
Модуляция пектинами проницаемости кишечной стенки и иммунного ответа на овальбумин 03. 00. 13 физиология
| Вид материала | Автореферат |
- Кровотечения. Кровотечение, 46.13kb.
- «Владивостокский государственный медицинский университет», 470.77kb.
- Тема № Методы диагностики туберкулеза. Знанятие № Туберкулинодиагностика, 89.55kb.
- Жирков Анатолий Михайлович доктор медицинских наук профессор Симбирцев Андрей Семенович, 418.79kb.
- Под кровотечением понимают истечение крови из кровеносных сосудов при нарушении целостности, 444.5kb.
- Аномалии иммунного ответа, 360.45kb.
- Место для шифра, 117.79kb.
- Тема «Внутренняя политика Александра, 36.52kb.
- Актуальность темы, 509.67kb.
- Постановление Правительства Российской Федерации от 05. 11. 97 г. N 1387 «О мерах, 917.38kb.
На правах рукописи
ХРАМОВА ДАРЬЯ СЕРГЕЕВНА
МОДУЛЯЦИЯ ПЕКТИНАМИ ПРОНИЦАЕМОСТИ КИШЕЧНОЙ СТЕНКИ И ИММУННОГО ОТВЕТА НА ОВАЛЬБУМИН
03.00.13 – физиология
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Сыктывкар – 2009
Работа выполнена в лаборатории молекулярной физиологии и иммунологии Отдела молекулярной иммунологии и биотехнологии Учреждения РАН Института физиологии Коми научного центра Уральского отделения Российской академии наук (заведующий отделом к.б.н., доцент С.В. Попов).
Научный руководитель: кандидат биологических наук, доцент
Попов Сергей Владимирович
Научный консультант: академик
Оводов Юрий Семенович
Официальные оппоненты: доктор медицинских наук, профессор,
заслуженный деятель науки РФ
Добродеева Лилия Константиновна
доктор биологических наук, ст.н.с.
Головко Владимир Александрович
Ведущая организация: Учреждение РАН Институт иммунологии и физиологии УрО РАН
(г. Екатеринбург)
Защита состоится «24» декабря 2009 г. в 10 ч. на заседании диссертационного совета Д 004.017.01 при Учреждении РАН Институте физиологии Коми НЦ УрО РАН по адресу: 167982, Республика Коми, Сыктывкар, ул. Первомайская, 50.
С диссертацией можно ознакомиться в библиотеке Учреждения РАН Института физиологии Коми НЦ УрО РАН по адресу: 167982, Республика Коми, Сыктывкар, ул. Первомайская, 50.
Автореферат разослан « » ноября 2009г.
Ученый секретарь
диссертационного совета
кандидат биологических наук Н.Г. Варламова
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность исследования. Регуляция иммунного ответа на пищевые белки является актуальной проблемой иммунофизиологии. Иммунный ответ на антигены пищи зависит от барьерной функции кишечника, которая является важнейшим механизмом, препятствующим проникновению в кровь макромолекул пищевых белков (Kraehenbuhl et al., 1997). В последние десятилетия получены данные, свидетельствующие о том, что небольшие количества белков проникают из просвета кишки в кровь в непереваренном виде путем эндоцитоза (Udall et al., 1981; Bendayan et al., 1990; Ziv, Banyana, 2000; Bruneau et al., 2003; Cloutier et al., 2006; Cammisotto et al., 2007). Показано, что проникновение в кровь овальбумина (OVA) в высокомолекулярном виде в количестве, составляющем 0.001 – 0.01% от введенного белка, не препятствует развитию оральной толерантности к этому белку (Bruce, Ferguson, 1986; Friedman, 1996). Известно, что повышение проникновения белковых антигенов в кровь стимулирует иммунный ответ на эти антигены. Так, адъювантное действие бактериальных токсинов обусловлено увеличением транспорта антигенов в кровь (Kosecka et al., 1994; Rappuoli et al., 1999), а аллергические реакции на сою связывают с повышением проникновения белков соевого боба в кровоток (Weangsripanavalу et al., 2005). Повышение проницаемости кишечной стенки наблюдается при различных патологических состояниях (Hollander et al., 1986; Benard et al., 1996; Kiliaan et al., 1998; Лусс, 2003). При этом модулировать кишечную проницаемость могут медиаторы воспаления, выделяемые активированной тучной клеткой (гистамин, брадикинин) (Stein et al., 1998), цитокины (фактор некроза опухоли - ФНО-, интерферон - ИФН-, интерлейкины 1 и 4 - ИЛ-1 и ИЛ-4) (Berin et al., 1999; Perdue 1999; Ma et al., 2004; Al-Sadi, Ma, 2007).
Однако в литературе отсутствуют данные об изменении проницаемости кишечной стенки и иммунного ответа на пищевые белки при их совместном поступлении с компонентами питания. Известно, что пектины, входящие в состав продуктов питания и широко применяемые в пищевой промышленности, присутствуют в пищевом рационе человека наряду с белками (Скальный и др., 2005) и обладают иммуномодулирующим действием (Sakurai et al., 1999; Inagaki et al., 2001; Kiyohara et al., 2002; Vos et al., 2007; Оводов и др., 2009). Способность пектинов модулировать активность тучных клеток (Sawabe et al., 1992; Kobayashi et al., 2004) и секрецию цитокинов иммунными клетками (Inngjerdingen et al., 2007б; Guo et al., 2000), а также взаимодействовать со слизистой кишечника (Liu et al., 2005) определяет их возможность изменять проницаемость кишечной стенки для белковых антигенов. Однако до сих пор не изучена проницаемость стенки кишки и развитие иммунного ответа на белковый антиген, поступающий с пектинами. Не исследована секреция цитокинов, модулирующих кишечную проницаемость, у животных, получающих белок с пектиновыми полисахаридами. Слабо охарактеризовано действие пектинов на протеолиз белков, между тем расщепление белка в желудочно-кишечном тракте (ЖКТ) влияет на его молекулярные характеристики и тем самым может изменить его способность проникать через кишечный барьер.
Выяснение механизмов действия пектинов на проницаемость кишечной стенки и иммунный ответ на белковые антигены важно для регулирования барьерной функции и иммунитета кишечника.
Цель исследования - изучить проницаемость кишечной стенки для OVA и иммунный ответ на белок, поступающий с пектиновыми полисахаридами.
Для достижения цели исследования поставлены следующие задачи:
- Изучить проницаемость кишечной стенки для OVA, введенного перорально с пектиновыми полисахаридами.
- Определить содержание цитокинов в слизистой тонкой кишки мышей после введения OVA с пектинами.
- Изучить способность макрофагов секретировать цитокины у мышей, получивших OVA с пектинами.
- Установить действие пектинов на протеолиз белка in vitro.
- Исследовать развитие иммунного ответа на белковый антиген у мышей, получивших OVA с пектинами.
Научная новизна. Получены новые данные об иммуностимулирующем действии пектиновых полисахаридов. Впервые установлено, что проницаемость стенки кишечника для белкового антигена изменяется при поступлении белка с пектинами. Выявлены пектиновые полисахариды как повышающие (лемнаны, цирсиуман и цитрусовый пектин), так и снижающие (бадан, комаруман, гераклеуман) проницаемость кишечной стенки для OVA. Установлено, что модулирование пектинами проникновения белка обусловлено их влиянием на секрецию цитокинов и протеолиз белковой макромолекулы. Впервые показано, что пектины, повышающие проникновение OVA в кровь, стимулируют иммунный ответ на белок и ингибируют оральную толерантность к OVA. Показано, что фрагменты разветвленной области макромолекулы лемнана и цирсиумана обладают иммуностимулирующим действием.
Научно-практическая значимость. Расширены представления о модуляции проницаемости кишечной стенки и иммунного ответа на пищевые белки, а также о связи между структурой и иммуностимулирующей активностью пектинов. Получены экспериментальные данные, свидетельствующие о том, что иммуностимулирующее действие пектинов может быть обусловлено повышением проницаемости кишечной стенки для антигенов. Эти результаты могут быть использованы для быстрого тестирования растительных полисахаридов с целью выяснения их иммуностимулирующей активности. Способность пектинов повышать проникновение совместно вводимого с ними белка необходимо учитывать при составлении диет для пациентов, страдающих пищевыми аллергиями. Пектины, стимулирующие иммунный ответ на совместно вводимый с ними антиген, могут представлять интерес при создании пероральных вакцин.
Положения, выносимые на защиту:
- Проницаемость кишечной стенки для белкового антигена изменяется при поступлении белка с пектиновыми полисахаридами.
- Увеличение проникновения белка, вызванное пектинами, приводит к повышению иммунного ответа на антиген.
- Модулирование пектинами проницаемости кишечной стенки для белка связано с изменением секреции цитокинов и протеолиза белковой макромолекулы.
Апробация работы. Результаты исследований доложены и обсуждены на Всероссийской конференции молодых ученых, посвященной памяти профессора Н.Н.Кеворкова «Иммунитет и аллергия: от эксперимента к клинике» (Пермь, 2006), XIII Молодежной научной конференции Института биологии Коми НЦ УрО РАН «Актуальные проблемы биологии и экологии» (Сыктывкар, 2006), VI Молодежной научной конференции Института физиологии «Физиология человека и животных от эксперимента к клинической практике» (Сыктывкар, 2007), 14-ом Европейском симпозиуме по углеводам “Eurocarb 14”, (Любек, Германия, 2007), I Всероссийской молодежной научной конференции «Молодежь и наука на севере» (Сыктывкар, 2008), V Всероссийской школе-конференции «Химия и технология растительных веществ» (Уфа, 2008).
Публикации. По теме диссертационной работы в соавторстве опубликовано девять печатных работ (из них четыре опубликованы в изданиях, рекомендованных ВАК).
Объем и структура диссертации. Диссертация изложена на 129 машинописных страницах, состоит из введения, четырех глав (обзор литературы, материалы и методы исследования, результаты исследования, обсуждение результатов), выводов и списка литературы (192 источника). Диссертация содержит 20 таблиц и 6 рисунков.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Объект исследования. Работа выполнена на белых лабораторных мышах обоего пола массой 20-25 г, животные содержались в стандартных условиях вивария на лишенной животного белка диете. Доступ к корму и воде ad libitum. Проведение экспериментов на животных выполняли в соответствии с рекомендациями Комитета по биоэтике Учреждения РАН Института физиологии Коми НЦ УрО РАН.
Характеристика пектинов, используемых в работе. В работе использованы коммерческие препараты цитрусового (PC, MP Biomedicals) и яблочного (PA, Sigma) пектинов, а также пектины, выделенные из следующего растительного сырья: ряски малой Lemna minor L., (лемнан LM), каллуса ряски малой (лемнан LMC), стеблей бодяка съедобного Cirsium esqulentum Siev. (цирсиуман СЕ), бадана толстолистного Bergenia crassifolia L. (бергенан BC), борщевика сибирского Heraclеum sibiricum L. (гераклеуман HS), сабельника болотного Comarum palustre L. (комаруман CP). Выделение пектиновых полисахаридов и установление их химической структуры проведено в лаборатории гликологии Отдела молекулярной иммунологии и биотехнологии Учреждения РАН Института физиологии Коми НЦ УрО РАН. Пектины растворяли в фосфатно-буферном растворе (PBS) непосредственно перед введением животным. В работе исследовали пектины, имеющие разный тип построения углеводной цепи. Бергенан BC, значительная часть макромолекулы которого представлена метилэтерифицированным линейным галактуронаном; цирсиуман CE, в состав макромолекулы которого входят линейный галактуронан и разветвленный рамногалактуронан I; гераклеуман HS, главным компонентом макромолекулы которого является линейный галактуронан; комаруман CP, в состав которого входят разветвленный и линейный галактуронан, а также разветвленный рамногалактуронан I; лемнаны LM и LMC, в состав которых входят линейный галактуронан, апиогалактуронан и разветвленный рамногалактуронан I.
Определение концентрации белка в крови. Для определения количества OVA (Appli Chem), проникающего через стенку кишки, мышам перорально при помощи полиэтиленового зонда вводили белок в дозах 50, 250, 500 и 1000 мг/кг. Содержание белка определяли через полчаса, один, три, шесть часов и спустя сутки после введения OVA. При помощи кардиопункции у мышей забирали кровь, центрифугированием (4000 об/мин, 20 мин) отделяли сыворотку крови, в которой измеряли концентрацию OVA. Для изучения проникновения в кровь белка, поступающего с пектинами, мышам перорально вводили смесь OVA (доза 1000 мг/кг) и пектинов (доза 50 мг/кг). Контрольные животные получали только белок (доза 1000 мг/кг). Концентрацию OVA в крови измеряли с помощью конкурентного иммуно-ферментного анализа (ИФА), используя поликлональные анти-OVA-антитела (Кэтти, Райкундалиа, 1991).
Определение концентрации цитокинов в слизистой тонкой кишки. Через три часа после введения белка или белка с пектинами у мышей изолировали тонкую кишку. Содержимое тонкой кишки и слизистую соскабливали и собирали в пробирки, взвешивали, гомогенизировали в 1 мл PBS и перемешивали в течение 30 минут. Гомогенат слизистой центрифугировали 10 минут при 1000×g. Надосадочную жидкость делили на аликвоты и замораживали для дальнейшего определения в них цитокинов: ФНО-, ИЛ-1 и ИФН-. Измерение концентрации цитокинов проводили при помощи сэндвич ИФА, используя моноклональные и биотилированные анти-ФНО--, анти-ИЛ-1- и анти-ИФН--антитела (Кэтти, Райкундалиа, 1991).
Оценка продукции цитокинов перитонеальными макрофагами. Для определения секреции цитокинов (ФНО- и ИФН-) макрофагами перитонеального смыва использовали общепринятый способ активации лейкоцитов и получения смыва брюшной полости (Клаус, 1990). Суспензию лейкоцитов (100 мкл), содержащую 2×106 клеток в питательной среде RPMI 1640 с добавлением 10% фетальной сыворотки, вносили в лунки плоскодонных микрокультуральных планшетов и инкубировали в CO2-инкубаторе в течение 2 часов при 37 °C. Неадгезированные лейкоциты удаляли, трижды отмывая планшеты питательной средой. Микроскопическим путем было установлено, что монослой клеток, адгезированных на пластике, содержит преимущественно макрофаги. Далее в лунки вносили по 100 мкл свежей питательной среды, содержащей 10% фетальной сыворотки, -меркаптоэтанол, антибиотики и липополисахарид грамотрицательных бактерий (1 мг/л), необходимый для стимулирования макрофагов. Образцы инкубировали в CO2-инкубаторе в течение 24 часов при 37 °C, после чего планшеты центрифугировали (800×g, 10 мин) и отбирали надосадочную жидкость клеточной суспензии, которую затем делили на аликвоты и замораживали (Petursdottir et al., 2002) для последующего определения в них концентрации цитокинов (ФНО- и ИФН-) с помощью сэндвич ИФА, используя моноклональные и биотилированные анти-ФНО- - и анти-ИФН--антитела (Кэтти, Райкундалиа, 1991).
Протеолиз белка в присутствии пектинов in vitro. OVA подвергали протеолизу, используя схему, разработанную ранее (Mouecoucou et al., 2006). Белок обрабатывали пепсином и соляной кислотой, а затем протеолитическими ферментами поджелудочной железы (трипсином и химотрипсином). Смесь белка и пектинов (пектин борщевика – гераклеуман HS, цитрусовый пектин) в отношении 1:1 готовили на фосфатном буфере (pH-7.3) и оставляли на ночь при 4 ºC для агрегации молекул. Затем к смеси добавляли пепсин (соотношение фермент/субстрат: 1/125), инкубировали при 37 ºC. При этом в течение инкубации в раствор добавляли 0.06 н HCl, которая снижала pH с 7.3 до 1.85. После обработки пепсином и HCl рН раствора доводили до 7.85 добавлением 2н NaOH. Смесь протеолитических ферментов поджелудочной железы (трипсин/химотрипсин 1:2.3) добавляли к белку с пектином в соотношении фермент/субстрат 1/50. Пробы отбирали через 0.5, 1, 2.5, 4.5, 6.5 и 8.5 часов протеолиза (Mouecoucou et al., 2004). Исследуемые образцы диализовали (пропускная способность пор диализных мешков: 3-5 кДа) для удаления солей. После диализа образцы были лиофильно высушены. Контрольные пробы белка подвергали протеолизу в тех же условиях, но без пектинов. Степень протеолиза оценивали, определяя молекулярную массу расщепленных фрагментов, используя электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия (Laemmli, Fare, 1979).
Регистрация иммунного ответа у мышей на белковый антиген, введенный с пектиновыми полисахаридами. Для изучения развития иммунного ответа на белок у мышей цирсиуман СЕ, лемнан LMC, лемнан LM, цитрусовый и яблочный пектины вводили перорально при помощи полиэтиленового зонда мышам по 1 мг/день в течение 7 дней до и 7 дней после введения белка. Толерантность индуцировали пероральным введением 20 мг OVA, растворенного в 200 мкл PBS. Контрольные животные получали эквивалентный объем PBS. Через 7 дней мышей сенсибилизировали введением 50 мкл OVA в присутствии полного адъюванта Фрейнда (CFA) (OVA/CFA 2 мг/мл) подкожно в основание хвоста (Leishman et al., 2000). Спустя 14 дней после сенсибилизации регистрировали интенсивность иммунного ответа: у животных измеряли реакцию гиперчувствительности замедленного типа (ГЗТ) и концентрацию антиген-специфичных антител. ГЗТ вызывали с помощью инъекции агрегированного нагреванием OVA (25 мкл раствора 1 мг/мл) в левую лапку. В правую лапку вводили 25 мкл PBS. Через 24 часа измеряли толщину лапок с помощью плетизмометра (Ugo Basil, Comerio, Italy) или микрометра (Россия). Интенсивность ГЗТ определяли как разность толщины между левой и правой лапкой (утолщение) и выражали в микролитрах/микрометрах. Концентрацию антиген-специфичных антител определяли с помощью непрямого ИФА (Кэтти, Райкундалиа, 1991).
Оценка противоаллергического действия цирсиумана СЕ. Для исследования развития анафилактической реакции на OVA животным, сенсибилизированным к OVA, трижды перорально с интервалом в 5 дней вводили OVA (1мг) с цирсиуманом СЕ (1 мг) или только белок. Первое введение OVA или белка с пектином бодяка проводили одновременно с сенсибилизацией животных. Контрольные сенсибилизированные мыши перорально получали PBS. Животных сенсибилизировали введением 50 мкл OVA в присутствии СFA (20 мг/мл) подкожно в основание хвоста. Спустя 15 дней после сенсибилизации мышам перорально вводили разрешающую дозу антигена (50 мг) и в течение 30 минут наблюдали развитие анафилактической реакции (Hino et al., 2004). Содержание иммуноглобулинов и цитокинов в крови определяли сразу после последнего перорального введения (через 10 дней после сенсибилизации) и после регистрации анафилаксии (через 15 дней после сенсибилизации). Развитие симптомов анафилактической реакции наблюдали в течение 30 минут после введения разрешающей дозы антигена. Интенсивность анафилаксии определяли по баллам: 0 – нет симптомов; 1 – зуд, почесывание носа и головы; 2 – отечность глаз, рта, снижение активности, повышение частоты дыхания; 3 – свистящее дыхание, цианоз в области носа, хвоста; 4 – продолжительное замедление активности или ее отсутствие после толчка, судороги; 5 – смерть (Li, 2003).
Установление структурных особенностей пектиновой макромолекулы, определяющих стимулирование иммунного ответа на овальбумин. С целью выявить активную область макромолекулы пектинов бодяка и ряски, стимулирующую иммунный ответ на OVA, сотрудники лаборатории гликологии Отдела молекулярной иммунологии и биотехнологии Учреждения РАН Института физиологии Коми НЦ УрО РАН получили фрагменты разветвленной области пектиновой макромолекулы (лемнан LM-P и цирсиуман CE-P). Мышей трижды перорально иммунизировали 1 мг OVA или смесью 1 мг белка и 1 мг пектинов (Kato et al., 2001).. Иммунизацию проводили с интервалом в 7 дней, через неделю после последней иммунизации у животных забирали кровь из сердца и в сыворотке определяли титр антиген-специфичного IgG (Кэтти, Райкундалиа, 1991)
Статистическая обработка результатов. При обработке данных вычисляли среднее арифметическое значение и среднее квадратичное отклонение. Для оценки различий применяли t-критерий Стьюдента для сравнения выборок с нормальным распределением данных, при ненормальном распределении данных вариационного ряда использовали критерий Манна-Уитни, для анализа качественных признаков применяли критерий χ2. Различия признавали значимыми при p<0.05 (Лакин, 1990).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Концентрация овальбумина в крови у мышей после его перорального введения совместно с пектинами. Установлено, что при введении OVA в крови обнаруживается около 1100 нг/мл OVA, что составляет 10-4 от количества введенного белка. Показано, что через три часа после введения содержание OVA в крови мышей, которым вводили белок с лемнанами LM и LMC, цирсиуманом СЕ и цитрусовым пектином PC, в три раза выше, чем у мышей, получавших только белок (рис. 1). Совместное введение OVA с комаруманом СР, бергенаном ВС и гераклеуманом HS приводит к снижению концентрации белка в крови (рис. 1). Проникновение OVA не изменяется в присутствии яблочного пектина РА (рис. 1).

Рис. 1 Концентрация OVA в крови мышей, получавших белок с пектинами: PC, LM, CE, LMC, BC, CP, HS и PA. Контрольные мыши получали только белок.
Примечание: количество животных в группе-8; - p<0,05 по сравнению с OVA.
Изучение динамики проникновения OVA показало, что белок обнаруживается в крови через полчаса, один, три и шесть часов после введения. Содержание белка в сыворотке мышей достигает максимума через шесть часов после введения, в дальнейшем его концентрация постепенно уменьшается (рис. 2). Установлено, что количество белка в сыворотке мышей, получавших OVA с цитрусовым пектином РС, не отличается от контроля через полчаса или час после поступления белка, а в дальнейшем повышается. Цитрусовый пектин не только повышает проникновение OVA в кровь (количество белка у мышей, получавших цитрусовый пектин РС, в три раза выше, чем у животных, получавших только OVA), но и увеличивает длительность нахождения белка в крови. Показано, что OVA обнаруживается спустя сутки после введения в крови мышей, получавших OVA с цитрусовым пектином РС, тогда как в сыворотке мышей, получавших только белок, через сутки OVA отсутствует (рис. 2).

Рис. 2. Динамика проникновения в кровь OVA, введенного перорально с цитрусовым пектином (
 ). Контрольные мыши получали только белок (
). Контрольные мыши получали только белок ( ). Примечание: количество животных в группе-7; - p<0.05 по сравнению с OVA.
). Примечание: количество животных в группе-7; - p<0.05 по сравнению с OVA. Полученные нами данные согласуются с результатами, полученными ранее, в которых установлено, что количество OVA, проникшего в кровь после перорального введения, составляет 10-5 – 10-4 от общего количества введенного белка (Peng et al., 1990; Fujihashi et al., 2001). Пенг (Peng) и соавторы, определяя содержание иммуногенного OVA, проникшего в кровь, выявили, что иммуногенный белок в крови обнаруживается уже через пять минут после введения, а через час его содержание составляет 10-5 от общего количества введенного белка (Peng et al., 1990). Японские исследователи показали, что OVA, перорально введенный мышам, проникает в кровь уже через полчаса после введения, в дальнейшем его концентрация в крови постепенно возрастает и достигает 10-4 от общего количества введенного белка (Fujihashi et al., 2001). Вероятнее всего, в основном транспорт белка происходит через энтероциты тонкой кишки, в которую OVA попадает через три часа после введения. Отсутствие OVA в крови через сутки после введения объясняется, вероятнее всего, тем, что весь белок к этому времени переваривается и удаляется из кишечника. Проницаемость кишечной стенки для OVA изменяется, если белок поступает с пектиновыми полисахаридами. Модулирование проникновения белков пектинами ранее не было показано. Известно лишь, что пектины снижают всасывание микронутриентов и глюкозы благодаря их способности образовывать комплексы с другими молекулами (Fuse et al., 1989). Установлено, что пектины по-разному влияют на проникновение OVA. Вероятнее всего, различия в структуре исследованных пектинов определяют их действие на проницаемость кишечной стенки для OVA, вводимого с этими пектинами.
Содержание цитокинов в слизистой оболочке тонкой кишки и продукция цитокинов макрофагами у мышей, получавших пектины перорально. Показано, что концентрация цитокинов в слизистой тонкой кишки мышей, получавших перорально OVA, не отличается от уровня цитокинов, обнаруженного в слизистой животных, которым вводили PBS (контроль). Установлено, что содержание ИФН-γ, ИЛ-1β, ФНО-α в слизистой кишки мышей, получавших OVA с цитрусовым пектином, лемнаном LM и цирсиуманом СЕ, выше, чем у мышей, которым вводили только белок. Совместное пероральное введение OVA с бергенаном ВС и комаруманом СР приводит к снижению уровня цитокинов в слизистой (рис. 3).
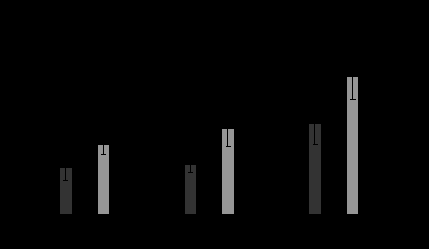
Рис. 3. Содержание ИФН-, ИЛ-1, ФНО- в слизистой тонкой кишки мышей через три часа после введения OVA или OVA с пектинами.
Примечание: количество животных в группе-8; - p<0.05 по сравнению с OVA.
Изменение проницаемости кишечной стенки для OVA у мышей, получавших белок с пектинами, может быть обусловлено изменением содержания цитокинов, повышающих проницаемость кишечной стенки. Известно, что увеличение концентрации ФНО-, ИФН-, ИЛ-1 в среде приводит к повышению как парацеллюлярного, так и трансцеллюлярного транспорта белков (Stein et al., 1998; Ma et al., 2005; Berin et al.,1999).
Изучение секреции цитокинов иммунными клетками у мышей, получавших пектины, показало, что продукция ФНО- и ИФН- перитонеальными макрофагами выше у мышей, получавших овальбумин с цитрусовым пектином (табл. 1).
Таблица 1
Продукция цитокинов перитонеальными макрофагами у мышей, получавших OVA с цитрусовым пектином
-
п/о введение
n
концентрация цитокинов, нг/мл
ФНО-
ИФН-
OVA
7
59±3
138±10
OVA+PC
7
84±8*
184±15*
Примечание: - p<0.05 по сравнению с OVA; n - количество животных в группе.
Способность пектинов, поступающих перорально, стимулировать секрецию цитокинов макрофагами ранее не была показана. Опубликованы работы, свидетельствующие о способности растительных полисахаридов активировать продукцию провоспалительных цитокинов макрофагами (Inngjerdingen et al., 2007б; Leung et al., 2004). Однако в данных работах полисахариды вводили в перитонеальную полость (Leung et al., 2004) или использовали макрофагальную культуру клеток (Inngjerdingen et al., 2007б). Это обеспечивает непосредственный контакт макрофага с полисахаридом. В нашем же исследовании пектин поступает перорально, активация макрофагов перитонеальной полости, по-видимому, является опосредованной.
При определении степени протеолиза белка, гидролизованного в присутствии цитрусового пектина РС и гераклеумана HS, установлено, что OVA подвергается протеолизу быстрее в присутствии цитрусового пектина. На это указывает наличие фрагмента с молекулярной массой 14.4 кДа через 1 час протеолиза белка. В контрольных пробах подобный фрагмент появляется только через 4.5 часа гидролиза. Более того, даже через 8.5 часов протеолиза в контрольных пробах сохраняется высокомолекулярный фрагмент, а в образцах, гидролизованных с цитрусовым пектином, он отсутствует (рис. 4Б). Показано, что в пробах белка, подвергнутого протеолизу в присутствии гераклеумана HS, фрагмент с молекулярной массой 14.4 кДа через 4.5 часа протеолиза отсутствует, что указывает на ингибирование протеолиза белка пектином борщевика (рис. 4В).
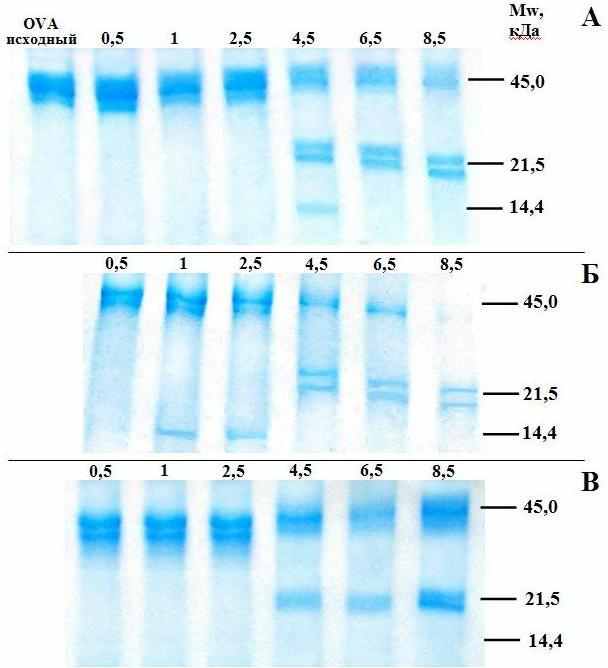
Рис. 4. Электрофореграмма фрагментов OVA, полученных в результате протеолиза белка in vitro в отсутствии (А) и в присутствии цитрусового пектина (Б) или гераклеумана HS (В).
Расщепление белковой макромолекулы в ходе протеолиза приводит к образованию фрагментов различной молекулярной массы. Очевидно, что фрагмент OVA, имеющий молекулярную массу 14 кДа, легче проникает через кишечный барьер, чем нативный OVA, молекулярная масса которого в три раза выше. По-видимому, усиление протеолиза OVA в присутствии цитрусового пектина способствует его проникновению в кровь. В результате протеолиза OVA с гераклеуманом HS, напротив, такого фрагмента с молекулярной массой 14 кДа не образуется, поэтому OVA меньше проникает в кровь.
Характеристика иммунного ответа на овальбумин, введенный перорально с пектиновыми полисахаридами. Установлено, что подкожная иммунизация мышей OVA в присутствии полного адъюванта Фрейнда приводит к развитию иммунного ответа против OVA. В ответ на подкожную инъекцию агрегированного нагреванием OVA в лапку сенсибилизированным мышам (контроль) развивается реакция ГЗТ, что говорит о вовлечении Т-клеточного звена в иммунный ответ (рис. 5А). Кроме того, в сыворотке сенсибилизированных мышей обнаруживается высокий титр антиген-специфичных антител класса IgG (рис. 5Б). Это свидетельствует о выраженном гуморальном ответе на введенный белок. Введение белка (20 мг) мышам за 7 дней до сенсибилизации вызывает толерантность – существенное снижение иммунного ответа. У толерантных мышей (группа «OVA») снижена реакция ГЗТ и продукция IgG (рис. 5А, Б). У животных, получавших лемнаны LM и LMC, цирсиуман CE и цитрусовый пектин с OVA, интенсивность ГЗТ (рис. 5А) и концентрация антител в сыворотке (рис. 5Б) выше, чем у толерантных мышей. Таким образом, пектиновые полисахариды ряски LM и LMC, бодяка CE и цитрусовый пектин повышают иммунный ответ на белковый антиген. Яблочный пектин не влияет на развитие толерантности к OVA (рис. 5А, Б).
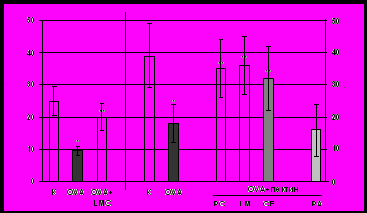
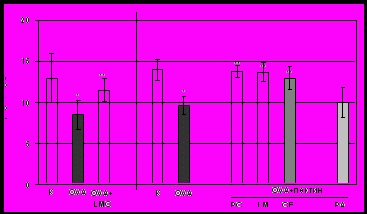
Рис. 5 Развитие оральной толерантности к OVA: ГЗТ (А) и продукция анти-OVA-IgG (Б). Примечание: количество животных в группе -7; *- p<0.05 по сравнению контрольными мышами; **- p<0.05 по сравнению с OVA.
Определение подклассов иммунноглобулина IgG (IgG1, IgG2a) и IgЕ показало, что контрольные сенсибилизированные мыши имеют высокие титры антител. В сыворотке толерантных мышей концентрация иммуноглобулинов существенно ниже, чем в контроле. Концентрация анти-OVA-IgG1 у мышей, получавших цитрусовый пектин, в два раза выше по сравнению с толерантными животными, тогда как концентрация IgG2а существенно не меняется. Введение цирсиумана CE приводит к стимуляции продукции IgG2а. В сыворотке мышей, получавших лемнаны LM и LMC, содержание обоих подклассов IgG выше, чем у толерантных животных. Концентрация IgЕ выше у мышей, получавших цитрусовый пектин и пектиновый полисахарид ряски LMC по сравнению с толерантными. Цирсиуман СЕ и лемнан LM не стимулируют продукцию IgЕ. Яблочный пектин не влияет на продукцию подклассов иммунноглобулина IgG и IgЕ (табл. 2).
Таблица 2
Концентрация анти-OVA-IgG1, IgG2a и IgЕ в сыворотке мышей, получавших OVA или OVA с пектинами
| п/о введение | n | титр IgG1, log2 | титр IgG2a, log2 | IgE, мкг/мл | |
| анти-OVA | общий | ||||
| PBS | 7 | 13.0±2.1 | 10.8±2.0 | 6.6±1.0 | 930±159 |
| OVA | 7 | 7.3±2.6* | 4.8±1.5* | 4.4±0.5* | 590±48* |
| OVA+LMC | 7 | 11.5±1.4** | 8.6±1.7** | 6.1±1.0** | 925±127** |
| OVA+LM | 7 | 10±1.2** | 7.8±1.5** | 5.0±1.1 | 520±88 |
| OVA+СЕ | 7 | 9.5±1.8 | 13.0±0.7** | 3.2±1.3** | 480±32** |
| OVA+PC | 7 | 13.4±1.7** | 7.1±2.7 | 6.1±0.8** | 780±59** |
| OVA+AP | 7 | 5.6±2.4 | 5.0±2.4 | 7.8±1.5 | 630±150 |
Примечание: - p<0.05 по сравнению с контролем; **- p<0.05 по сравнению толерантными мышами; n - количество животных в группе.
Повышение ГЗТ и стимуляция продукции IgG2a указывает на активацию Т-хелпер 1 (Th-1) лимфоцитов и других иммунокомпетентных клеток (макрофагов, нейтрофилов), участвующих в генерировании ГЗТ. Активация Th-1 иммунного ответа пектиновыми веществами была показана в ряде работ (Lee, 2004; Vos, 2004; Vos, 2007). В свою очередь, Т-хелпер 2 (Th-2) лимфоциты, выделяют ряд цитокинов, заставляющих В-лимфоцит секретировать IgG1, IgЕ (Marinaro et al., 1995). Поэтому стимуляция продукции IgG1, наряду с IgG2a у мышей, получавших лемнаны LM и LMC, свидетельствует об активации также и Th-2 лимфоцитов пектинами. Смешанный (Th-1 и Th-2) тип активации иммунитета характерен и для известных иммуноадъювантов (Su et al., 2004; Richards et al., 2001; Boyaka et al., 2001). Цитрусовый пектин активирует продукцию IgG1, IgE, что свидетельствует о стимуляции Th-2 лимфоцитов. Пероральное введение цирсиумана СЕ не приводит к увеличению IgG1 и IgE антител, а, напротив, стимулирует только образование анти-ОVА-IgG2а, что, по-видимому, указывает на способность пектина бодяка активировать преимущественно Th-1 тип иммунного ответа.
Действие пектина бодяка, цирсиумана СЕ, на развитие аллергической реакции. Установлено, что у сенсибилизированных мышей в ответ на введение разрешающей дозы аллергена развивается сильная анафилактическая реакция (100% контрольных животных имеют высокие баллы: 3 – 4). Подобная интенсивность анафилаксии наблюдается у 75% сенсибилизированных мышей, перорально получивших OVA, тогда как степень анафилаксии в 2 раза ниже (38%), если животные получают OVA с цирсиуманом СЕ.
Показано, что содержание анти-ОVА-IgE в сыворотке контрольных сенсибилизированных животных составляет 40 нг/мл спустя 10 дней после сенсибилизации и увеличивается после введения разрешающей дозы белка. Пероральная иммунизация OVA сенсибилизированных мышей стимулирует продукцию антител: концентрация анти-ОVА-IgE выше контрольных значений через 10 дней после сенсибилизации, а после введения разрешающей дозы белка (через 15 дней) уровень IgE сравнивается с контрольными значениями. Введение OVA с цирсиуманом СЕ приводит к уменьшению концентрации анти-ОVА-IgE как через 10, так и через 15 дней. Аналогичные результаты получены при определении содержания общего IgE в сыворотках животных (табл. 3).
Таблица 3
Содержание IgE в сыворотке крови у мышей, перорально иммунизированных OVA или ОVА с цирсиуманом СЕ
| п/о введение | n | 10 дней | 15 дней | ||
| анти-ОVА, нг/мл | общий, мкг/мл | анти-ОVА, нг/мл | общий, мкг/мл | ||
| PBS | 8 | 40±4 | 986±108 | 60±10 | 1318±190 |
| ОVА | 8 | 68±4* | 1678±306* | 50±16 | 1152±185 |
| ОVА+СЕ | 8 | 43±7** | 1090±128** | 28±8** | 756±65** |
Примечание: - p<0.05 по сравнению с контролем; **- p<0.05 по сравнению с мышами, иммунизированными OVA; n - количество животных в группе.
Показано, что цирсиуман ингибирует анафилаксию, снижая содержание IgE и IgG1, и, напротив, повышает продукцию ИФН- и IgG2a. Подобный путь противоаллергического действия веществ уже был показан в ряде работ (Hasegawa et al., 1999; Shida et al., 2002; Saito et al., 2003; Marinaro et al., 1999). Судя по всему, цирсиуман может снижать аллергическую реакцию на белок на стадии сенсибилизации, ингибируя образование аллерген-специфичного иммуноглобулина IgE. Наиболее вероятной мишенью воздействия пектина бодяка является антиген-презентирующие клетки кишечника, секретирующие ИЛ-12 (макрофаги, дендритные клетки). В свою очередь, ИЛ-12 стимулирует продукцию ИФН-γ Th-1 лимфоцитами, что приводит к ингибированию секреции IgЕ. Наши данные подтверждают ранее полученные результаты ингибирования аллергии веществами, сдвигающими иммунный ответ с Th-2 пути на Th-1 путь иммунного ответа.
Иммунный ответ на OVA у мышей, иммунизированных OVA или смесью OVA с лемнаном LM-P и цирсиуманом СЕ-Р. Показано, что у животных, перорально иммунизированных только OVA (контроль), содержание антиген-специфичных антител в сыворотке незначительно. Трехкратная пероральная иммунизация OVA как с исходными пектинами бодяка и ряски (цирсиуман СЕ и лемнан LM), так и с лемнаном LM-P и цирсиуманом СЕ-Р, полученными в результате ферментативного гидролиза, приводит к повышению титра анти-OVA-IgG в сыворотке (табл. 4).
Таблица 4
Концентрация антиген-специфичных антител в крови мышей, иммунизированных только OVA или OVA с пектинами
-
п/о введение
n
титр анти-ОVА-IgG, log2-
OVA (контроль)
8
7.3±1,3
OVA+LM
8
11.3±0.5*
OVA+LM-P
8
12.0±1.7*
OVA+CE
8
13.3±1.2*
OVA+CE-P
8
14.8±1.7*
Примечание: - p<0.05 по сравнению с OVA; n - количество животных в группе
Стимуляция иммунного ответа лемнаном LM-P и цирсиуманом CE-P свидетельствует о том, что именно разветвленный участок пектиновой молекулы определяет иммуностимулирующее действие пектинов. Результаты, согласующиеся с нашими, были получены ранее японскими исследователями (Matsumoto, Yamada, 1995; Inngjerdingen et al., 2007a). Более того, установлено, что повышение содержания остатков галактуроновой кислоты в макромолекуле пектина приводит к снижению его иммуностимулирующей активности (Kiyohara et al., 2002).
ВЫВОДЫ
- Овальбумин, введенный перорально, частично проникает в кровь, сохраняя свои антигенные свойства. Проницаемость кишечной стенки для овальбумина увеличивается при его совместном введении с цитрусовым пектином (РС), пектинами ряски (лемнанами LM, LMC) и бодяка (цирсиуманом СЕ).
- Увеличение проницаемости кишечной стенки обусловлено повышением концентрации цитокинов (ФНО-, ИЛ-1, ИФН-) в слизистой оболочке кишки, стимуляцией продукции цитокинов макрофагами, а также усилением протеолиза белка под действием пектинов.
- Повышение концентрации иммуногенного овальбумина в крови, вызванное пектинами, приводит к активации у мышей иммунного ответа на белок: происходит усиление реакции ГЗТ и стимулирование образования антител.
- Пектин бодяка, цирсиуман СЕ, препятствует развитию аллергической реакции, что обусловлено уменьшением продукции IgE.
- Активной областью лемнана LM и цирсиумана СЕ является разветвленный фрагмент пектиновой макромолекулы.
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
Статьи:
- Храмова Д.С., Гюнтер Е.А. Действие лемнана, пектина из каллуса ряски малой Lemna minor L. на толерантность к белковому антигену // Вестник уральской медицинской академической науки. Екатеринбург. 2006. №3-1(14). С.324-326.
- Popov S.V., Günter E.A., Markov P.A., Smirnov V.V., Khramova D.S., Ovodov Yu.S. Adjuvant effect of lemnan, pectic polysaccharide of callus culture of Lemna minor L. at oral administration // Immunopharmacol. Immunotoxicol. 2006. Vol.28. P.141-152.
- Popov S.V., Golovchenko V.V., Ovodova R.G., Smirnov V.V., Khramova D.S., Popova G.Yu., Ovodov Yu.S. Characterisation of the oral adjuvant effect of lemnan, a pectic polysaccharide of Lemna minor L. // Vaccine. 2006. Vol.24. P.5413-5419.
- Khramova D.S., Popov S.V., Golovchenko V.V., Vityazev F.V., Paderin N.M., Ovodov Yu.S. Abrogation of the oral tolerance to ovalbumin in mice by citrus pectin // Nutrition. 2009. Vol. 25. Р. 226-232.
Тезисы докладов на конференции:
- Храмова Д.С. Ингибирующее действие цитрусового пектина на толерантность к белковому антигену // Тезисы докладов VI Молодежной научной конференции Института физиологии Коми НЦ УрО РАН, «Физиология человека и животных: от эксперимента к клинической практике». Сыктывкар, 2007. С. 128-130.
- Храмова Д.С., Гюнтер Е.А Выделение и иммуностимулирующая активность лемнана, пектина из каллуса ряски малой Lemna minor L. // Материалы докладов XIII Молодежной научной конференции Института биологии Коми НЦ УрО РАН «Актуальные проблемы биологии и экологии». Сыктывкар, 2007. С.294-267.
- Khramova D.S., Günter E.A. Effects of citrus pectin and lemnan, pectin from callus culture of duckweed Lemna minor (L.) on oral tolerance and intestinal permeability // Abstract of the 14th European carbohydrate symposium “Eurocarb 14 ”, Lübeck, 2-8 September 2007. Lubeck, 2007. P.383.
- Храмова Д.С., Головченко В.В. Выделение и иммуностимулирующая активность цирсиумана, пектина из стеблей бодяка съедобного Cirsium esculentum (Siev.), при пероральном введении // Тезисы докладов V Всероссийской школы-конференции «Химия и технология растительных веществ». Уфа, 2008. С.62
- Khramova D.S., Golovchenko V.V., Otgonbayar D., Chimidsogzol A. Isolation, characterization, immunostimulatory and anti-allergic activities of cirsiuman, pectin from ground thistle Cirsium esculentum Siev. // Abstract of the III International conference «Сhemical investigation and utilization of natural resources». Ulan-Bator, 2008. P. 62
Считаю своим приятным долгом выразить искреннюю благодарность своему научному руководителю кандидату биологических наук, доценту Сергею Владимировичу Попову и научному консультанту академику Юрию Семеновичу Оводову за ценные советы и консультации, кандидату химических наук, доценту Виктории Владимировне Головченко, кандидату химических наук, ст.н.с. Раисе Григорьевне Оводовой и кандидату биологических наук, доценту Елене Александровне Гюнтер за предоставленные образцы пектиновых полисахаридов и их структурно-химические характеристики, а также всем сотрудникам Отдела молекулярной иммунологии и биотехнологии Учреждения РАН Института физиологии Коми НЦ УрО РАН за поддержку и помощь в подготовке и выполнении работы.
Настоящая работа выполнена в соответствии с планами НИР Учреждения РАН Института физиологии Коми НЦ УрО РАН на 2005-2008 гг. (ГР № 01.200107401) и поддержана грантами РФФИ (06-04-48079-а; 07-04-90120_монг_а), Программы ведущих научных школ (№ НШ-5796.2006.4), Программы Президиума РАН, «Молекулярная и клеточная биология», Программы президиума РАН, «Фундаментальные науки – медицине», Фонда содействия отечественной науке по программе "Лучшие аспиранты РАН", научных проектов молодых ученых и аспирантов УрО РАН.

2
19

3
18
4
17
5
16
6
15
7
14
А
Б
8
13
9
12
10
11