
Роуз С. Р79 Устройство памяти. От молекул к сознанию: Пер с англ
| Вид материала | Решение |
СодержаниеИстоки поведения Одиночные клетки Многоклеточные организмы |
- «хм «Триада», 9393.37kb.
- Анастази А. А 64 Дифференциальная психология. Индивидуальные и групповые разли- чия, 11288.93kb.
- Роджер Л. М2Э вирус ответственности.: Пер с англ, 2943.44kb.
- Новые поступления литературы (июль сентябрь 2002) математика инв. 62350 в 161., 125.41kb.
- Указатель произведений литературы зарубежных стран (библиотека кф ат и со), 250.17kb.
- Литература для клинических интернов по специальности «терапия» Кафедра факультетской, 55.33kb.
- Куртц П. К93 Искушение потусторонним: пер с англ, 7904.74kb.
- The guilford press, 6075kb.
- The guilford press, 6075.4kb.
- Жизнь счастливого человека (А. Маслоу), 76.37kb.
Истоки поведения
Адаптивное поведение как обязательное условие выживания свойственно всем организмам, однако многие процветающие формы жизни прекрасно обходятся без научения и вспоминания, даже без мозга. Возьмем подсолнечник, соцветие которого всегда обращено к источнику света, т. е. энергии. По мере роста подсолнух не учится на опыте, как лучше поворачиваться, и он не перестанет делать это, если после каждого поворота «наказывать» его электрическим ударом. Это поведение (поворачивание соцветия), носит оно адаптивный характер или нет, Дано организму от природы, зафиксировано в его генетическом коде и программе развития. Такое присущее виду поведение (обычно говорят врожденное, но я избегаю пользоваться этим термином, так как он несет дополнительную идеологическую нагрузку) гарантирует адекватный ответ на специфические раздражители без научения методом проб и ошибок, но за счет ограничения диапазона и гибкости реакций. Поведение изменяется не только у отдельной особи, но и в длинном ряду поколений как следствие эволюции. В отличие от этого приобретенные формы поведения вырабатываются на опыте, на практике; их преимущества — большая гибкость, свобода корректировки при изменениях окружающей среды и возможность учитывать наблюдаемые последствия при жизни данного индивидуума. Очень важно, однако, помнить, что приобретенные путем научения реакции не наследуются потомством. Передаются лишь степень пластичности и способность обучаться, которые в свою очередь «заданы» генетическим кодом и программой развития. Вот почему пора забыть наскучившие старые споры о врожденном и приобретенном. Чтобы познать мозг и поведение, нужно отрешиться от этого противопоставления и попытаться вместо того понять диалектическую связь между специфичностью и пластичностью.
Способ существования растений определяется их способностью выживать, используя энергию Солнца для усвоения двуокиси углерода и преобразования ее в нужные питательные вещества, в частности в сахар. Для этого надо лишь оставаться на месте с раскинутыми ветвями, осуществлять фотосинтез и стараться избегнуть поедания, чтобы успеть произвести потомство. За это приходится расплачиваться большой сложностью способов размножения, которые по самой своей сути не очень надежны, так как зависят от других организмов или природных сил (например, ветра), разносящих пыльцу и семена. Жизнь животных не столь проста. Не обладая способностью к фотосинтезу, они вынуждены использовать частично готовую пищу, либо непосредственно поедая растения, либо охотясь на животных, питающихся растениями. Для этого им приходится непрерывно передвигаться, прибегать к специализированным способам поиска корма, спасаться от опасностей и разыскивать партнеров. Таким образом, неспособность к фотосинтезу компенсируется расширением репертуара адаптивного поведения и его большей гибкостью, развитием органов чувств, позволяющих обнаруживать пищу и врагов на расстоянии, и двигательными навыками для перемещения в желаемых направлениях.
Одиночные клетки
Некоторые из описанных форм поведения встречаются даже у простейших одноклеточных организмов. Еще в 80-х годах прошлого столетия в Германии В. Пфейфер провел следующий эксперимент. Он опускал тонкую капиллярную трубочку, заполненную раствором глюкозы, в каплю жидкости, содержавшей бактерии, и наблюдал, как они концентрировались у отверстия капилляра. Бактерии вели себя так, будто «знали», где находится глюкоза, и демонстрировали адаптивную реакцию. Объясняется это довольно просто: глюкоза медленно диффундировала из капилляра в окружающую жидкость, и здесь создавался градиент концентрации сахара. В мембранах бактериальных клеток имеются рецепторные молекулы — белки, строение которых позволяет им узнавать молекулы глюкозы, и крошечные бичевидные жгутики бактерий, работая синхронно, перемещают клетку «вверх» по градиенту. У мутантных форм, лишенных рецепторов, такое поведение отсутствует, что не мешает им использовать глюкозу как источник энергии, если их поместить в раствор.
Более сложные одноклеточные организмы, например парамеции, имеют выросты иного типа, называемые ресничками. Основания ресничек соединены системой тонких нитей, координирующих их биение. В интервалах между эпизодами питания (добычей им служат бактерии) парамеции находятся в непрестанном движении и часто наталкиваются на препятствия, после чего меняют направление движения, заставляя реснички работать на той или другой стороне тела. Таким образом эти животные, перемещаясь подобно игрушечным машинкам, обходят препятствия и достигают участков, богатых пищей. Точно так же они способны уплывать из зон со слишком высокой или низкой температурой или избегать таких раздражающих веществ, как кислоты. Эти реакции поиска и избегания у существ размером не более 0,2 мм, не имеющих нервной системы, есть не что иное, как зачатки более сложных форм активного поведения.
Многоклеточные организмы
Одноклеточные организмы должны быть автономными системами и вмещать в своем крошечном объеме все, что необходимо для их поведенческого репертуара. Зато у них нет особых трудностей со связью между отдельными частями клетки. В процессе эволюции многоклеточных организмов спектр поведения расширяется, и им приходится решать все более сложные организационные проблемы. Возникает надобность в какой-то системе быстрой внутренней сигнализации, чтобы координировать деятельность клеток в различных частях одного организма. Главным средством межклеточной связи у наиболее примитивных многоклеточных форм, по-видимому, служила химическая сигнализация. Вещество, выделяемое одной клеткой и служащее, например, сигналом к сокращению, могло достаточно быстро диффундировать в другие клетки, заставляя их тоже сокращаться. Такие химические сигналы были возможными предшественниками нынешних гормонов. Один из самых интригующих аспектов биохимической эволюции — близкое сходство многих гормонов с веществами, которые служат в нервной системе медиаторами; оно может означать, что последние ведут свое происхождение от гормонов.
Многоклеточность делает возможной специализацию. Отдельные свойства и фукнции могут распределяться между различными клетками: одни клетки выполняют, например, сократительную функцию, другие специализируются на синтезе и секреции сигнальных веществ, в третьих, расположенных на поверхности тела, сосредоточены рецепторы, реагирующие на химические воздействия (подобно рецепторам глюкозы у бактерий) или даже на свет.
Система сигнализации путем диффузии различных веществ удобна для мелких организмов, но ее возможности ограниченны: диффузия на большие расстояния занимает много времени и малоэффективна, так как сигнал не может быть направлен настолько точно, чтобы достигнуть именно той клетки, для которой он предназначен. Если же, с другой стороны, сигнализирующая клетка приобретает форму, которая позволяет ей прийти в соприкосновение с клеткой-мишенью, то химический сигнал может быть непосредственно передан через «синаптическую» щель между клетками. Это обеспечивает направленность, но не решает проблему передачи сигнала с одного конца генерирующей его клетки на другой, которые теперь разделены довольно большим расстоянием.
В этом случае большое значение приобретают электрические свойства клетки. Наличие электрического заряда на наружных клеточных мембранах — всеобщая особенность живых клеток. Такой заряд, создающий мембранный потенциал, обусловлен присутствием во внутриклеточной жидкости (цитоплазме) различных растворенных солей, образующих электрически заряженные ионы — Na+, К+, Са2+, Сl- и т. д. (например, из хлористого натрия, NaCl, образуются положительно заряженные ионы натрия, Na+, и отрицательно заряженные ионы хлора, Сl-). Однако внутриклеточная среда отличается от наружной среды более высокой концентрацией калия и более низкой концентрацией натрия. В клетках содержатся также белки, а входящие в их состав аминокислоты тоже несут электрические заряды. Таким образом, ионный состав жидкостей по обе стороны клеточной мембраны различен, и в результае на мембране создается разность потенциалов порядка 70 милливольт с отрицательным полюсом внутри (рис. 7.1).

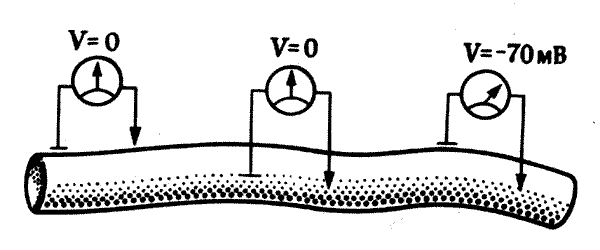
Рис. 7.1. Мембранный потенциал покоя. Если на поверхность нервного волокна (или любой другой клетки) наложены два регистрирующих электрода, разность потенциалов между ними отсутствует. То же самое наблюдается, если оба электрода введены внутрь волокна. Но если один из них находится на поверхности, а другой внутри, то внутри волокна регистрируется потенциал около -70 милливольт по отношению к поверхности. Это мембранный потенциал покоя; он обусловлен биохимическими и физическими свойствами мембраны, которые приводят к различиям в концентрациях натрия, калия и других ионов по обе ее стороны.
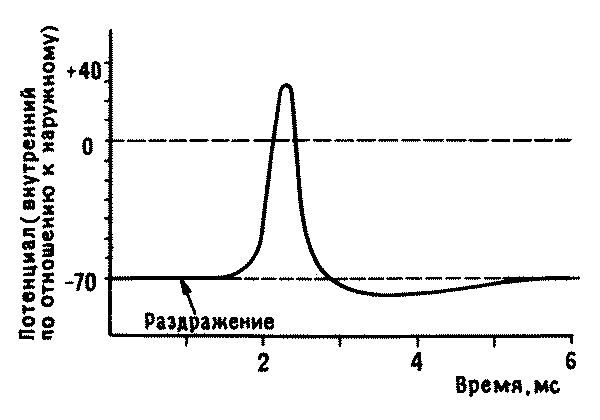
Рис. 7.2. Потенциал действия. Способность генерировать потенциал действия — уникальное свойство возбудимых клеток, таких, как нейроны. Если аксон нервной клетки подвергнуть электрической, механической или химической стимуляции, свойства мембраны в месте раздражения быстро изменяются. Из наружной среды в аксон входят ионы натрия, и это приводит к быстрому скачку потенциала от -70 до + 40 милливольт; затем поступление ионов натрия внутрь прекращается, и они выводятся наружу, пока не установится первоначальная разность потенциалов. Этот кратковременный сдвиг потенциала виден на приведенной кривой; он распространяется вдоль по аксону подобно волне, которая обычно возникает в месте отхождения аксона от тела клетки и доходит до выходного синапса, где вызывает освобождение нейромедиатора; медиатор диффундируют через синаптическую щель и таким образом передает постинаптической клетке нервный сигнал.
Нервные клетки (нейроны), как и все другие, обладают таким мембранным «потенциалом покоя». Но они отличаются уникальным свойством своей мембраны, которая у них возбудима, т. е. быстро становится проницаемой для ионов натрия, находящихся во внеклеточной среде, в ответ на соответствующий сигнал — на небольшое местное изменение ионных концентраций по обе ее стороны. Поступление в клетку ионов натрия приводит к деполяризации мембраны, потенциал на которой изменяется с минус 70 милливольт до плюс 40 милливольт. Это генерирует в клеточной мембране волну электрической активности, называемую потенциалом действия (рис. 7.2), которая за несколько миллисекунд распространяется по аксону от тела клетки до ее выходного синапса. Потенциал действия в свою очередь служит сигналом для высвобождения в синаптическую щель нейромедиатора, вызывающего реакцию другого нейрона. Эволюция клеток, обладающих потенциалами действия и системами химической сигнализации на своих окончаниях, возможно, лежала в основе формирования нервной системы современного типа.
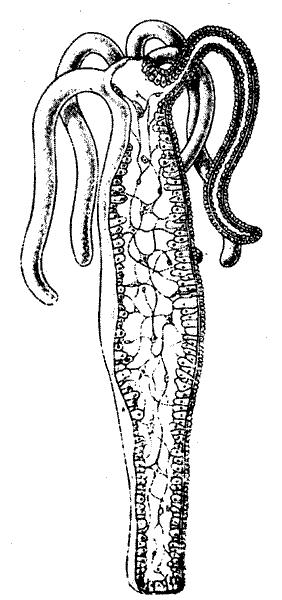
Рис. 7.3. Гидра. Обратите внимание на диффузную нервную сеть во всем теле. ионы натрия, Na+ и отрицательно заряженные ионы хлора, Сl-). Однако внутриклеточная среда отличается от наружной среды более высокой концентрацией калия и более низкой концентрацией натрия. В клетках содержатся также белки, а входящие в их состав аминокислоты тоже несут электрические заряды. Таким образом, ионный состав жидкостей по обе стороны клеточной мембраны различен, и в результае на мембране создается разность потенциалов порядка 70 милливольт с отрицательным полюсом внутри (рис. 7.1).
Примером организма с примитивной нервной системой может служить крошечная гидра, живущая в наших водоемах (рис. 7.3). Гидры прикрепляются к камням или водным растениям на дне пруда или ручья, и их щупальца совершают волнообразные движения около ротового отверстия. В случае прикосновения к нему животное, подобно актинии, сжимается в комочек. Гидры питаются мельчайшими организмами, проплывающими мимо щупалец, причем предварительно выбрасывают специальные ядовитые нити для обездвиживания добычи, которая затем проталкивается щупальцами в рот. Это сложное поведение требует механизмов для обнаружения добычи или опасности, принятия решения и соответствующей реакции — атаки или сжатия в шарообразный комок. Такими механизмами служат чувствительные, секреторные и мышечные клетки и прежде всего вся сеть поверхностных клеток, объединенная электрической сигнализацией и способная координировать поведение гидры.
Отдельные клетки этой сети несколько отличаются от нейронов более сложно организованных животных, так как функции сети лишены специфичности или направленности. Если подвергнуть раздражению любую часть тела, то от этого места во все стороны пойдет волна возбуждения, которая в конце концов охватит всю нервную сеть. Нервная система гидры напоминает такую систему телефонной связи, в которой вас рано или поздно соединят со всеми другими абонентами, какой бы номер вы ни набрали. Напротив, важнейшей чертой высокоразвитой нервной системы является специфичность, наличие определенного набора соединений, благодаря которым сигнал, возникающий в какой-либо сенсорной клетке, проходит строго определенный путь и достигает определенной эффектор ной клетки: это нечто вроде «частной» линии связи, в основном изолированной от множества других нейронов нервной системы.