на доорганических стадиях становления живого (Hengeveld, Fedonkin, 2007) и сохранялись на протяжении миллиардов лет вопреки радикальным изменениям среды. Вся биологическая эволюция может интерпретироваться как результат отбора в пользу более надежных способов сохранения эффективности базовых типов метаболизма, сформировавшихся в условиях ранней Земли, в ответ на необратимые изменения физико-химических параметров биосферы.
Сложность организации живой клетки не допускает предположения о возможности ее формирования единым актом самосборки: клетка является результатом длительного, многошагового исторического процесса, который можно отнести к категории бутстэпинг-процессов (Hengeveld, Fedonkin, 2007): увеличение метаболической сложности и эффективности клетки происходило при сохранении базовых (древнейших) механизмов жизнедеятельности. Хотя многие детали эволюционного бутстрэпинг-процесса еще предстоит установить, уже сейчас представляется необходимым выделять три его последовательных стадии. Первая стадия становления живого вовлекала тяжелые неметаллы нижних периодов Периодической системы элементов, например, селен. Эти неметаллы с большим ионным радиусом формировали легко диссоциирующие соединения, которые не требовали большой энергии и специальных катализаторов для их расщепления. Реакция неметаллов с водородом катализировалась металлами переходной группы (W, Fe, Ni). Гидрогенация и дегидрогенация молекул определяется обменом электронов в ходе редокс-реакции. Металлы и соответствующие реакции присущи кофакторам многих ферментов. На втором этапе система дополнялась кислотно-щелочными реакциями, формирующими цикл конденсация-гидролиз с активным вовлечением неметаллов более высоких периодов, например, серы и фосфора. Поскольку реакции конденсации ведут к полимеризации, на этом этапе могли формироваться не только липиды, но и нуклеотидные коэнзимы, начиная с молекулы АТФ и заканчивая многими типами РНК, а в дальнейшем и ДНК. Третий этап усложнения метаболических систем был связан с вовлечением еще более легких химических элементов, имеющих более высокую энергию связей (углерод, азот, и, наконец, кислород), синтезом протеинов и использованием солнечной энергии для фотолиза молекул H2S, а позднее и H2O. Изначальный избыток генерируемой энергии требовал формирования биохимических систем ее хранения и освобождения – молекул углеводов. Реконструкция определяющих факторов становления метаболических систем клетки требует данных об условиях среды формирования последних.
4. Проблема среды происхождения жизни
Физико-химические параметры ранней Земли и ближайшего космоса более 4 млрд. лет назад радикально отличались от современных. Данные наук о Земле, сравнительная планетология и астрофизика проливают некоторый свет на особенности среды становления жизни. Некоторое представление об этом можно составить на основе данных экологии и физиологии эубактерий и архей, судя по их невероятной толерантности к воздействию разнообразных факторов (высокие дозы радиации, широкий диапазон температуры, давления, рН и концентрации растворенных соединений) и по их колоссальному биохимическому разнообразию. Некоторые биотопы экстремофилов, например, упоминавшиеся выше высокотемпературные гидротермальные системы типа «черных курильщиков», продуцирующие большие объемы CO2, H2S и металлов, или среднетемпературные (40-90С°) высокощелочные обстановки гидротерм Lost City, испускающие СН4 и Н2, рассматриваются в качестве основы моделирования среды происхождения жизни (Boetius, 2005; Kelly et al., 2005; Proskurowski et al., 2008). Подобные обстановки были распространены более широко на ранней Земле.
Реконструкции параметров древней биосферы посвящена обширная литература (см. Галимов, 2001; Сорохтин, Ушаков, 2002; Schopf, Klein, 1992; Knoll, 2003). Раннюю Землю отличали следующие особенности: 1) высокая фоновая радиация; 2) мощный поток радиогенного тепла (по некоторым данным, более чем на порядок превышавшего современный уровень, в основном, за счет энергии распада короткоживущих изотопов); 3) высокий вклад близкой Луны в тепловой бюджет Земли за счет механического разогрева недр; 4) интенсивный вулканизм, океаны лавы; быстрое формирование металлического ядра планеты (более 90% его массы в течение первых 100 млн. лет после аккреции); 5) рано сформировавшееся магнитное поле с его функцией эффективной защиты планеты от космического излучения; 6) полная переработка первичной коры; 7) низкий рельеф поверхности; 8) плотная восстановительная вторичная атмосфера с мощным парниковым эффектом, который компенсировал низкую светимость Солнца (на 30% ниже современного уровня); 9) практически полное отсутствие свободного кислорода и озонового экрана в атмосфере; 10) мелкий обширный океан, формировавшийся по мере того, как температура поверхности Земли опускалась ниже точки кипения воды; 11) высокая концентрация растворенных вулканических газов и ионов тяжелых металлов в древнем океане. Перечисленные факторы и формировали среду становления жизни.
Французский биолог Р. Дюбуа оставил замечательную метафору: «Жизнь – это одушевленная вода». Действительно, вода – это не только растворитель, агент гидролиза и среда химических реакций, а также преобладающий компонент химического состава живой клетки. В аспекте энергетики метаболизма вода – важнейший источник H2, e-, H-, H., OH-, H3O+, H+, H2O2, O2 и O3, играющих главную роль в процессах жизнедеятельности (Russell, 2007). Проблемы происхождения гидросферы Земли активно обсуждаются (см. обзор Marty, Yokochi, 2006). Наиболее ранними косвенными свидетельствами воды на Земле являются эродированные детритовые цирконы (4.4 млрд. лет) в Западной Австралии (Wilde et al., 2001; Harrison et al., 2005), высокие отношения O18/O16 в кристаллах циркона хадейского возраста (Mojzsis et al., 2001) и данные титановой термометрии по древнейшим цирконам (Trail et al., 2007). Древнейшие слоистые железные руды, подушечные базальтовые лавы и другие структуры подводных излияний в разрезах вулканогенно-осадочных толщ западной Гренландии и северного Квебека более определенно указывают на существование гидросферы 3800 млн. лет назад (Cates, Mojzsis, 2007). Облегченный изотопный состав графитовых включений в кристаллах апатита из супракрустальных пород (метаосадков) формации Акилиа (возраст более 3830 млн. лет) западной Гренландии интерпретируется как возможное древнейшее свидетельство жизни на Земле (McKeegan et al., 2007). Если это предположение справедливо, то процессы становления жизни происходили до начала формирования древнейших известных пород и не оставили следов в геологической летописи.
5. Биологическая роль водорода
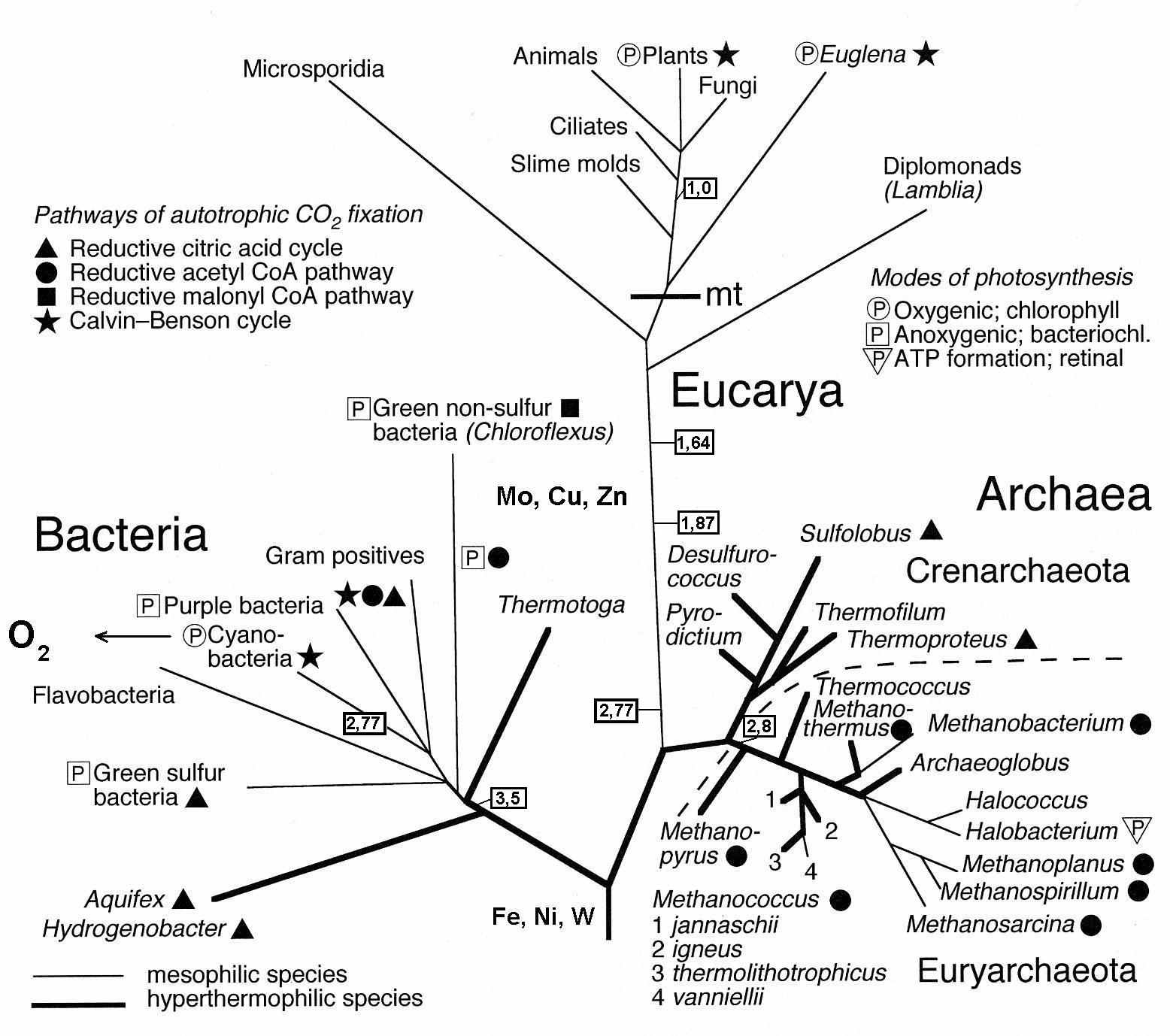
Исследуя энергетический аспект происхождения жизни, следует обратить внимание на водород как первичный источник электронов и протонов – основы энергии живого. Будучи самым распространенным химическим элементов Вселенной, водород был и остается главным субстратом микробной жизни и основной энергетики метаболизма (Wackett et al., 2004). Биологическая роль водорода связана не только с тем, что в массе живой клетки преобладает вода (H2O). Мягкие водородные связи придают прочность и пластичность макромолекулам. Различные микробные ферменты осуществляют перенос H+ катиона. Градиент H+ используется клеткой в процессе синтеза АТФ. Анион водорода H- известен как «энергетическая валюта» клетки (эквивалент двух электронов). Молекулярный водород H2 как ключевой промежуточный продукт реакций анаэробного метаболизма фактически осуществляет универсальную роль трофической (энергетической) связи между микроорганизмами, живущими на разных субстратах – чрезвычайно важная (системообразуюшая, экосистемная) особенность с момента возникновения жизни. Многие группы прокариот используют H2 в качестве донора электрона и источника энергии, и фактически конкурируют за доступ к водороду: хемолитоавтотрофы, хемолитогетеротрофы, фотолитоавтотрофы и фотолитогетеротрофы. Водородный метаболизм доминирует среди микроорганизмов, населяющих среду гидротермальных систем и глубоких водоносных горизонтов. Некоторые реакции обмена, осуществляемые хемоавтотрофными термофильными археями (например, реакция FeS+H2SFeS2+H2 + свободная энергия), рассматриваются как прототип или аналог первых шагов к становлению хемиосмотического потенциала и синтеза АТФ. Ряд прокариот, в частности, сульфатредукторы, пурпурные несерные бактерии, метаногены, живущие во внутриклеточном симбиозе с эвкариотами, служат метаболическим резервуаром для стока Н2 (Fenchel et al., 1998).
Способности молекулярного водорода служить восстановительным фактором, быть источником энергии и формировать протонные градиенты как средство временного сохранения энергии показывают, что доступность водорода на ранней Земле была ключевым фактором не только древнейших типов метаболизма (Hoehler, 2004), но и происхождения жизни (Fedonkin, 2007, 2008). В системе рассуждений о первичной роли водорода в энергетике древнейших биологических систем нам представляются важными следующие обстоятельства: 1) высокая диффузионная подвижность водорода, его способность проникать в молекулярном и ионном виде через металлы, стекло и другие твердые вещества; 2) низкая энергия активации Н2; 3) формирование протонов и электронов при контакте молекулярного водорода с поверхностью железа; 4) прямое использование протонов протонным насосом АТФ-азы для синтеза АТФ; 5) широчайшее использование протонов и электронов во множестве биохимических реакций клетки. Биохимические реакций живой клетке по большой степени является электрохимическими, и прокариоты демонстрируют в этом отношении выдающиеся способности. Они как бы встраивают свои метаболические системы в электрохимические градиенты среды. Высокая зависимость микроорганизмов от электрохимических реакций может указывать на характеристики среды происхождения жизни на ранней Земле (Lane, 2006). Фундаментальные различия физиологии прокариот и эвкариот, прежде всего в отношении энергетики метаболизма, можно интерпретировать в пользу гипотез о хемоавтотрофном происхождении жизни (Martin, Russell, 2003). Фотосинтез, включая более простые формы аноксигенного фотосинтеза, сформировался позже.
Огромная часть метаболических реакций ныне живущих прокариот вовлекает молекулярный водород или простые летучие соединения водорода в качестве источника энергии. Виды метаболизма метаногенов (Н2/CO2) или других прокариот (H2/SO2), широко распространенные в микробных сообществах гидротермальных систем океанского дна (Staley, 2006), могут указывать на то, что становление жизни на ранней Земле было связано с реакциями молекулярного водорода с такими окислителями, как двуокись углерода или двуокись серы. Но есть ли надежные основания предполагать высокое содержание водорода в атмосфере ранней Земли?
6. Абиогенные источники водорода на Земле
Хондритовые гипотезы формирования Земли предполагают, что ранние вулканические газы были обогащены метаном и молекулярным водородом (Javoy, 1995). Эти предположения подтверждаются данными лабораторных экспериментов. Хондритовые метеориты при высокой температуре и высоком давлении испускают преимущественно H2, CH4 и NH3, что указывает на вероятность высокого содержания именно этих газов в ранней атмосфере Земли (Schaefer, Fegley, 2006). Независимые исследования первых стадий формирования металлического ядра нашей планеты (Kadik, Litvin, 2007) показывают, что этот процесс протекал в восстановительных условиях, обусловленных природой первичного земного материала, и сопровождался эмиссией CH4, H2, NH3 и, в меньшей степени, H2O в атмосферу. Принимая во внимание современные представления о высокой скорости формирования металлического ядра Земли – 95% его массы сформировалось в течение первых 100 млн. лет после аккреции нашей планеты (Galimov, 2005), можно предположить, что эмиссия водорода и его простых летучих соединений была чрезвычайно высокой на самых ранних этапах геологической истории.
Некоторые модели древней атмосферы Земли (до появления жизни) показывают вероятность высокой концентрации H2, превышающей на 3-4 порядка его концентрацию в современной атмосфере (Hoehler, 2004, 2005). Несмотря на быструю потерю этого легкого газа, водород мог составлять до 30% древней атмосферы (Tian et al., 2005). Точные оценки концентрации Н2 в древнейшей атмосфере Земли затруднительны, но в аспекте рассуждений о роли водорода в происхождении и ранней эволюции жизни более важной представляется вероятность его высокой концентрации в составе газов, растворенных во флюидах земной коры, гидротермальных, поровых и грунтовых водах ранней Земли в виду низкой скорости миграции и ограниченного выхода во внешнюю среду.
В качестве источников абиогенного водорода указываются: 1) реакции между растворенными газами в системе C-H-O-S в магматических расплавах, особенно, в основных и ультраосновных магмах; 2) диссоциация метана CH4 на углерод (графит) и молекулярный водород H2 при температуре >600°C; 3) реакция между CO2, H2O и CH4 при повышенной температуре пара; 4) радиолиз воды под воздействием энергии распада радиоактивных изотопов урана, тория, их дочерних элементов, и короткоживущих изотопов калия, железа и др. 5) катализ ряда силикатов при высоком давлении в присутствии воды; 6) гидролиз железистых минералов мафических и ультрамафических пород; 7) дегазация верхней мантии, которая освобождала нейтральные или слабо кислотные флюиды, насыщенные H2, CH4, H2S и CO2; 8) фотолиз - диссоциация молекул воды под воздействием энергии излучения ультрафиолетового диапазона; 9) бомбардировка альфа-частицами газообразных углеводородов в недрах Земли (Apps, van de Kamp, 1993; Morita, 1999; Fedonkin, 2007). Известный факт, что никель в твердой фазе и в расплаве способен поглощать большие количества водорода (до 17 объемов), образуя твердый раствор, также может иметь отношение к рассматриваемой здесь проблеме.
На ранней Земле все перечисленные абиогенные источники водорода были гораздо более мощными, чем ныне, прежде всего по причине высокой интенсивности теплового потока. Водородная составляющая вулканических эмиссий была очень высокой, пока мантия оставалась восстановленной, и снизилась после достижения мантией окисленного состояния около 3,9 млрд. лет назад почти до современного уровня (Hoehler, 2005). Преимущественно ультрамафический характер древней недифференцированной коры делал серпентинизацию одним из главных источников водорода. В современной биосфере серпентинизация изверженных пород, протекающая при температуре 90-400°С и высоком значении рН, остается важнейшим источником Н2. Этот широко распространенный процесс преобразования вулканитов, идущий с образованием магнетита, гидроокислов и серпентиновых минералов, высвобождает молекулярный водород, который служит энергетическим субстратом жизни для разнообразных хемосинтезирующих организмов (Schulte et al., 2006). Высокотемпературные магматические очаги при низком рН также вносят существенный вклад в водородный бюджет современных экосистем.
Несмотря на обилие генерируемого водорода, его содержание в большинстве сред весьма низкое, что объясняется не столько его легкой диффузией, сколько активным поглощением прокариотами. Присутствие метана в среде обычно является результатом водородного метаболизма. Особенно активно поглощают водород хемолитоавтотрофные гипертермофилы, живущие за счет энергии следующих реакций: 2H2 + O2 2H2O; H2 + NO3- NO2- + H2O; H2 + S° H2S; 4H2 + SO42- S2- + 4H2O; 4H2 + CO2 CH4 + 2H2O (Stetter, 1995). Широчайшее распространение водородного метаболизма среди прокариот, представляющих нижние (наиболее древние) ветви молекулярного древа жизни (рис. 1), предполагает, что водород как источник энергии использовался очень рано в истории живого. Абиотические источники водорода на древней Земле были мощнее, чем ныне, реактивный резервуар атмосферы для стока Н2 был слабее.