
Роуз С. Р79 Устройство памяти. От молекул к сознанию: Пер с англ
| Вид материала | Решение |
СодержаниеШестой критерий1: биохимия коррелирует с нейрофизиологией Конец рассказа? Пятый критерий Двойные волны Порядок из хаоса? |
- «хм «Триада», 9393.37kb.
- Анастази А. А 64 Дифференциальная психология. Индивидуальные и групповые разли- чия, 11288.93kb.
- Роджер Л. М2Э вирус ответственности.: Пер с англ, 2943.44kb.
- Новые поступления литературы (июль сентябрь 2002) математика инв. 62350 в 161., 125.41kb.
- Указатель произведений литературы зарубежных стран (библиотека кф ат и со), 250.17kb.
- Литература для клинических интернов по специальности «терапия» Кафедра факультетской, 55.33kb.
- Куртц П. К93 Искушение потусторонним: пер с англ, 7904.74kb.
- The guilford press, 6075kb.
- The guilford press, 6075.4kb.
- Жизнь счастливого человека (А. Маслоу), 76.37kb.

Шестой критерий1: биохимия коррелирует с нейрофизиологией
1
Занятия нейрофизиологией требуют целого ряда навыков: не только умения искусно оперировать мелких животных, но и довольно основательных познаний в электронике, что недоступно простым биохимикам вроде меня. Мои возможности в электротехнике ограничиваются тем, что я могу подсоединить провод к штепселю, но и это нам запрещается (официально) делать в лаборатории: инструкция по технике безопасности требует, чтобы столь квалифицированную работу выполнял электрик-профессионал. Для подхода к шестому критерию мне был нужен человек, владевший техникой осциллоскопии, но только в середине 80-х годов к нам поступил для подготовки диссертации на степень доктора философии толковый, хотя и несколько чудаковатый Роджер Мейсон, заядлый аквалангист.

*1) Я прекрасно знаю, что за четвертым должен следовать пятый. Причина, по которой я откладываю обсуждение пятого критерия, скоро станет понятной.
Милтон-Кинс находится настолько далеко от моря, насколько это возможно в Англии, и я никогда не мог до конца понять, что заставило Роджера избрать именно это место (думаю, не вполне понимал это и он сам, потому что после четырех лет напряженных исследований он написал «черновик» своей диссертации на нескольких сотнях страниц — гораздо больше, чем требуется, но так и не представил ее к защите). По прибытии в лабораторию Роджер тут же исчез среди проводов и проблесковых ламп, звуковых сигналов, бесконечных рулонов регистрационных лент с десятками метров разноцветных записей. Приближаться к его рабочему месту было просто опасно, потому что приходилось прокладывать путь через свисающий откуда-то привод к аквалангу и валяющиеся на полу остатки разобранных велосипедов1. Но примерно после 18 месяцев технического затворничества он предстал перед нами, успешно справившись с задачей обеспечить эксперименты оборудованием.
*1) Сочетание современной оснащенности и домашнего беспорядка весьма типично для многих лабораторий. Непосвященного это просто изумляет, и он вправе усомниться, что из всего виденного вообще может получиться что-нибудь путное. Недавно я посетил один из ведущих мировых центров позитронно-эмиссионной томографии. В этом комплексе, сооружение которого обошлось в миллионы фунтов стерлингов, ничтожные объемы воды или органического вещества помещают в центральную камеру циклотрона и бомбардируют потоками разогнанных до огромной скорости ионов, что приводит к образованию короткоживущих изотопов. Эти изотопы по защищенным свинцовым покрытием трубопроводам перекачивают в радиохимическую лабораторию, где на автоматической установке в считанные мгновения осуществляется сложный синтез высокочистых меченых соединений. Затем полученные вещества передаются в комнату, где находится больной, голова которого окружена кольцом детекторов позитронов, которые в свою очередь соединены с системой компьютерных банков данных. Прежде чем вступить в этот мир научной фантастики, приходится облачаться в лабораторный халат и надевать на обувь защитные чехлы. Но, войдя в лабораторию, натыкаешься то на оставленные в беспорядке скамьи, то на другие тривиальные следы повседневной работы и жизни: шпатели и ножницы, торчащие из пустой банки из-под меда; кофеварку, кем-то поставленную прямо у экранированного шкафа; тайком курящего техника, обслуживающего оборудование. Как уживается весь этот хаос с фантастическим миром, где все измеряется наносекундами и пикограммами?
В сущности, мы собирались делать очень простые вещи. Нужно было предложить цыпленку бусину, смоченную водой или метилантранилатом, а потом усыпить его. Затем его помещали в так называемый стереотаксический аппарат с миниатюрной системой жизнеобеспечения. Здесь наркотизированного птенца аккуратно, но жестко закрепляли таким образом, чтобы в его обнаженный мозг в соответствии с заданными координатами можно было ввести электроды (конечно, теперь это уже не животное, а «препарат», у которого исчезнут признаки жизни, стоит только отключить систему жизнеобеспечения). Есть разные типы электродов: тонкие стеклянные трубочки, заполненные растворами солей или активных веществ, которые требуется подвести к определенным клеткам мозга, или так называемые «стимулирующие электроды», с помощью которых подводят залпы электрических импульсов. Наконец, электродами могут служить тонкие металлические проволочки, назначение которых — всего лишь регистрировать электрическую активность близлежащих клеток. В наших экспериментах использовались электроды этого последнего типа: мы хотели выяснить, изменяется ли электрическая активность нейронов в IMHV в результате ознакомления цыпленка с бусиной, смоченной метилантранилатом.
Все это выглядит довольно просто, но скрывает массу сложностей. Очень трудно оказалось подобрать подходящее наркотизирующее средство, которое не мешало бы поддерживать жизнь цыпленка в течение нескольких часов. Другая сложность состояла в интерпретации электрических сигналов и отделения их от посторонних «шумов». Каждый отдельный эксперимент может занять много часов, так как требует предварительной подготовки животного и аппаратуры; поэтому нейрофизиологи в еще большей степени, чем другие известные мне лабораторные исследователи, склонны трудиться по ночам (во всяком случае, когда работают над диссертацией) и плохо приспособлены для нормального человеческого общения.
Моя роль в этом эксперименте была несложной. Я только предлагал цыплятам клевать бусину, смоченную метилантранилатом или водой, и потом передавал их Роджеру, а тот исчезал с ними в нейрофизиологической лаборатории и много часов спустя выныривал с ворохом записей, которые начинал анализировать. До завершения первой серии экспериментов я не говорил ему, к какой группе принадлежит тот или иной цыпленок (это обычная практика в нашей лаборатории: по возможности, особенно если опыт проводят два сотрудника, мы стараемся работать «вслепую» до завершения анализа полученных данных, чтобы избежать подсознательной предвзятости в оценке). После регистрации данных для шестнадцати птенцов Роджер сообщил, что выявил закономерные различия между ними, столь значительные, что может подразделить всех цыплят на две группы без моей подсказки. Когда я попросил его сделать это, он правильно определил 14 цыплят из 16.
Как мы и ожидали, во всех случаях регистрировался постоянный «фон», отражавший спонтанные разряды нейронов IMHV. Но на этот фон накладывались кратковременные «вспышки» высокочастотной активности — ритмического синхронного возбуждения целых ансамблей клеток (рис. 10.10). Эта активность у цыплят, клевавших бусину с метилантранилатом, была выражена намного (иногда вчетверо) сильнее, чем у контрольных особей, клевавших бусину, смоченную водой. Превышение могло сохраняться на протяжении суток после тренировки. Все это действительно несколько напоминало эффект ДВП, только вызывалось не искусственным пропусканием тока, а приобретенным поведенческим опытом [22]. Возможно, что существует и полная аналогия: спустя пару лет другим исследователям удалось получить ДВП-подобные явления при стимуляции срезов IMHV in vitro [23]. Чтобы окончательно убедиться в специфичности «вспышек», мы с Роджером повторили опыт, используя описанную выше индукцию амнезии электрошоком.
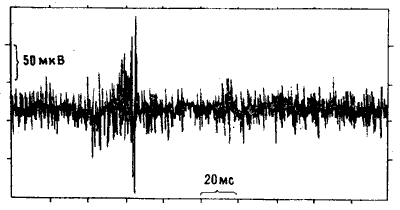
Рис. 10.10. Пульсирующая активность нейронов IMHV. Запись активности группы нейронов IMHV у наркотизированного цыпленка. По вертикальной оси отложены потенциалы, по горизонтальной — время. Можно видеть интенсивную фоновую активность с довольно низкой амплитудой и высокочастотный разряд с амплитудой до 300 микровольт. Такая пульсирующая активность резко усиливается после обучения.
И снова ритмические вспышки, подобно биохимическим и структурным изменениям, отмечались только у тех цыплят, которые помнили усвоенную задачу [24].
Конец рассказа?
Итак, для того чтобы в мозгу цыпленка сформировалась ассоциативная связь между клеванием бусины и горьким вкусом, приводящая к стойкому изменению поведенческой реакции, в определенной области переднего мозга должен произойти ряд последовательных событий. Эти события завершаются структурной модификацией синапсов и дендритов, и в итоге проявляются также в изменении электрических свойств клеток, в частности в характере их ритмической активности на протяжении нескольких часов после обучения. Все это, видимо, удовлетворяет шестому критерию.
Таким образом, узнал ли я, наконец, как и где образуются у цыплят следы памяти? Отчасти, уважаемый читатель, только отчасти. Вся эта биохимия и нейрофизиология, все структурные изменения — прекрасны: десять лет славной экспериментальной работы, внесшей некоторый порядок в кажущийся хаос живого мира. Я чувствую, что меня не ввели в заблуждение артефакты и что я правильно интерпретировал свои результаты, хотя даже мне самому, не говоря уже о посторонних критиках, ясно, что, анализируя цепь событий, я формально не подтвердил еще наличия всех необходимых биохимических звеньев. Некоторые из моих доводов балансируют на той опасной грани, где исследователя подстерегает классическая ловушка «post hoc ergo propter hoc», хотя из того, что фосфорилирование предшествует синтезу гликопротеинов, не следует автоматически, что последний зависит от первого. Однако это, вероятно, не самый существенный вопрос. Гораздо важнее выяснить, действительно ли, даже без всей этой биохимии, память — столь простой механический процесс, незамысловатое связывание нейронов в новую сеть в IMHV, нечто вроде переключения элементов компьютера. Значит ли это, что прав был Хебб? Специфичны ли обнаруженные мною эффекты для кур и даже только для цыплят, запоминающих горький вкус бусины, или же я могу с полным правом утверждать, что они иллюстрируют какие-то общие принципы формирования следов памяти? Не должен ли удивлять сам масштаб наблюдавшихся явлений? Четырехкратное усиление ритмической активности, 60%-ное увеличение числа шипиков на дендритах — и все это только для того, чтобы запомнить маленькую бусинку? Если такое будет случаться всякий раз, когда курице придется что-то запоминать на протяжении всей ее жизни, то где в ее крошечном мозгу найдется место для всех этих синаптических перестроек?
Если бы сам я не пытался ответить на такие вопросы, их, несомненно, поставил бы кто-нибудь другой. Правда, моя повседневная лабораторная работа проходит в мире артефактов, создаваемых техническим оснащением. У меня нет возможности непосредственно наблюдать природу хотя бы на примере моих цыплят. Подобно всем научным данным, мои данные на самом деле не что иное, как результаты регистрации измерений, записи на бумажных лентах, цифры на шкале или экране приборов (философ-позитивист и физик Эрнст Мах в начале века назвал такие наблюдения «показателями стрелок», я манипулирую ими, пытаясь понять их значение, а поняв, произвожу обратную экстраполяцию, чтобы сделать выводы о поведении молекул, клеток и организмов в реальном мире. Вместе с тем меня совершенно не трогает текущая полемика в кругах философов и социологов относительно статуса реализма и науки. Я придерживаюсь того, о чем уже писал раньше; это правда о том, что я наблюдал в изучаемой мною материальной вселенной. Всякий, кто возьмется организовать такую лабораторию, как моя, и провести те же эксперименты, получит сходные результаты, ибо они не плод тайного искусства или трюкачества, и наука, по мнению страстно приверженных ей философов — это все же публичное знание. Но то, что описано мною, — это правда в моем истолковании, и люди, получившие от меня новое знание, должны будут усвоить и мои представления (или по крайней мере значительную их часть) о том, как следует его интерпретировать. К тому же это не вся правда: описание экспериментов избранным мною способом (как я говорил в начале этой главы) является лишь логическим вариантом подачи материала, еще не получившим достаточного теоретического обоснования. Это был риторический прием, хотя и необходимый (как убеждают меня мои друзья-литераторы, потому что именно так распространяется научное знание), но все же риторический прием. Теперь позвольте мне начать новую главу и поведать еще одну историю.

Глава 11
Порядок, хаос, порядок: критерий пятый

Пятый критерий
В восьмидесятые годы результаты наших экспериментов так хорошо согласовались с представлениями о каскаде клеточных процессов, что мне на ум снова и снова приходил мой пятый критерий: удаление анатомического участка, в котором происходят биохимические, клеточные или физиологические изменения, должно препятствовать образованию следов памяти и/или вспоминанию в зависимости от того, когда по отношению к времени тренировки произведено удаление.
Для этого были три причины. Во-первых, я знал, что нам нужно будет провести соответствующий эксперимент, который потребует освоения совершенно новых методов. Во-вторых, я не мог без волнения думать о возможных результатах. И наконец, в-третьих, как я говорил раньше, меня всегда тревожила эстетическая и отчасти даже моральная сторона опытов, связанных с травмированием живых существ, а также проблема интерпретации получаемых данных. Эту проблему мне рано или поздно придется решать. В 1988 году Сэри Дейвис, бывший студент Габриела Хорна, работающий сейчас в Лондоне, опубликовал статью об экспериментах с повреждением левого и правого IMHV у цыплят. Он производил операцию в день их вылупления, а на следующий день обучал пассивному избеганию; после этого они клевали бусину, проявляли недовольство ее вкусом и во всем остальном вели себя нормально, однако у них обнаруживалась амнезия: они вторично клевали горькую бусину [I]1. Нечто в этом роде наблюдал и сам Габриел при изучении импринтинга. Всего этого можно было ожидать, исходя из биохимии, морфологии и нейрофизиологии IMHV после обучения, но мы должны были провести более систематические эксперименты. Я решил уделить этому большую часть 1989 и 1990 годов, так как получил исследовательский грант, позволивший мне намного сократить объем преподавательской и административной работы. Ниже излагается история этих двух лет с их итогами.
*1) Меня не перестает удивлять, что цыплята, у которых повреждена (или удалена, что безразлично) такая сравнительно большая область мозга, все же не погибают и по видимости сохраняют нормальное поведение. Скептики могли бы задаться вопросом, что же делает этот мозг большую часть времени. На это фермер-птицевод мог бы ответить, что единственная функция мозга состоит в том, чтобы удерживать птиц от непрерывной беготни, — это известно с незапамятных времен, с тех пор как первый крестьянин отрубил голову первому петуху, чтобы положить его горшок.
Мы смогли воспроизвести результаты Сэри, а потом продолжить исследования на их основе благодаря приезду в лабораторию двух очень разных физиологов, уже имевших ученую степень, — Терри Паттерсон и Дейва Гилберта. Терри только что защитила диссертацию, работая на цыплятах с Марком Розенцвейгом в Беркли. Карьера Дейва после получения степени в Бирмингеме была не совсем гладкой, и к тому времени, как я получил субсидию, он явно недоиспользовал свои возможности в качестве доктора философии, перебиваясь мытьем витрин, поскольку получить деньги на научную работу в Англии восьмидесятых годов было довольно трудно. Практически с момента появления Терри и Дейва в лаборатории между ними возникла личная неприязнь, но и как экспериментаторы, и в теоретическом плане они составляли великолепную пару. Мы спланировали эксперименты так, чтобы один производил повреждение мозга, второй обучал цыплят, не зная о характере повреждения, а третий (обычно это был я) проводил последующие испытания, опять-таки не зная о сделанном двумя другими.
Мозг повреждали так же, как в нейрофизиологических экспериментах. Цыпленка наркотизировали и стереотаксически вводили в мозг электрод (тонкую проволочку) так, чтобы его кончик оказался в нужном месте. После этого пропускали ток, разогревавший кончик электрода. Под действием повышенной температуры (или высокочастотных колебаний) клетки вокруг электрода погибали; размеры повреждения контролировали, изменяя силу тока и продолжительность воздействия. Затем электрод извлекали, кожу на голове зашивали и цыпленку давали возможность оправиться от наркоза в течение ночи. Для оценки влияния самой операции и наркоза в каждой подопытной группе имелись цыплята, подвергавшиеся «ложной» операции: с ним проделывали все, что и с остальными, но через электрод не пропускали ток. Все это тонкая работа, но при необходимых навыках за рабочий день можно прооперировать до дюжины цыплят. Они хорошо переносили операцию и наркоз: придя в себя, цыплята с поврежденным IMHV или LPO выглядели совершенно нормальными и не отличались ни от ложно оперированных, ни от интактных контрольных особей. После оценки поведения цыплят забивали, а поврежденные участки мозга исследовали под микроскопом. Как и в других случаях, для получения статистически достоверных результатов эксперимент многократно повторяли, чтобы в каждой опытной (не контрольной) группе насчитывалось двенадцать или более цыплят. При четырех рабочих днях в неделю (это дни, когда вылупляются цыплята) с учетом времени, затрачиваемого на ложные операции и на проверку локализации повреждений, для получения одной серии результатов требуются примерно три недели; практически же на это уходит около месяца, поскольку что-то всегда идет не так: то цыплята не вылупляются, то нужно идти на какое-то заседание и т.п.1.
*1) Я давно уже на собственном печальном опыте убедился, что при планировании эксперимента нужно взять максимальное потребное время, удвоить его и добавить еще немного — тогда ваши расчеты окажутся более или менее верными. Следует учитывать еще один важнейший момент если вы налаживаете метод, которым раньше не пользовались, а позаимствовали из чьей-то статьи, он обычно начинает работать не раньше чем с третьей попытки; в двух первых вы каким-то непонятным образом обретаете в кончиках пальцев ту магическую, не поддающуюся описанию силу («безмолвное знание», как назвал ее Майкл Полани, некогда химик, а потом философ), которая позволяет освоить метод не только в теории, но и на практике.
Потратив нужное число месяцев и сделав достаточно ошибок, чтобы освоить методику повреждения мозга, мы смогли приступить к первому этапу — к проверке результатов Сэри. В начале 1989 года мы подтвердили их. Сомнений быть не могло: цыплята с поврежденным IMHV явно обучались пассивному избеганию, т. е. клевали горькую хромированную бусину, так же, как их ложно оперированные собратья, трясли головами, ощутив ее вкус, и отворачивались при повторном предъявлении. Но спустя несколько часов при вторичном испытании они полностью забывали приобретенный опыт и клевали сухую бусину столь же энергично, как и контрольные цыплята, которым раньше давали бусину, смоченную водой. Таким образом, повреждение мозга не влияло на поведение птенцов (клевание бусины), на чувство вкуса или общую подвижность — они лишь не помнили, что следует избегать бусин определенного вида. Теперь нужно было пойти дальше Сэри и производить одностороннее повреждение. И снова мы получили ожидавшийся результат. У цыплят с поврежденным левым IMHV реакция избегания отсутствовала, а при правостороннем повреждении полностью сохранялась. Все было нормально — в полном соответствии с прогнозом, основанным на ранее полученных данных, и с пятым критерием, для запоминания необходим был левый, но не правый IMHV (рис. 11.1).
Рис. 11.1. Повреждение IMNV. На этом и следующих рисунках затемненные участки — места повреждения.
По логике нашего подхода следовало теперь проверить, что произойдет, если повредить мозг после обучения. Это можно было сделать не раньше чем через час, так как иначе на результатах испытаний могло бы сказаться действие наркоза. К нашему удивлению, даже двустороннее повреждение мозга спустя час после обучения не приводило к амнезии. Итак, первый парадокс: для запоминания необходим интактный левый IMHV, но коль скоро цыплята уже усвоили задачу (по крайней мере в первый час после обучения), IMHV оказывался ненужным [2].
Куда же переместилась память? Учитывая все полученные ранее биохимические и морфологические данные, следовало искать ее следы в LPO. Эксперимент подтвердил это. Двустороннее повреждение LPO через час после обучения действительно вызывало амнезию, которой не было при одностороннем правом или левом повреждении. По-видимому, в норме след памяти после обучения каким-то путем мигрирует из IMHV в LPO, чем и объясняется отсутствие амнестического эффекта повреждения после тренировки. Это был весьма интересный результат, так как он позволял понять, почему мы находим биохимические и морфологические изменения и в IMHV, и в LPO. Тот факт, что для сохранения памяти достаточно одного LPO, правого или левого (одностороннее повреждение не вызывало амнезии!), согласовался с тем, что многие из выявленных нами изменений имели место как в правом, так и в левом LPO, т. е. след памяти о бусине и реакции избегания сохранялся в обоих полушариях (рис. 11.2).
Не довольствуясь столь простым результатом, мы пошли дальше. Что будет, если повредить LPO до обучения? К нашему удивлению, такая операция никак не влияла на память [3] (рис. 11.2).
Рис. 11.2. Повреждение LPO.
Если пассивное избегание вырабатывается и при отсутствии LPO, то не происходит ли реорганизации следа памяти каким-то иным образом? Может быть, в этом случае след просто закрепляется в IMHV? Если это так, то повреждение LPO до тренировки и повреждение IMHV после тренировки - две операции, по отдельности не приводящие к амнезии, - при их сочетании у одного и того же цыпленка должны будут вызвать амнезию. Так оно и оказалось в действительности (рис. 11.3). Получив такие результаты, я предположил, что решающая роль принадлежит IMHV. Но Дейв думал иначе. Тогда мы произвели односторонние повреждения, и выяснилось, что он прав. У цыплят, не имевших LPO во время обучения, последующее повреждение левого IMHV не влияло на запоминание, а повреждение правого IMHV вызывало амнезию. Все это схематически показано на рис. 11.3.
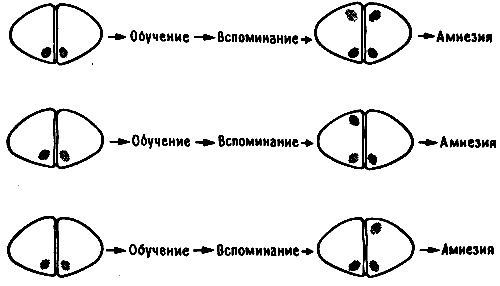
Рис. 11.3. Повреждение LPO до обучения и LMHV после него.
Результаты были интригующими. Размышляя над ними, я решил, что все можно объединить, исходя из несколько механистической модели, в которой первичный след памяти о бусине и реакции избегания образуется в левом IMHV, а затем, спустя несколько часов после обучения, «перекочевывает» сначала в правый IMHV, а потом в правый и левый LPO (рис. 11.4).
Рис. 11.4. Объяснение результатов: возможные перемещения следов памяти.
Я пришел к этой схеме исключительно на основании логики экспериментов с повреждением мозга. Но подобная схема шокирует нейроанатомов, у которых тут же возникает очевидный вопрос: разве есть какие-то прямые нервные пути между IMHV и LPO? Согласно простой концепции коннекционизма, такие пути действительно необходимы. В то время, когда мы приступали к исследованию, мы не знали, существуют ли они, но это казалось маловероятным. Нам было известно, что все нервные пути от органов чувств, таких как глаза и вкусовые рецепторы, сходятся к IMHV. Мы знали даже о непрямых связях между IMHV и LPO. Именно поэтому IMHV рассматривались как некие «ассоциативные зоны», где интегрируется разнообразная входная информация - например, сопоставляется зрительное восприятие бусины с ощущением ее вкуса. Напротив, LPO находятся на выходной стороне мозга, в области, ответственной за двигательные функции (как, например, клевание) и, возможно, также за реакции «эмоционального» типа. После начала наших опытов анатомия мозга цыплят стала несколько яснее. Между IMHV и LPO действительно нет простых связей, хотя, по-видимому, существует непрямой путь через третий отдел мозга - так называемый архистриатум.
Но и без учета анатомических данных приведенная гипотетическая схема (рис. 11.4) перемещения следов памяти позволяет сделать ряд предсказаний. В частности, из нее следует, что повреждение правого IMHV до начала обучения (само по себе не вызывающее амнезии) должно прерывать последовательность событий, и тогда след памяти «застрянет» в левом IMHV, не имея возможности переместиться в LPO. Повреждение после обучения, обычно приводящее к амнезии, теперь уже не вызовет ее. Высказав это предположение, мы тут же (во всяком случае не позднее чем через месяц) проверили его (рис. 11.5).
Рис. 11.5. Блокирование перемещений.
Оно подтвердилось. Тогда Дейв, Терри и я решили опубликовать обобщающую статью с описанием всех новых экспериментов. Мы быстро подготовили черновой вариант, но, когда уселись, чтобы вместе доработать его, кто-то (сейчас уже не помню кто) неожиданно сказал: «А что, если до обучения повредить правый IMHV?». Мы знали, что само по себе такое повреждение не дает амнестического эффекта, а наша модель предсказывает, что след памяти в этом случае должен оставаться в левом IMHV -- просто из-за отсутствия выхода. Поэтому у цыплят с поврежденным до обучения правым IMHV повреждение левого IMHV после обучения приведет к амнезии. Такой вариант опыта мог действительно стать окончательной проверкой. Мы отложили статью в сторону и потратили еще месяц на новый эксперимент. Вот что мы получили (рис. 11.6):
Рис. 11.6. Но след находит другой путь...
До этого я был готов, как некий научный Пуаро, собрать всех подозреваемых в гостиной и указать виновного, как было совершено преступление, то бишь как работает память. Теперь это было исключено. Эксперимент дал однозначный результат - целиком отрицательный. Ничего, пустота, нуль, никакой амнезии. След памяти не мог перейти в правый IMHV, но он определенно не остался и в левом. Куда же он подевался?
Почти полвека назад психолог Карл Лэшли написал классическую статью, обобщив результаты десятилетних экспериментов с обучением крыс. Он обучал их ориентироваться в сложных лабиринтах, а потом удалял отдельные участки мозговой коры, чтобы выяснить, где сохраняются следы памяти. К своему удивлению, он не обнаружил специфических участков, удаление которых приводило бы к полной утрате усвоенного навыка; вместо этого навык постепенно ухудшался по мере увеличения размеров удаленной области коры. Создавалось впечатление, что память зависит просто от количества мозгового вещества. На основе этих наблюдений Лэшли построил концепцию «эквипотенциальности» коры. Свою статью он озаглавил «В поисках энграммы» и закончил ее выводом, что память одновременно присутствует в мозгу везде и нигде [4].
Опыты Лэшли и его неутешительные выводы утратили актуальность в результате последующих экспериментов, однако парадокс локализации памяти остался. Это напоминает мне, во-первых, о том, что при повреждении IMHV и LPO мы изучаем не функции этих областей, а функции остального мозга, которые быстро перестраиваются в отсутствие двух первых; и, во-вторых, о том, что память нельзя понимать как нечто статичное, «находящееся» строго в одном месте или в небольшой группе клеток; она существует в более динамичной и рассеянной форме. Кроме того, мозг - это тонко и сложно организованная структура с многообразными средствами функционального обеспечения. Вы мешаете ему осуществлять какую-то деятельность, а он находит другие пути к той же цели. Блокируете оптимальные связи между IMHV и LPO, a цыпленок находит им замену. Мозг действует не как набор стандартно соединенных мелких ячеек, а как функциональная система, насыщенная разнообразными связями. Конечно, всегда следует помнить об анатомии мозга, но нельзя замыкаться в ней, поскольку ясно, что и сам мозг не сводится к анатомически выявляемым структурам. Настаивать на его механистическом объяснении, игнорируя мои предостережения о неразумности редукционизма, - это значит обрекать эксперимент на погружение в трясину парадоксальных результатов.
Двойные волны
Мысль о том, что у нормальных цыплят при обучении возникает своего рода поток запоминаемой информации из левого IMHV в LPO, приводит к еще одному парадоксу и подсказывает новые эксперименты. Вся совокупность накопленных за десятилетие биохимических и морфологических сведений, рассмотренных в предыдущей главе, сводится к тому, что у цыплят, обучавшихся избегать горькую бусину, происходят долговременные изменения в химии и структуре клеток LPO и левого IMHV. Однако опыты с повреждением мозга, по-видимому, показали, что обученному цыпленку IMHV вовсе не нужен. Почему же тогда сохраняются изменения в этой области?
Я размышлял об этом по вечерам, а днем пытался заполнить пробелы в знаниях, занимаясь биохимией. Если результаты опытов с повреждением мозга отражали реальные события, то вскоре после обучения должны были происходить биохимические изменения в левом IMHV, а позже такие же изменения должны появляться и в LPO. Значит, наряду с потоком информации из IMHV к LPO должен существовать «поток» биохимических изменений. Неожиданно я вспомнил о странных результатах, полученных магдебургской группой лет десять или больше назад в опытах на крысах, обучавшихся различению яркости. Я снова просмотрел статьи этих исследователей и убедился, что память не подвела меня. Они наблюдали появление двух волн повышенного синтеза белков и, в частности, гликопротеинов: первой в гиппокампе, а второй, примерно через шесть часов после обучения, - в коре мозга. В то время никто не знал, что можно извлечь из этих данных, и авторы не пошли дальше. В моей памяти начали всплывать разрозненные сведения, почерпнутые на недавних конференциях, которые в свое время, казалось, не представляли существенного интереса. Как их проверить?
В нашей биохимической лаборатории мы еще изучали вопрос о гликопротеинах, используя ингибитор их синтеза 2-дГал. Работая ранее с Рейнхардом Йорком, я показал, что если вводить 2-дГал в мозг в любое время от двух часов до обучения до двух часов после него, у цыплят развивается амнезия. Однако она не развивалась при инъекции спустя 3 часа после тренировки. Я не стал тогда изучать эффект введения 2-дГал в более поздние сроки, так как не видел в этом особого смысла, но теперь решил попробовать. Я повторил эксперимент, обучая цыплят по утрам, а затем вводя отдельным группам в разные сроки (с интервалом в 1 час) вплоть до 12 часов 2-дГал или для контроля физиологический раствор. На следующее утро, через 24 часа после обучения, я проверял их поведенческую реакцию. Полученные результаты представлены на рис. 11.7.
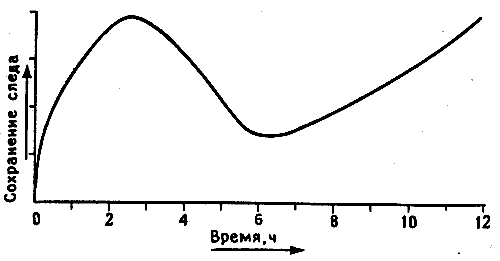
Рис. 11.7. Двойная волна памяти. В этом опыте цыплятам в разное время до и после обучения (О) вводили ингибитор синтеза гликопротеинов - 2-дГал - и спустя 24 часа после тренировки тестировали память. Запоминание отсутствовало, если 2-дГал вводили в первые 1-2 часа после обучения или спустя 5-7 часов после него. Амнестический эффект не наблюдался при введении 2-дГал в интервале между этими сроками или поздне 8 часов после обучения.
Первая часть кривой повторяет картину, выявленную раньше: инъекция 2-дГал вблизи периода обучения вызывала амнезию, а инъекция спустя 3 часа или позднее не давала эффекта. Это было так, пока я не подошел к шестичасовой отметке: тогда введение 2-дГал стало сопровождаться второй волной амнезии. Таким образом, в период после обучения должны быть две волны синтеза гликопротеинов: одна сразу после обучения, а вторая спустя несколько часов. Не локализована ли первая волна в IMHV, а вторая в LPO? Может быть, синтезируются разные гликопротеины? Это как раз те вопросы, что я изучал в эксперименте, который описан в главе 2, когда весной 1992 года начал писать эту книгу. Больше года нас осаждали бесчисленные технические проблемы. Я возненавидел эти проклятые гликопротеины и заодно себя за то, что вспомнил о двойной волне. Но я не собираюсь останавливаться на полпути, хотя ответ на поставленные вопросы лишь смутно брезжит где-то вдали. Сейчас, когда я пишу эти строки, выясняется, что мы скорее всего правы: две волны различны, и вторая локализована в LPO. Но время, отпущенное мне на исследования, на исходе, и я опаздываю с представлением этой рукописи. Если ко времени получения корректуры у меня появятся новые данные, я постараюсь вставить их в книгу1.
*1) Я сейчас держу корректуру (май 1992 года) и на 90% уверен, что мы правы.
Между тем нейрофизиологические исследования тоже шли полным ходом. Место Роджера Мейсона в паутине проводов и нагромождении осциллоскопов занял - на этот раз с более сложным компьютерным оснащением - очередной диссертант Джон Джигг, изучавший временную картину ритмической активности нейронов IMHV; одновременно он не упускал из вида и LPO. Он получил необходимые данные примерно в то же время, когда мы пытались интерпретировать результаты повреждений мозга. Те и другие как будто согласовались между собой. Сразу после обучения электрические разряды возникали в обоих полушариях, но спустя 4-6 часов их усиление регистрировалось главным образом в правом IMHV и одновременно они достигали максимальной интенсивности в правом и левом LPO (рис. 11.8). Иными словами, здесь тоже были две волны активности. При наложении кривой электрической активности на двухфазную кривую биосинтеза гликопротеинов отмечалось их близкое соответствие, несмотря на некоторые расхождения во времени. Все это подкрепляло мысль о «переходе» памяти из левого IMHV в правый, во всяком случае у нормальных цыплят.
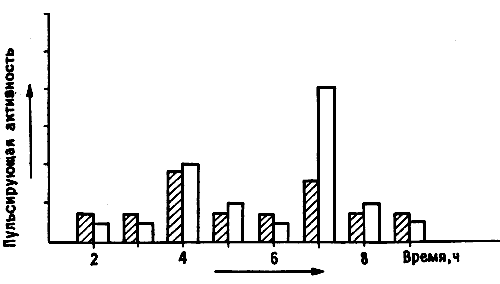
Рис. 11.8. Динамика пульсирующей активности. На диаграмме показано усиление пульсирующей активности в левом (темные столбики) и правом (светлые столбики) IMHV в различные сроки после обучения. Обратите внимание, что спустя 3-4 часа после тренировки активность возрастала на обоих сторонах мозга, а спустя 6-7 часов она усиливалась в правом IMHV гораздо больше, чем в левом.
Порядок из хаоса?
Когда вы участвуете в какой-нибудь исследовательской программе, нередко случается, что, проснувшись в понедельник утром с идеей новых экспериментов, вы мчитесь в лабораторию, чтобы тут же начать их; но проходят недели или месяцы, а у вас нет ничего, кроме неоднозначных результатов, которые никак не вписываются в общую картину, уже начавшую складываться в вашем воображении. Вы решаете прекратить опыты или опубликовать их в каком-нибудь малоизвестном журнале, надеясь, что никто не заметит, как классическое совершенство вашей теории превратилось в абстракционистский сумбур Джексона Поллока. Нечто в этом роде годами испытывали все мы в лаборатории. Но вдруг мне стало ясно, как согласовать мои теории с обнаруженной ранее двухфазной клеточной активностью. Если цыплятам вместо бусины, смоченной очень горьким метилантранилатом, предложить что-нибудь с менее резким вкусом, например ту же бусину, смоченную очень слабым раствором метилаитранилата или хинина, у них тоже появится реакция избегания и она будет сохраняться несколько часов, после чего они забудут о горьком вкусе бусины. Предположим, что эта «слабая» память связана только с первой волной клеточной активности в IMHV, которая затем не передается в форме более стабильной второй волны в LPO: такие слабые воспоминания индуцируют только первую, но не вторую волну биохимических изменений. Оказывается, дело обстоит именно так [5]. Более того, в экспериментах с выработкой «условнорефлекторного отвращения к пище», описанных в главе 6, было установлено, что даже акт клевания сухой бусины вызывает первую волну биосинтеза гликопротеинов.
Дело в том, что цыпленок все время наблюдает, замечает, исследует и запоминает различные факты окружающей его действительности, хотя бы потому, что поначалу не знает еще, что важно помнить, - у него формируется нечто вроде эйдетической памяти, как у ребенка. В обедненных условиях клетки с ее монотонным интерьером появление даже вовсе безвкусной бусины вносит новизну, которую цыпленок по меньшей мере не может не заметить; он клюет бусину, и память о ней запечатлевается, скорее всего в IMHV. Если потом, как в эксперименте с условным избеганием, цыпленок чувствует легкое недомогание, то он приходит к вполне разумному, хотя и ошибочному, выводу, что причина этого - именно клевание бусины (если, конечно, где-то в его мозгу сохранился след этого события). В очень простом эксперименте с подавлением синтеза гликопротеинов 2-D-галактозой мы показали, что для образования такого следа необходима первая волна их биосинтеза (рис. 11.9) [6].
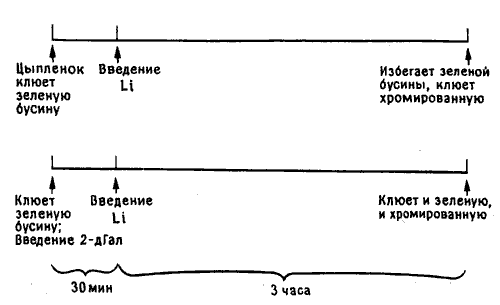
Рис. 11.9. Эксперимент с выработкой условнорефлекторного отвращения к пище.
Пока все хорошо. Теперь я могу свести воедино картину процессов кратковременной и долговременной памяти, локализованных в разных отделах мозга, объединенных потоком информации между ними и связанных с двумя волнами клеточной активности. Однако я еще не выбрался на гладкую дорогу. Долговременная память оставляет в левом IMHV длительно сохраняющийся след, но после того, как воспоминание сформировалось, этот IMHV цыпленку больше не нужен, чтобы помнить о горькой бусинке. Как это можно понять?
Я не знал этого. Когда я начал писать книгу, то собирался закончить эту главу парадоксом превращения порядка в хаос. В первоначальном варианте она оканчивалась следующими словами:
Этим рассказом я и хотел бы закончить главу. Понимайте его как хотите: с одной стороны, это серия строгих экспериментов для выявления необходимых и достаточных клеточных процессов в определенных областях мозга, которые в целом создают след простой ассоциативной памяти, изменяющий последующее поведение цыпленка; с другой стороны - параллельное описание памяти как ускользающего от наблюдения динамического процесса, столь важного для цыплят, что его не удается окончательно связать с определенными местами и «выключить».
Теперь это не годится. Это никого не устроит: ни меня, ни издателя, ни вас, моих читателей. Конечно, я мог бы сделать нечто большее. В начале этого года, совершенно неожиданно, решение стало так ослепительно ясно, что мне оставалось только изумиться, как туго моя мысль была затянута в смирительную рубаху механистического редукционизма. Все это время я рассказывал, как цыплята клевали хромированную бусину, будто она была для них простым, неразложимым объектом. Ибо мы сами считали ее такой - простой хромированной бусиной, и думали, что так же считают цыплята. Но, конечно, есть множество причин, по которым этого не должно быть. Клевавший горькую бусину цыпленок не имел понятия о том, что в ней важнее: цвет, величина или форма, время дня или место, где она появлялась. Не зная, на чем остановиться, цыпленок, для того чтобы выжить, должен был учитывать все эти особенности. Он не мог сразу же сделать вывод, что важна только бусина данной конфигурации. Поэтому цыпленку необходимо было классифицировать то, что он видел и испытывал.
Предположим теперь, что из всех свойств бусины важны цвет, размеры и форма (есть основания думать, что это действительно так, но я не буду углубляться здесь в подробности [7]). После того как цыпленок познакомился с бусиной, любой из этих признаков в дальнейшем помогает избегать ее. Это очень похоже на попытку вспомнить забытое имя: мы вызываем в воображении лицо человека, подбираем имена, которые, как нам кажется, близки по звучанию к забытому, или последовательно пробуем все буквы алфавита. Все это - разные стратегии поиска признаков забытого имени.
Если цыпленок действительно помнит не «бусину», а набор ее ключевых признаков, то последние не обязательно должны «храниться» в одном и том же участке мозга. Возможно, информация о цвете бусины сохраняется в одном месте, о форме - в другом и т. д. Просто мы никогда не пытались проверить такое предположение, а вместо этого обучали цыплят не клевать хромированную бусину и потом наблюдали их поведение, повторно предъявляя такую же бусину. Но если информация о цвете хранится в IMHV, а о других признаках в LPO, становится понятно, почему после обучения цыплятам не нужен IMHV, чтобы вспомнить бусину: в их LPO отложилась информация о других ее особенностях. В то же время в IМНV долгое время сохраняются клеточные изменения, так как «цветовое представительство» бусины все еще существует в этом отделе. Таким образом, мы нашли способ избежать парадокса Лэшли.
После того как я обдумал все это, проверка не составляла особой трудности. Нужно было лишь обучить цыплят не клевать горькую желтую бусину, а потом предложить им на выбор две бусины: ту же самую и новую - синюю. Цыплята с неповрежденным мозгом будут клевать вторую и отвергнут первую А как поступят цыплята с повреждениями? Если память о цвете хранится в IMHV, удаление LPO до обучения ни на чем не скажется: птенцы будут по-прежнему обучаться и помнить различие между бусинами, как и в предшествующих экспериментах (см. рис. 11.2). Но теперь удаление IMHV будетиметь последствия: хотя у цыплят сохранится способность у научению в последующих тестах они будут избегать обеих бусин - и желтой, и синей, так как без IMHV у них останется информация только о величине и форме бусины, находящаяся в LPO; по этим признакам они не смогут отличать одну бусину от другой и будут избегать всех мелких предметов округлой формы подозревая, что они горьки на вкус.
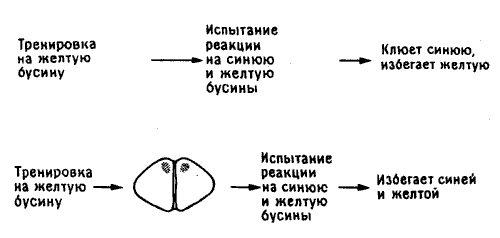
Рис. 11.10. Эксперимент с различением цветов цыплятами.
К лету 1991 года мы С Терри уже провели такой эксперимент [8]. Тем самым был найден выход из редукционистской ловушки и, признав мозг открытой обучающейся системой, мы воссоздали порядок из хаоса.
Глава 12
Интерлюдия: лабораторные исследования - это еще не все
Начинается почти всегда с одного и того же. Рано утром самолет уносит вас из Хитроу, а потом, преодолев несколько часовых поясов, вы оказываетесь у выхода из незнакомого аэровокзала, несколько обескураженные видом послеполуденного солнца, и ждете такси или, если повезет, автобуса со специальной табличкой, чтобы добраться до гостиницы, где размещаются участники конференции. Унылое чувство, которое вы пытались подавить на протяжении всего полета, начинает перерастать в досаду. Зачем вы приехали? Вы никого здесь не знаете. Программа, казавшаяся такой интересной, когда с год назад вы получили первое извещение о конференции, при более внимательном просмотре ее в самолете кажется теперь скучной: часть сообщений утратила с тех пор новизну, часть сформулирована непонятно. Место, где проводится конференция, выглядевшее довольно экзотично в рекламной брошюре, - на самом деле всего лишь шумный город в разгар лета, или же курортное местечко во внесезонное время, чтобы удешевить проживание в гостинице. И при всем этом ваша рабочая программа на дни конференции так насыщена, что не останется даже времени, чтобы побывать где-то, кроме заседаний и встреч.
Вы размещаетесь в гостинице, оставляете вещи в комнате, похожей как две капли воды на комнату в любом другом месте, где вы уже побывали на конференции в этом году, и направляетесь на регистрацию. Если дело происходит в США, то скорее всего штаб-квартира конференции будет тут же, в отеле, а если в континентальной Европе - то в специально выстроенном дворце конгрессов; если же год неудачный и конференция проходит в Великобритании - то в простом, захудалом университетском здании, а вместо похожего на все другие отели вас ждет обветшалое студенческое общежитие. Во всех случаях придется выстоять очередь, чтобы приблизиться к нескольким откуда-то принесенным и сдвинутым вместе столам, за которым сидят озабоченные секретарши или специально набранные студенты; они просмотрят свои бумаги, чтобы выявить, не являетесь ли вы приглашенным докладчиком и уплатили ли регистрационный взнос (при особо солидном мероприятии на их месте могут оказаться профессиональные организаторы, облаченные в аккуратную униформу и напоминающие служащих конторы по прокату автомобилей). Наконец, вам вручают пластмассовый портфельчик - пожертвование, какой-нибудь фармацевтической компании или местного инициативного комитета.
Теперь разумнее всего присесть где-нибудь неподалеку и разгрузить этот портфельчик от неизбежных дарственных экземпляров ненужных вам журналов и рекламных брошюр, оставив только программу конференции и сборник ее материалов. Большинство из нас слишком скаредны, чтобы заодно избавиться и от блокнота с шариковой ручкой, на которой красуется название биохимической компании, и мы оставляем их про запас, а потом привозим домой, где в конце концов выбрасываем или теряем. Среди подобных предметов мы находим также ярлычок со своей фамилией и стопку конвертов с билетами или раззолоченными приглашениями на приемы, обеды и частные встречи, без которых не обходятся такие съезды. Первым в списке встреч неизбежно стоит вечер по случаю открытия конференции, где ее участники знакомятся друг с другом. Он вот-вот начнется. Поэтому вы мчитесь в отель, бросаете все ненужное, распаковываете зубную щетку (позднее, перед сном, вам будет не до нее), прицепляете ярлычок к лацкану пиджака и отправляетесь на вечер с выпивкой (но не надевайте свежую рубашку: она понадобится вам завтра и, кроме того, требования к вашему костюму на таких вечерах не выше качества кушеток, на которых вам придется сидеть).
Ваше мрачное настроение с примесью беспокойства готово теперь превратиться в откровенно панический страх перед неизвестностью, но постарайтесь подавить эти чувства, ибо вы становитесь участником одного из важнейших событий научного мероприятия. Проще всего было бы относиться к этому как к академическому туризму, который Эшли Монтегю как-то назвал досугом ученого сообщества и который служит фоном дюжины романов из университетской жизни. Но все обстоит сложнее. Уже через несколько минут после появления в изукрашенном по-старинному зале городской ратуши, в танцевальном зале отеля или у бортика бассейна вы уже не бродите рассеянно в поисках бокала посредственного вина и не присматриваетесь к значкам с неразборчивыми надписями на пиджаках участников в надежде найти хотя бы смутно знакомое вам имя, а ведете доверительный разговор с людьми, которых встречали на предыдущих конгрессах, что уже дает вам право считать их старыми друзьями. Конференция для вас уже началась, и то, что происходит во время таких приемов и вечеров, составляет такой же элемент научного общения, как и доклады о новейших результатах в аудитории.