
Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году в сборниках по проблемам кибернетики. Тогда же вышла статья
| Вид материала | Статья |
СодержаниеОнтогенетическое правило полового диморфизма Филогенетическое правило полового диморфизма Динамика полового диморфизма в онтогенезе |
- Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году, 4857.78kb.
- I вступление, 32.31kb.
- Культура Кольского Севера; Творческие работы посвященные проблемам студенческой молодежи, 67.08kb.
- «Мой учитель» посвященные к Году учителя, 64.26kb.
- Михаил Александрович Шолохов (родился. В 1905 году) Когда в 1925 году появились Донские, 334.1kb.
- Лекция 21. Генетика пола Определение пола, 33.63kb.
- Публичный доклад о результатах деятельности школы моу сош п. Пола в 2008-2009 учебном, 335.49kb.
- Курс «Обзор перспективных технологий Microsoft. Net» Губанов Ю. А., математико-механический, 177.56kb.
- Больше-Кочинский школьный музей этнографии и фольклора, 25.15kb.
- Воспитание и обучение одаренных детей, 267.86kb.
Онтогенетическое правило полового диморфизмаРассмотрим три явления—филогенез, онтогенез и половой диморфизм. Уже давно ученые заметили существование связи между явлениями филогенеза и онтогенеза. Согласно биогенетическому закону Геккеля-Мюллера (закон рекапитуляции), каждый организм в своем индивидуальном развитии повторяет взрослые формы своих предков. По мнению К. Бэра (1828 г.) “Эмбрионы последовательно переходят в своем развитии от общих признаков типа ко все более специальным признакам. Позднее всего развиваются признаки, указывающие на принадлежность эмбриона к определенному роду, виду, и, наконец, развитие завершается появлением характерных особенностей данной особи” (закон зародышевого сходства). Хотя точка зрения Э. Геккеля не признается современной биологической наукой, связь между филогенезом и онтогенезом невозможно отрицать (Gould, 1977). Филогенетическое правило полового диморфизма связывает явления филогенеза и полового диморфизма. Таким образом, мы имеем три явления—филогенез, онтогенез и половой диморфизм,—которые связаны между собой двумя закономерностями. (законом Геккеля-Мюллера и филогенетическим правилом полового диморфизма). Можно предсказать, существование третьей закономерности—непосредственной связи между явлениями полового диморфизма и онтогенеза (Рис. 10.2). Р 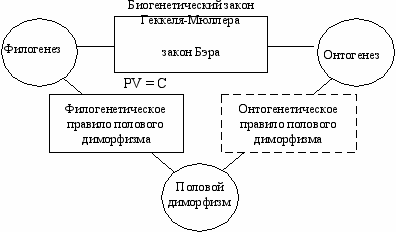 ис. 10.2 ис. 10.2Закономерности, связывающие явления филогенеза, онтогенеза и полового диморфизма (Геодакян, 1983а). Новую закономерность можно назвать “Онтогенетическим правилом полового диморфизма”. Если по какому-либо признаку cyществует генотипический популяционный половой диморфизм, то в онтогенезе (с возрастом) этот признак меняется, как правило, от женской формы к мужской, то есть для начальной, ювенильной стадии онтогенеза (стадии детства, роста и становления) более характерна женская форма признака, а для дефинитивной (зрелой, взрослой)—мужская. Иными словами, женские формы признаков с возрастом должны, как правило, ослабевать, а мужские—усиливаться. онтогенетическое правило полового диморфизма если по какому-либо признаку существует популяционный половой диморфизм, то в онтогенезе этот признак меняется, как правило, от женской формы к мужской Динамика полового диморфизма в онтогенезеКаждую из филогенетических фаз, рассмотренных в Гл. 9 (Рис. 9.5), можно спроецировать на онтогенез и получить соответствующие им 6 типов динамики полового диморфизма в индивидуальном развитии (Рис. 10.3) (Геодакян, 1994). Дихронизм проявится в онтогенезе как возрастное запаздывание в развитии признака у женского пола, то есть доминирование женской формы диморфного признака в начале онтогенеза и мужской в конце. Р 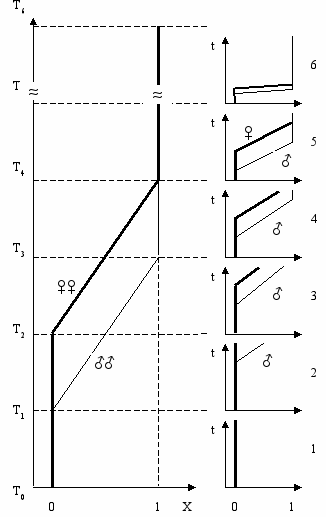 ис. 10.3 Фазы эволюции признака (0→1) в филогенезе и соответствующие им 6 последовательных типов динамики полового диморфизма в онтогенезе (Геодакян, 1991). По осям: Х - среднепопуляционный генотип. Т - время в филогенетическом масштабе. t - время в онтогенетическом масштабе 1 — Онтогенез доэволюционной фазы. Признак одинаков у мужского и женского пола и постоянен. Половой диморфизм отсутствует. 2 — Онтогенез дивергентной фазы. Признак постояннен у женского пола, у мужского пола он меняется в конце. Половой диморфизм появляется в позднем возрасте и только растет. 3 — Онтогенез параллельной фазы. Признак меняется и у мужского пола и позже у женского. Половой диморфизм возникает в более раннем возрасте (чем в 2), сначала растет, потом постоянен. 4 — Онтогенез конвергентной фазы. Половой диморфизм возникает в более раннем возрасте (чем в 3). В отличие от 3 половой диморфизм после возраста роста и постоянства в конце уменьшается, но не исчезает. 5 — Онтогенез послеэволюционной фазы. Вся картина сдвинута к началу онтогенеза и в конце онтогенеза половой диморфизм исчезает. 6 — Онтогенез межэволюционной фазы. "Петля" полового дихронизма спрессована в эмбриогенез. А в постнатальном онтогенезе половой диморфизм отсутствует. Такая трактовка возрастной динамики полового диморфизма подтверждается, например, хорошо известной картиной появления рогов у самцов и самок оленей и антилоп в онтогенезе и филогенезе. Можно проследить четкую связь между степенью рогатости вида, с одной стороны, и возрастом появления рогов у самцов и самок—с другой. Чем сильнее выражены рога у вида, тем в более раннем возрасте они появляются сначала у самцов, а затем у самок. Это возрастное запаздывание развития признака у самок есть не что иное как онтогенетическая проекция предсказанного теорией филогенетического полового дихронизма. На более тесную связь женского пола с начальной фазой онтогенеза обратил внимание ещё Дарвин. Он писал: “Во всем животном царстве, если мужcкoй и женский пол отличаются друг от друга по внешнему виду, видоизменяется, за редкими исключениями, самец, а не самка, потому что последняя обыкновенно остается схожей с молодыми животными своего вида и с прочими членами всей группы” (Дарвин, 1953). Антропологи также отмечали близость женского типа с детским (более грацильные кости, слабовыраженные надбровные дуги, меньшее обволошение тела и др.) (Гинзбург, 1963; Рогинский, Левин, 1955). Для проверки новой закономерности необходимо сопоставить направление полового диморфизма по разным признакам с динамикой этих признаков в онтогенезе. Например, существует половой диморфизм по горбинке носа: она чаще встречается и сильнее бывает выражена у мужского пола (Рогинский, Левин, 1955). Значит, женская форма признака—вогнутая спинка носа, мужская—выпуклая. В онтогенезе спинка носа должна меняться от вогнутой к выпуклой. Как известно, это так и есть. В Табл. 10.1 (Приложение В) приведены данные для некоторых антропологических признаков (относительная длина ссылка скрыта, предплечья, 4-го и 2-го пальцев, головной индекс, окружность зубной дуги, ссылка скрыта, горбинка спинки носа, обволошение тела, лица и головы, концентрация ссылка скрыта в ссылка скрыта, ссылка скрыта, скорость опорожнения желчного пузыря, асимметрия мозга, время реакции, ощущение горького вкуса фенилтиомочевины и обоняние). Видно, что предсказанная теорией закономерность действительно существует. Глава 11 Родительские эффекты В ряде случаев фенотип потомства зависит от генетического вклада отца или матери (так называемые родительские эффекты (“parent-of-origin effect”)). Допустим, что существуют два варианта организма—А и B. Классическая генетика предполагает, что фенотип гибрида АB будет одним и тем же независимо от того скрещивается ли самка типа А с самцом типа B или наоборот. Напротив, когда ♀А x ♂B ≠ ♀B x ♂А, мы имеем дело с родительскими эффектами (Gray, 1972). Так например, скрещивание самки тигра (Panthera tigris) с самцом льва (Panthera leo) дает гибрид под названием лигр. Лигры больше каждого из родителей и часто достигают 3 м в длину и весят больше 350 кг (Morison et. al., 2001). Потомство, получаемое от обратного скрещивания (самки льва с самцом тигра) обычно меньше каждого из родителей. |