
Роуз С. Р79 Устройство памяти. От молекул к сознанию: Пер с англ
| Вид материала | Решение |
СодержаниеКритерии соответствия Первый критерий Второй критерий |
- «хм «Триада», 9393.37kb.
- Анастази А. А 64 Дифференциальная психология. Индивидуальные и групповые разли- чия, 11288.93kb.
- Роджер Л. М2Э вирус ответственности.: Пер с англ, 2943.44kb.
- Новые поступления литературы (июль сентябрь 2002) математика инв. 62350 в 161., 125.41kb.
- Указатель произведений литературы зарубежных стран (библиотека кф ат и со), 250.17kb.
- Литература для клинических интернов по специальности «терапия» Кафедра факультетской, 55.33kb.
- Куртц П. К93 Искушение потусторонним: пер с англ, 7904.74kb.
- The guilford press, 6075kb.
- The guilford press, 6075.4kb.
- Жизнь счастливого человека (А. Маслоу), 76.37kb.
Критерии соответствия
К 1980 году исследователи памяти уже достаточно хорошо разбирались, чему можно верить, а чему нет. Стало, например, ясно, что не существует уникальных молекул памяти. Сложилось общее мнение, что наиболее перспективна модель Доналда Хебба, в которой предполагалось изменение эффективности связей между нервными клетками, вероятно, в результате образования новых или роста существующих синапсов, и этим объяснялась перестройка функциональных отношений между нейронами. Модификации хеббовского типа, по-видимому, можно было бы выявлять нейрофизиологическими или биохимическими методами. Однако ни у кого не было уверенности, что какой-либо специфический биохимический процесс, кроме самого общего процесса белкового синтеза, можно однозначно связать с формированием следов памяти; ученые стали проявлять большую осторожность при оценке новых сообщений на эту тему. Что же такое в конце концов биохимия памяти и как проверить правильность предлагаемых ответов на этот вопрос? Иными словами, какой ответ был бы убедительным и для биохимиков, и для психологов? Каким требованиям должен отвечать эксперимент, чтобы изучаемый процесс можно было считать необходимым, достаточным и, быть может, даже специфичным для образования следов памяти?
На этот вопрос я попытался ответить в начале восьмидесятых годов [1]. К тому времени уже не составляло труда обнаружить биохимические изменения в мозгу обучавшегося животного; трудность заключалась в доказательстве того, что они действительно связаны с процессом запоминания. Я чувствовал, что не могу разумно планировать эксперименты, не имея критериев для оценки их результатов, а без этого я не знал бы, как двигаться дальше. Я решил, что любое изменение, которое можно будет рассматривать как часть следа памяти (энграммы), должно соответствовать критериям, описанным ниже1.

*1) Я несколько изменил формулировку этих критериев и порядок их обсуждения по сравнению с тем, как они были представлены в оригинальной статье, но это не затрагивает их сути. Я не хочу переписывать историю.
Первый критерий
Предполагаемый фактор (система, вещество, скорость его образования или обновления) в локализованном участке мозга должен претерпевать количественные изменения в процессе формирования следов памяти.
Если образуются новые или видоизменяются существующие синапсы, то в их химических или структурных компонентах должны появляться изменения, доступные для выявления биохимическими или микроскопическими методами (например, повышенное содержание синаптических мембранных белков, изменение размеров определенных синапсов или увеличение их числа). Однако если бы я выявил такое изменение в период обучения животного, но не убедился в его соответствии всем последующим критериям, я бы недалеко ушел от экспериментов шестидесятых годов, которые критиковал в предыдущей главе. То или иное изменение может быть действительно необходимым, но это еще не значит, что оно достаточно или специфично. Тем не менее первый критерий, безусловно, имеет фундаментальное значение для любой материалистической модели памяти. Он предполагает также условие локализации, т. е. изменения не могут быть распределены по всему мозгу - они должны быть сосредоточены в какой-то определенной области. Это, разумеется, возвращает нас к старому спору о возможности локализации энграмм (к этой теме я вернусь позднее; значительная часть двух последних глав будет посвящена вопросу о локализации памяти во времени и пространстве).
Обратите внимание, что первый критерий не включает таких условий, как направление и величина изменений. Вполне возможно представить себе «отрицательное» кодирование воспоминаний путем снижения уровня какого-то вещества или процесса, хотя на практике почти цсе исследователи пытаются выявить его повышение. А что же с величиной изменения?
Если что-то запомнилось вдвое лучше или вместо одного события или предмета запомнились два, значит ли это, что и изменение выражено вдвое сильнее? Совсем не обязательно, ибо в наших опытах мы измеряем память в условных единицах, которые сами же изобрели. При этом мы не можем знать, соответствуют ли наши масштабы тем, которыми пользуются сами животные, формируя следы памяти. Например, я могу обучить цыпленка не клевать горькую хромированную бусину, потом горькую красную и, наконец, горькую синюю бусину. Однако цыпленок не обязательно помнит их как три отдельных предмета. Скорее всего он вместо этого прибегает к иной, более рациональной стратегии, производя обобщение: «Все предметы определенного размера, независимо от цвета, имеют горький вкус, и их следует избегать». В этом случае он будет помнить один предмет, а не три.
Второй критерий
Изменение должно совпадать во времени с формированием следов памяти.
Очевидно, что следы образуются не мгновенно, словно нажатием кнопки, а на протяжении нескольких часов после запоминаемого события. В этот период изменяется форма сохранения следа. По крайней мере в случае декларативной памяти (глава 4) имеет место переход, занимающий несколько минут или часов, от первоначальной лабильной и кратковременной фазы к долговременной стабильной памяти.
Можно представить себе разные способы такого перехода (рис. 9.1). Одним из крайних вариантов может быть непрерывный процесс, в котором некая цепь биохимических реакций в определенной группе клеток неизбежно приводит от ранней, неустойчивой фазы к фиксированному конечному состоянию, как при затвердевании клея или проявлении фотоснимка. Другой вариант допускает возможность двух (или нескольких) более или менее независимых параллельных процессов, сопровождающихся изменениями электрических свойств и чувствительности какой-то группы нейронов, которые могут в первые минуты «кодировать» память, а затем постепенно отключаться. Между тем, если воспоминание достаточно важно, чтобы стоило сохранять его долгое время, могут потребоваться такие биохимические процессы, как реконструкция синапсов, которые будут постоянно представлять память в другой группе нейронов, возможно даже в другой части мозга. Эти два типа процессов - последовательная и параллельная обработка воспоминаний - служат, разумеется, крайними примерами; есть множество промежуточных вариантов [2], и становится все труднее планировать эксперименты, в которых можно было бы четко провести различие между ними.
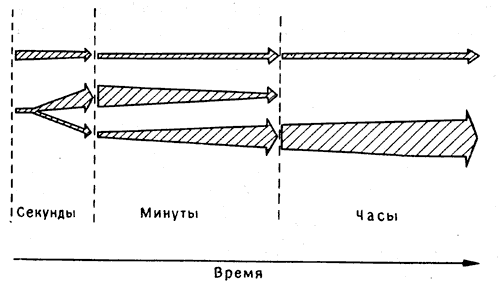
Рис. 9.1. Кратковременная и долговременная память. Две модели перехода от кратковременной памяти к долговременной. Вверху показан вариант с последовательным, линейным переходом от фазы к фазе. Внизу - вариант с двумя независимыми процессами. Первоначальная активация клеток служит сигналом как для запуска механизмов кратковременного запоминания, так и для образования и последующего укрепления долговременных энграмм. При блокировании второго из этих процессов воспоминание утрачивается по мере угасания кратковременных следов. Имеющиеся данные говорят скорее в пользу нижнего варианта.
Тем не менее весь опыт изучения человеческой памяти указывает именно на такое подразделение памяти на кратковременную и долговременную. Результаты исследований, проведенных на таких больных, как X. М., показывают, что гиппокамп, не участвуя в проявлениях долговременной памяти, все же играет важную роль в переходе от кратковременной к долговременной фиксации ее следов. Но как бы ни работал мозг - путем последовательной или параллельной обработки информации (в дальнейших главах этому будет уделено больше внимания) - в обоих случаях с памятью связаны определенные клеточные процессы, и мы не можем не пытаться дифференцировать их экспериментальным путем.