
Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году в сборниках по проблемам кибернетики. Тогда же вышла статья
| Вид материала | Статья |
- Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году, 4857.78kb.
- I вступление, 32.31kb.
- Культура Кольского Севера; Творческие работы посвященные проблемам студенческой молодежи, 67.08kb.
- «Мой учитель» посвященные к Году учителя, 64.26kb.
- Михаил Александрович Шолохов (родился. В 1905 году) Когда в 1925 году появились Донские, 334.1kb.
- Лекция 21. Генетика пола Определение пола, 33.63kb.
- Публичный доклад о результатах деятельности школы моу сош п. Пола в 2008-2009 учебном, 335.49kb.
- Курс «Обзор перспективных технологий Microsoft. Net» Губанов Ю. А., математико-механический, 177.56kb.
- Больше-Кочинский школьный музей этнографии и фольклора, 25.15kb.
- Воспитание и обучение одаренных детей, 267.86kb.
Половой диморфизм и репродуктивная структура популяции
Половой диморфизм должен быть связан с репродуктивной структурой популяции: у строгих моногамов он минимален, поскольку моногамы используют специализацию полов только на уровне организма, но не популяции, а у панмиктных видов и полигамов, полнее использующих преимущества дифференциции, он возрастает с ростом степени полигамии.
Реверсия полового диморфизма при полиандрии
Полиандрия, при которой самка спаривается с несколькими самцами, то есть имеет более высокий репродуктивный индекс (число реализованных гамет) чем самец, встречается у беспозвоночных, рыб, птиц, млекопитающих. При этом часто наблюдается реверсия полового диморфизма (самки крупнее самцов и ярче окрашены, самцы строят гнездо, насиживают яйца и заботятся о выводке, отсутствует борьба за самку). При полигинии—картина обратная. Это значит, что направление полового диморфизма, а стало быть и асинхронии и соотношение скоростей эволюции полов зависят от направления полигамии, или соотношения репродуктивных индексов полов. У моногамов число отцов и матерей одинаково, то есть индексы самцов и самок равны. Тогда одинаковы также и дисперсии Y-хромосом у сыновей и материнских Х-хромосом у дочерей. В случае полигинии, когда отцов меньше, чем матерей, эта дисперсия у сыновей меньше, чем у дочерей. В случае полиандрии—все наоборот, так как дисперсия Y-хромосомы у сыновей пропорциональна числу отцов, тогда как дисперсия материнских Х-хромосом у дочерей пропорциональна числу матерей.
С другой стороны, для того, чтобы половой диморфизм по норме реакции предшествовал эволюции любого признака необходимо, чтобы ширина нормы реакции определялась половыми гормонами (ведь согласно теории половые гормоны—вещества, регулирующие “расстояние” системы от среды: андрогены—приближающие, эстрогены—удаляющие). Кроме того, известно, что Y-хромосома запускает синтез тестостерона, концентрация которого определяет половой диморфизм.
Все это позволяет высказать гипотезу о том, что норма реакции определяется половыми гормонами, а ее величина обратно пропорциональна концентрации тестостерона в организме. Тогда, объединив эти индуктивные и дедуктивные выводы, можно построить причинно-следственную зависимость и связать направление эволюционного полового диморфизма, то есть соотношение скоростей эволюции мужского и женского пола ( Эм/Эж ) c соотношением их репродуктивных индексов:
( Рм/Рж ). Рм/Рж ~ Nматерей/Nотцов ~ σX /σY ~ Тм/Tж ~ Нж/Нм ~ sм/s ж ~ Эм/Э ж ~ ЭПД [ 4 ]
где: N—численность, σY , и σX—дисперсии Y хромосомы у сыновей и материнской Х-хромосомы у дочерей, Тм , Тж—концентрации тестостерона, Нм , Нж—нормы реакций, sм , sж—коэффициенты отбора, ~—знак пропорциональности.
Следовательно, роль эволюционного “авангарда” всегда получает более полигамный пол, а “арьергарда”—моногамный. И чем выше индекс полигамии, тем больше может быть эволюционный половой диморфизм (у строгих моногамов ЭПД = 0, поэтому минимальный половой диморфизм).
В панмиктной популяции, могут быть все три формы брачных отношений: моногамия—рождаются полные сибсы ( ПС ), полигиния—отцовские полусибсы ( ОпС ) и полиандрия—материнские полусибсы ( МпС ). Их концентрации в популяции связаны между собой подвижным равновесием: [ ОпС ] ↔ [ ПС ] ↔ [ МпС ] и регулируются условиями среды: в оптимальных оно сдвигается вправо, повышая эволюционную стабильность популяции, а в экстремальных—влево, повышая её эволюционную пластичность.
Полигиния широко распространена в природе, а полиандрия встречается редко. Это связано с потенциально бóльшими репродуктивными возможностями мужского пола (в конечном счете бóльшим числом гамет). Практически, полиандрии, как таковой, не существует. Строго говоря это только олигоандрия, так как репродуктивные возможности женского пола ограничены.
Глава 10
Половой диморфизм—правила
Многие теории рассматривают половой диморфизм в тесной связи с полом. Например, Дарвин считал его следствием полового отбора. Поэтому эти теории не могли объяснить существование полового диморфизма по признакам не связанным с половым отбором, половой диморфизм у растений или половой диморфизм по врожденным порокам развития.
Эволюционная теория пола связывает половой диморфизм с эволюцией признаков. Поэтому он возникает в результате любого отбора как следствие асинхронной эволюции полов. В этой главе рассмотрены различные формы полового диморфизма и правила, которые можно сформулировать на основе нового подхода.
Филогенетическое правило полового диморфизма
Учитывая известную необратимость эволюции (закон Долло), можно рассматривать мужской пол как эволюционный “авангард” популяции, а половой диморфизм по признаку как “дистанцию” между полами, и следовательно как вектор, показывающий направление эволюции этого признака. Вектор этот направлен от популяционной нормы женского пола по данному признаку к норме мужского. Стало быть, признаки, чаще появляющиеся или/и четче выраженные у женского пола должны иметь “атавистическую” природу, а чаще появляющиеся у мужского пола—“футуристическую” (поиск). Иными словами, мужской пол осуществляет эволюционный поиск и пробы, поэтому включает и находки (новаторство, прогрессивность) и ошибки (несовершенство), а женский пол осуществляет отбор и закрепление уже опробированного (совершенство).
Если по признакам, встречающимся у обоих полов, cyщecтвует популяционный половой диморфизм, его можно связать с эволюционной тенденцией изменения этого признака и сформулировать “Филогенетическое правило полового диморфизма”: если по какому-либо признаку имеется генотипический популяционный половой диморфизм, то есть имеются две формы этого признака женская и мужская (по степени выраженности (экспрессивности) или по частоте встречаемости (пенетрантности)), то этот признак эволюционирует от женской формы к мужской.
Филогенетическое правило полового диморфизма
если по какому-либо признаку существует генотипический популяционный половой диморфизм, то этот признак эволюционирует от женской формы к мужской
Генотипический популяционный половой диморфизм может служить “компасом”, показывающим направление эволюции признака. При этом если дисперсия признака больше у мужcкoгo пола—фаза дивергентная, дисперсии равны—параллельная, дисперсия больше у женского пола—фаза конвергентная (“Филогенетическое правило дисперсии полов”).
Филогенетическое правило ДИСПЕРСИИ полов
если ДИСПЕРСИЯ признакА больше у мужcкoгo пола—фаза эволюции признака дивергентная, дисперсии равны—параллельная, дисперсия больше у женского пола—фаза конвергентная
Филогенетическое правило полового диморфизма было применено для исследования эволюции низших ракообразных семейства Chidoridae (173 вида) (Геодакян, Смирнов, 1968). У ветвистоусых ракообразных самцы во всех без исключения случаях меньше самок. Следовательно, на основании правила полового диморфизма можно предположить, что они на данном этапе эволюции проявляют общую тенденцию к уменьшению размеров. Действительно, хорошо известно, что морфологически более примитивные формы низших ракообразных отличаются крупными размерами. В пределах группы истинно листоногих и ветвистоусых ракообразных прослеживаются ряды форм с последовательным уменьшением размера, числа конечностей и специализацией в соответствии с филогенетической преемственностью и меньшим размером самцов.
По ряду признаков самцы из морфологически более примитивных секций предваряют те черты, которые появляются в последующих секциях (морфологически более развитых). Можно сделать вывод, что эволюционная тенденция признаков, определяемая, с одной стороны, по морфологическим критериям, а с другой стороны, на основании правила полового диморфизма, совпадает. Такой вывод дает основание применять новое правило для дальнейших исследований. Можно высказать некоторые предположения о биологическом смысле происходящих изменений. При уменьшении размера возрастает относительная поверхность, что способствует газообмену и увеличивает плавучесть. С. С. Четвериков (1915) предположил, что эволюция беспозвоночных с внешним скелетом в сторону уменьшения размеров связана с механическими свойствами внешнего хитинового скелета.
Размах крыльев у древней стрекозы меганевры (290 млн. лет назад) достигал 75 см, у современных стрекоз—1–11 см.
Половой диморфизм по размеру тела
Эволюция большинства групп позвоночных сопровождалась, как правило, укрупнением размеров, а многие группы насекомых и паукообразных, наоборот—мельчали. Филогенетическое правило полового диморфизма позволяет предсказывать превалирующее направление полового диморфизма в этих группах: более крупные, по сравнению с самками, самцы должны встречаться у крупных позвоночных, а более мелкие—у мелких форм насекомых и паукообразных.
Те же тенденции должны наблюдаться и внутри более мелких таксонов, скажем в классе млекопитающих: у крупных форм чаще крупнее самцы, а у мелких—самки. Например, самцы африканского саванного слона весят до 6.5 т, в то время как самки весят только до 3.5 т. У мелких форм—некоторых летучих мышей, белок-летяг, карликового мангуста, кролика и других самцы чаще бывают мельче самок. Та же закономерность прослеживается и внутри одного отряда. Данные по приматам (Рис. 10.1) полностью подтверждают предсказанные теорией зависимости полового диморфизма по размерам от массы животных и моногамной или полигамной структуры популяции.
Р
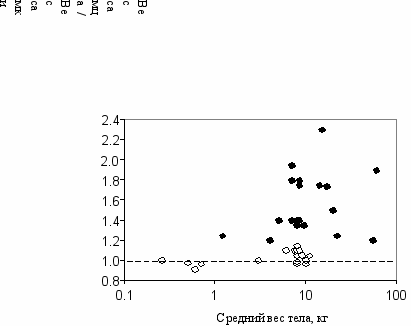 ис. 10.1
ис. 10.1Зависимость полового диморфизма
у приматов от размеров тела
(Clutton-Brock, Harvey, 1977)
○—моногамные виды,
●—полигамные.
Околоводные виды грызунов, берущие свое начало, видимо от “сухопутных”, в процессе адаптации к полуводному образу жизни должны были увеличить размеры тела. У четырех изученных видов самцы крупнее самок (Пантелеев, 2003, Corbet, 1977), а в отношении ондатры (Ondatra zibethicus) и водяной полевки (Arvicola amphibius) имеются палеонтологические сведения об их происхождении от более мелких предковых форм (Агаджанян, 1993, 2001). Напротив, у древесно-лазающих видов—белки (Sciurus vulgaris) и азиатского бурундука (Tamias sibiricus) самцы меньше самок (Пантелеев, 2003; Degn, 1973; Vericad-Corominas, 1970).
Дивергентная и конвергентная эволюция
При любой дивергенции или дизруптивном отборе, дивергенция мужского пола также должна опережать дивергенцию женского. Поэтому появление бимодального распределения самцов при мономодальном распределении самок (Алтухов, Варнавская, 1983) говорит о начавшемся дизруптивном отборе. По этой же причине экологические правила Брегмана и Аллена четче должны выполняться для мужского пола. Можно предположить, что длина выступающих частей: конечностей, ушей и хвостов у самцов арктических популяций должна быть относительно короче, чем у самок тех же видов, в то время как у самцов тропических популяций, наоборот, относительно длиннее, чем у самок.
Интересная адаптация к жаркому климату пустынь выработалась у рябков—транспортировка воды для птенцов в смоченных перьях. Видоизменившиеся брюшные перья позволяют им удерживать в 3–4 раза больше воды, чем другие птицы. При этом оперение самцов удерживает в 1.5 раза больше воды, чем оперение самок.
Процессы, приводящие к возникновению полового диморфизма могут также участвовать в видообразовании и морфологических различиях родственных видов, как изначально предполагал Ч. Дарвин. При этом, у раздельнополых организмов с хорошо выраженным половым диморфизмом самцы родственных видов животных (как позвоночных, так и беспозвоночных), часто очень сильно отличаются друг от друга, тогда как самки могут быть практически не различимы. Сходная картина биоразнообразия наблюдается и у растений: часто диверсифицированы мужские растения (у двудомных таксонов) или мужские части цветка (у обоеполых цветков) (Willson, 1991).
Филогенетическое правило полового диморфизма можно проверить на видах, эволюция которых хорошо известна (слон, лошадь, верблюд). Так у слонов имели место укрупнение размеров, развитие хобота, оригинальная специализация зубной системы, переход от бугорчатых зубов к складчатым и образование бивней. Значит можно предсказать наличие полового диморфизма по всем этим признакам. И оно во многом подтверждается.
Лошадь и осёл, возникли дивергентно ~4-8 тыс лет назад от общего предка тарпана. После дивергенции лошадь стала длиннее, выше, быстрее, хвост из длинных волос. У осла удлинились уши, укрупнилась голова, сузились копыта, утончился хвост с кистью из волос. Оба их гибрида: мул (отец осёл), и лошак (отец лошадь) похожи на отцов.
Правило справедливо также для растений. Например, существует половой диморфизм по форме листьев. У тополя женские экземпляры имеют более продолговатые листья, мужские—более округлые. У дерева гинкго листья женских особей имеют ровные края и мельче, а мужские—изрезанные и крупнее. Как известно, филогенетические предшественники тополя имели узкие (как у ив) листья, а гинкго—неизрезанные.
В процессе эволюции у покрытосеменных растений происходила редукция гаметофита (Рис. 6.4). При этом, в полном соответствии с теорией, редукция мужского гаметофита зашла дальше, чем женского: мужской гаметофит состоит уже из трех клеток, в то время как женский—ещё из восьми (Вилли, Детье, 1975).
Поскольку искусственная эволюция (селекция) домашних животных направлялась человеком, можно предположить, что хозяйственно-ценные признаки должны быть более развиты у самцов. Действительно, во всех случаях, когда признак присутствует в фенотипе обоих полов, у мужского пола он более продвинут. При мясном направлении селекции в животноводстве, как правило, самцы дают намного больше мяса (особенно при их кастрации), лучшего качества, с более высокой оплатой корма и динамикой привеса, чем самки. В тонкорунном овцеводстве более продуктивны бараны и валухи (кастраты). От них получают иногда в 1.5–2 раза больший настриг высококачественной шерсти, чем от ярок. В коневодстве также более ценным является мужской пол, так как он выгодно отличается от женского при использовании в качестве как спортивных, так и рабочих животных. То жe самое можно сказать о верблюдах и оленях. В пушном звероводстве самцы дают шкурки лучшего качества. В шелководстве продуктивность самцов выше на 20–25%, чем у самок. У растений, мужские экземпляры конопли дают в полтора раза больше волокон, чем женские.
У кур была сделана попытка вывести породы с высокой и низкой частотой копуляции. После 20 поколений у самцов наблюдалось существенное изменение частоты копуляций, в то время как у самок не было никаких изменений (Halliday, Arnold, 1987; Cheng, Siegel, 1990).
Таким образом, филогенетическое правило полового диморфизма полностью подтверждается на группе хозяйственно-полезных признаков, проявляющихся у обоих полов.
Филогенетическое правило полового диморфизма позволяет предсказать связь между половым диморфизмом и явлением гетерозиса. Рассматривая гетерозис как суммацию новых эволюционных достижений, приобретенных дивергентно, можно предположить, что, с одной стороны, вклад отца в гетерозис должен превышать вклад матери, с другой стороны, эффект гетерозиса четче должен проявляться у сыновей, чем у дочерей. Это предсказание теории также подтверждается (Levine, Cormody, 1967).
При конвергентной эволюции, наоборот самцы должны быть уже одинаковы по признаку, а самки ещё—разные. Такая картина бывает в одномерных нишах, где доминирует один фактор среды: холод за полярным кругом, или жара в пустыне.
Паразитические формы. Известно, что эволюция многих паразитических форм шла как бы в двух направлениях. С одной стороны такие виды выработали целый комплекс признаков, обеспечивающих поиск особей вида(ов)-хозяина и совместное существование. С другой стороны многие признаки, органы и даже системы органов подвергались редукции (Zimmer, 2001). Правило предсказывает, что как в случае развития новых признаков, так и редукции уже имеющихся, самцы должны опережать самок, поэтому по исчезающим признакам самцы должны быть более примитивны. Например, самка морского червя бонелия (Bonellia viridis) сохранила все функции, а самец, утратив их, стал мелким (1-3 мм) паразитом, живущим на её теле или в её половых путях.