
Лекция Структура нуклеиновых кислот. Роль молекул ДНК и рнк в клетках
| Вид материала | Лекция |
- Спецкурс «Химия нуклеиновых кислот и основы генной инженерии» для студентов 4 курса, 23.84kb.
- План состав нуклеиновых кислот Состав ДНК, 232.1kb.
- Урок 7 Тема: нуклеиновые кислоты, 118.42kb.
- «Нуклеиновые кислоты», 177.07kb.
- Тема нуклеиновые кислоты, 160.68kb.
- Лекция Нуклеиновые кислоты. Атф нуклеиновые кислоты, 119.07kb.
- Рабочей программы учебной дисциплины молекулярная биология уровень основной образовательной, 42.15kb.
- Нуклеиновые кислоты, 122.08kb.
- Нуклеиновые кислоты, 296.24kb.
- Функции в организме, 166.53kb.
Лекция 1.
Структура нуклеиновых кислот. Роль молекул ДНК и РНК в клетках
Нуклеиновые кислоты – это линейные полимеры, мономерами которых являются нуклеотиды. Полимеры – это макромолекулы, которые состоят из большого числа повторяющихся структурных единиц – мономеров. Мономерами ДНК являются дезоксирибонуклеотды, мономерами РНК – рибонуклеотиды.
Строение и номенклатура нуклеотдов. В состав нуклеотида входят три компонента: фосфат – сахар – основание.
углеводный компонент нуклеотида представлен рибозой или 2’-дезоксирибозой, имеющих D-конфигурацию.

D-рибоза 2’-дезокси- D-рибоза
В отличие от D-рибозы 2’-дезокси- D-рибоза не содержит атом кислорода при втором углеродном атоме. Атомы углерода в пентозе нумеруются со знаком штрих (‘) – 1’, 2’, 3’, 4’, 5’.
Азотистые основания – это гетероциклические органические соединения, содержащие атомы азота. В составе ДНК встречаются 4 типа оснований - аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т), в состав РНК входят А, Г, Ц и У (урацил). Аденин и гуанин являются производными пурина, цитозин, тимин и урацил – это производные пиримидина.
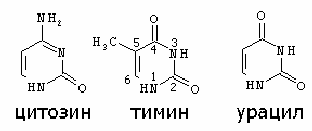
Пиримидиновые основания
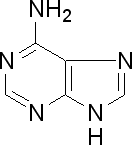
Аденин
Пуриновые основания
Номенклатура. Соединение, состоящее из основания и углевода, называется нуклеозидом. Азотистые основания соединяются с 1’ углеродным атомом пентозы β-гликозидной связью.
Аденозин, гуанозин, цитидин и уридин являются нуклеозидами рибозы. Дезоксиаденозин, дезоксигуанозин, дезоксицитидин, дезокситимидин являются дезоксинуклеозидами аденина, гуанина, цитозина и тимина, соответственно.
В свободном состоянии нуклеотиды находятся в виде 5’-трифосфатов: АТФ, ГТФ, ЦТФ и УТФ для рибозы и дАТФ, дГТФ, дЦТФ и ТТФ для дезоксирибозы.
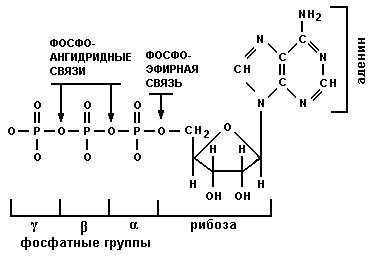
Рис. Структура аденозинтрифосфата (АТФ).
В молекулах ДНК нуклеотиды присутствуют в виде 5’-монофосфатов: дАМФ, дГМФ, дЦМФ и ТМФ. Монофосфаты РНК обозначают: АМФ, ГМФ, УМФ, ЦМФ. В мононуклеотидах фосфатная группа соединяет простой эфирной связью с 5’-углеродным атомом пентозы.
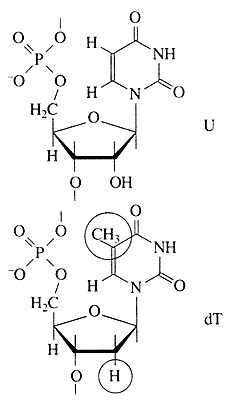
Рис. Структура уридилового (U) и тимидилового (dT) нуклеотидов (кружками отмечены отличия в структуре тимина и урацила, рибозы и дезоксирибозы).
Первичная структура полимера определяется последовательностью мономеров в цепи. Нуклеотиды соединяются друг с другом 3’,5’-фосфодиэфирной связью, образуя полинуклеотидные цепи из сотен тысяч и миллионов нуклеотидов. Короткие цепочки из десяти – пятнадцати нуклеотидов называются олигонуклеотидами. Фосфат связывает 3’-ОН группу одного нуклеотида с 5’-OH группой другого нуклеотида.
Первичная структура ДНК (РНК) определяется последовательностью нуклеотидов в цепи. Эта структура стабилизируется (поддерживается) 3’,5’-фосфодиэфирными ковалентными связями.
5’- конец полинуклеотидной цепи

3’-конец полинуклеотидной цепи
Рис. Первичная структура нуклеиновых кислот. Фрагмент полинуклеотидной цепи из четырех нуклеотидов. R – Азотистое основание, X – (ОН -) или Н- группа для рибозы или дезоксирибозы, соответственно.
Вторичная структура ДНК. Молекула ДНК в клетках прокариот и эукариот присутствует только в виде двойной спирали, т.е. состоит из двух полинуклеотидных цепей. Эти цепи комплементарны, антипараллельны и закручены в спираль вокруг общей оси. На один виток спирали приходится 10 пар оснований, диаметр спирали составляет 2 нм. Сахарофосфатный остов расположен снаружи (заряжен отрицательно), азотистые основания находятся внутри спирали и располагаются стопкой друг над другом. Эта модель строения ДНК была предложена Дж. Уотсоном и Ф. Криком в 1953 году. Основой для создания модели двойной спирали послужили результаты рентгеноструктурного и химического анализа молекул ДНК.
Правила Чаргаффа. В 1953 году с помощью кислотного гидролиза ДНК с последующей хроматографией и количественным анализом Эрвин Чаргафф установил следующие закономерности:
- количество пуриновых оснований (A+Г) в молекуле ДНК всегда равно количеству пиримидиновых оснований (Т+Ц).
- количество аденина равно количеству тимина [А=Т, А/Т= 1]; количество гуанина равно количеству цитозина [Г=Ц, Г/Ц=1];
- соотношение количества гуанина и цитозина в ДНК к количеству аденина и тимина является постоянным для каждого вида живых организмов: [(Г+Ц)/(А+Т)=К, где К - коэффициент специфичности].
Правила Чаргаффа, как правило, выполняются на двойной спирали ДНК за счет комплементарности аденина тимину, а гуанина - цитозину. В некоторых случаях содержание гуанина выше, чем цитозина, за счет метилирования некоторых цитозиновых остатков в ДНК.
Принцип комплементарности. Азотистые основания в молекуле ДНК могут образовывать канонические пары: А – Т, Г – Ц. это значит, что водородные связи и молекуле ДНК образуются только между комплеменатрными основаниями: между аденином и тимином образуется две, между гуанином и цитозином – три водородные связи. Эти пары называют уотсон-криковскими парами по имени ученых, открывших модель структуры ДНК.
Цепи ДНК антипараллельны. Каждая цепь ДНК имеет два конца – 5’- конец и 3’- конец. На 5’- конце полинуклеотидной цепи 5-ОН группа дезоксирибозы не связана с другим нуклеотидом, на другом конце цепи 3-ОН группа тоже не связана с другим нуклеотидом. Правило антипараллельности означает, что две цепи в молекуле ДНК имеют противоположную направленность. За направление цепи по соглашению принято направление 5’ → 3’.
Правила написания последовательности ДНК: в виде последовательности букв, обозначающих основания: 5’ – GATCCA - 3’, или в виде стрелок с противоположной ориентацией:
5’ 3’
 5’
5’3’
Связи, стабилизирующие вторичную структуру ДНК:
- Водородные связи между комплементарными азотистыми основаниями
- Стэкинг-взаимодействиями, которые возникают между плоскими азотистыми основаниями, расположенными стопкой друг над другом в двойной спирали ДНК.
Физические параметры ДНК. Типы двойных спиралей ДНК
В-форма ДНК – основной тип двойной спирали. В двойной спирали на один виток приходится 10 пар нуклеотидов (п.н.), длина витка – 3,4 нм. Расстояние между соседними нуклеотидами составляет 0,34 нм, диаметр спирали – 2,0 нм.
А-форма ДНК образуется при дегидратации. Характеризуется более широкой и короткой спиралью, основания имеют более сильный наклон к оси спирали (+19). На один виток спирали приходится 11 пар оснований. Расстояние между соседними нуклеотидами составляет 0,23 нм, длина витка – 2,5 нм, диаметр спирали – 2,3 нм. А-форму имеет спираль РНК-ДНК в комплексе матрица-затравка, а также спираль РНК-РНК и шпилечные структуры РНК (2’-гидроксильная группа рибозы не позволяет молекулам РНК образовывать В-форму). А-форма ДНК обнаружена в спорах. Установлено, что А-форма ДНК в 10 раз устойчивее к действию УФ-лучей, чем В-форма.
С-форма ДНК имеет структуру, сходную с В-ДНК. Число пар оснований на виток составляет 9,33, длина витка спирали равна 3,1 нм. Пары оснований наклонены на угол 8 градусов относительно перпендикулярного положения к оси. Желобки по размерам близки к желобкам В-ДНК. При этом главный желобок несколько мельче, а минорный желобок – глубже. В С-форму могут переходить природные и синтетические полинуклеотиды ДНК.
Z-форма ДНК – это двойная спираль, закрученная влево. На один виток этой формы приходится 12 пар оснований. Расстояние между соседними нуклеотидами составляет 0,38 нм, длина витка – 4,56 нм, диаметр Z-ДНК – 1,8 нм. Z- форма ДНК обнаружена в клетках прокариот и эукариот. Обычно Z-форма образуется в участках ДНК, где пурины чередуются с пиримидинами (например, 5’-ГЦГЦГЦ-3’), или в повторах 5’-ЦГЦГЦГ-3’, содержащих метилированный цитозин.
В клетках ДНК обычно существует в В-форме, которая является самой стабильной, но отдельные участки ДНК могут находиться в А-форме, Z-форме или даже в другой форме. Некоторые участки молекулы ДНК со специфическими последовательностями образуют изгибы, что имеет важное значение для регуляции экспрессии генов.
Типы двойных спиралей ДНК.
| Характеристика параметры спирали | Тип двойной спирали | |||
| А | В | Z | C | |
| Форма молекулы | широкая | Средняя ширина | Самая узкая | |
| Расстояние между соседними нуклеотидами | 0,23нм | 0,34нм | 0,38нм | 0,33нм |
Диаметр спирали | 25,5А | 23,7А | 18,4А | |
| Число пар оснований на виток спирали | 11 | 10,4 | 12 | 9,33 |
| Наклон оснований относительно оси спирали | 19 | 1 | 9 | 8 |
Большая бороздка | Узкая и очень глубокая | Широкая и глубокая | плоская | Широкая и мелкая |
Малая бороздка | Очень широкая и мелкая | Узкая и глубокая | Очень узкая и глубокая | Очень глубокая, узкая |
| | | | | |
Форма и размеры молекул ДНК. В клетках прокариот и эукариот молекула ДНК всегда состоит из двух цепей (дуплекс) и может иметь кольцевую или линейную форму. Геномы ДНК-содержащих вирусов могут быть представлены как одноцепочечной или двухцепочечной молекулой ДНК (кольцевой или линейной).
Кольцевая суперспирализованная двухцепочечная молекула ДНК представляет хромосомы бактерий, плазмиды, геномы митохондрий, пластид и некоторых вирусов.
Линейная двухцепочечная ДНК находится в хромосомах эукариот и некоторых вирусов.
Размеры молекул ДНК. За единицу длины молекулы ДНК обычно принимают тысячу пар нуклеотидов (т.п.н.), длину небольших фрагментов ДНК (олигонуклеотидов) измеряют в парах нуклеотидов (п.н.). Например, геном кишечной палочки E.coli представлен кольцевой молекулой ДНК размером 4х106 п.н., а ДНК вируса SV40 составляет всего 5243 п.н. Геном человека составляет 3,8х109 п.н.
Физические свойства молекулы ДНК. При повышении температуры или добавлении щелочей происходит разрыв водородных связей между комплементарными основаниям. При этом происходит денатурация (плавление) и нативная двухцепочечная ДНК переходит в одноцепочечную форму. При охлаждении или понижении рН среды происходит ренатурация (отжиг), т.е. восстановление двойной спирали ДНК.
Температура плавления (Тпл.), при которой денатурирует 50% всей ДНК, зависит от содержания Г=Ц пар в молекуле ДНК. У млекопитающих, в том числе и у человека, Тпл. ДНК составляет 870С. Способность ДНК денатурировать и восстанавливать двойную структуру используют для гибридизации при образовании дуплексов ДНК-РНК или ДНК-ДНК.
Структура и функции РНК.
Содержание РНК в 5 – 10 раз превышает содержание ДНК в клетках (кроме сперматозоидов).
Типы молекул РНК.
- Во всех клетках присутствуют тРНК, рРНК, мРНК
- Большинство клеток содержит большое количество малых цитоплазматических РНК (мцРНК)
- В клетках эукариот присутствуют малые ядерные РНК (мяРНК).
Основные функции молекул РНК:
- Участие в реализации генетической информации.
- Принимают участие в выполнении специфических эндонуклеазных реакций, регулирующих этапы экспрессии генов.
- Генетический материал некоторых вирусов (ретровирусов и реовирусов)
Относительное содержание в клетках молекул РНК
| Тип РНК | Количество видов молекул | % содержание массы клеточных РНК |
| мРНК | <1000 | 5% |
| тРНК | 100 | 15% |
| рРНК | 3 - 4 | 80% |
| мяРНК | | 2% |
| мцРНК | неизвестно | |
Компоненты РНК, химические связи в молекуле.
Мономерами РНК являются рибонуклеотиды. В состав рибонуклеотида входит рибоза, гетероциклическое основание и остаток фосфорной кислоты.
Обычно в состав РНК входит 2 пурина (аденин и гуанин) и 2 пиримидина – цитозин, а вместо тимина в состав РНК входит урацил, у которого отсутствует 5-метильная группа.
Модифицированные пурины и пиримидины в РНК содержат метил-, тиол-, водород и изопентил- заместители. 2’-O-метилнуклеотиды с модифицированными остатками рибозы, а также наблюдается иной способ связывания урацила с рибозой (псевдоуридиловая кислота). Модифицированные пурины и пиримидины часто встречаются в тРНК и никогда не присутствуют в иРНК.
Как правило, модификация оснований и рибозных остатков происходит после завершения синтеза РНК.
Первичная структура РНК определяется последовательностью рибонуклеотидов в цепи. Нуклеотиды в молекуле РНК соединяются посредством 3’,5’ - фосфодиэфирных связей, образуя полинуклеотидные цепи. Клеточные молекулы РНК состоят из одной полинуклеотидной цепи, геномы некоторых вирусов представлены двухцепочечными молекулами РНК, напоминающими А-форму ДНК. Наличие 2’-OH группы в рибозе определяет чувствительность фосфодиэфирной связи в молекуле РНК к действию щелочей и ферментов РНКаз, расщепляющих РНК.
Вторичная структура РНК. В большинстве молекул тРНК и рРНК имеются небольшие участки комплементарных последовательностей, которые спариваются и образуются петли. Эти области двухцепочечных участков РНК непротяженные и прерывисты. Комплементарные пары оснований в двухцепочечных участках РНК:
А=У, Г≡Ц, Г=У (менее стабильная, 2 водородные связи)
Структура клеверного листа. Зависимость функции молекулы от ее упаковки показана на примере тРНК. Компьютерные модели показали, что все тРНК имеют вторичную структуру типа «клеверный лист», которая имеет 4 стебля (акцепторный, D, антикодоновый и T) и три петли (D-петля, антикодоновая и T-петля). Спаренные основания образуют стебли, не спаренные основания - петли.
5’- и 3’-концы тРНК образуют акцепторный стебель. 3’-конец тРНК содержит три не спаренных нуклеотида ЦЦА. Псевдоуридиновая петля содержит модифицированные азотистые основания псевдоуридин (ψ) и риботимидин (Т). Антикодоновая петля находится напротив акцепторного участка и содержит антикодон (три нуклеотида, комплементарных кодону мРНК). Еще одна петля содержит дигидроуридин (DU). Вариабельная петля находится между антикодоном и псевдоуридиновой петлей и содержит разное число нуклеотидов.
Третичная структура тРНК. Рентгеноструктурный анализ показал, что эта структура более компактная, чем клеверный лист. Она образуется благодаря взаимодействиям, сближающим дигидроурациловую (DU) и псевдоуридиновые (Т ψ) петли. Такой общий вид молекулы получил название L-конфигурации, которая указывает на наличие двух взаимно перпендикулярных участков молекулы тРНК – в одной части находится акцепторный участок, в другой – антикодон.
Денатурация и ренатурация РНК. При повышении температуры двухцепочечные участки РНК разрушаются, но в отличие от молекул ДНК, при высоких значениях рН (действии щелочей) в РНК разрушаются фосфодиэфирные связи. После денатурации восстановление двуцепочечных участков РНК затруднено и в результате образуются структуры, отличные от исходной молекулы РНК.
Лекция 2.
Биосинтез ДНК. Молекулярные основы передачи генетической информации
Полуконсервативный механизм репликации ДНК. Перед каждым делением клетки в ней должно удвоиться содержание ДНК, чтобы каждая дочерняя клетка получила полный набор хромосом. Основу каждой хромосомы образует одна двухцепочечная молекула ДНК. Предложенная Дж. Уотсоном и Ф. Криком модель строения ДНК форме регулярной двойной спирали сразу же позволила понять принцип копирования ДНК. Ее репликация происходит полуконсервативным способом: две исходные цепи материнской ДНК расходятся, и каждая из них становится матрицей для синтеза новой комплементарной цепи. Таким образом, каждая новая двойная спираль ДНК содержит одну старую и одну новую цепь. Такой механизм репликации ДНК, при котором от одного поколения к другому передается одна из двух материнский цепей ДНК, получил название полуконсервативного и был экспериментально доказан в 1958 году М. Мезельсон и Ф. Сталь.
Легко представить, что удвоение ДНК происходит вследствие того, что цепи расходятся и каждая цепь служит матрицей для синтеза новой комплементарной цепи ДНК. Каждая дочерняя молекула состоит из одной старой материнской и одной новой синтезированной цепи ДНК.
Общие принципы репликации ДНК.
В основе процесса репликации лежит принцип копирования материнской цепи ДНК с образованием двух идентичных молекул ДНК. В основе синтеза новой цепи ДНК лежит принцип комплементарности азотистых оснований, т.е. последовательность нуклеотидов материнской цепи определяет последовательность нуклеотидов в синтезируемой цепи ДНК
Синтез новых цепей ДНК идет в направлении 5’ → 3’.
В основе репликации ДНК лежат принципы антипараллельности и униполярности. Синтез новых цепей ДНК идет в направлении от 5’-конца к 3’ - концу, при этом ДНК-полимеразы – ферменты, синтезирующие новые цепи ДНК, - добавляют нуклеотиды к 3’- концу наращиваемой цепи ДНК. При этом матричная цепь имеет противоположную ориентацию: 3’ → 5’, т.е. ДНК-полимеразы могут передвигаться вдоль матрицы только в направлении от 3’ к 5’-концам.
3’ 5’ матрица ДНК
5
 ’ 3’ направление синтеза новой цепи ДНК
’ 3’ направление синтеза новой цепи ДНКРис. Схема расположения матричной и дочерней цепи ДНК.
Реакция синтеза полинуклеотидной цепи ДНК.
Механизмы синтеза ДНК хорошо изучены в клетках бактерий, однако имеются доказательства, что в клетках эукариот процесс протекает аналогичным образом.
Инициация репликации ДНК. Репликация начинается в специфическом участке молекулы ДНК, который называется точка начала репликации или ориджин.
Точка начала репликации (origin) – это участок молекулы ДНК со специфической последовательностью нуклеотидов с большим содержанием пар АТ (последовательность 300 п.н., богата АТ). Специальные инициирующие белки необходимы для связывания ферментов репликации с молекулой ДНК:
- белок DnaA – для прокариот
- белок RPA (replication protein A) – для эукриот.
Кольцевая хромосома прокариот имеет одну точку начала репликации, которая называется OriC. В этой точке цепи расходятся и образуются две репликативные вилки, которые движутся в противоположном направлении. Скорость синтеза ДНК в клетках прокариот составляет 500 нукл./сек. Две вилки встречаются на противоположной стороне кольца. В клетках прокариот существует специальный фермент гираза (топоизомераза II), который разделяет две образующиеся кольцевые молекулы ДНК. Антибиотик налидиксовая кислота угнетает размножение бактерий, путем инактивации гиразы. В клетках эукариот этот фермент отсутствует, поэтому налидиксовую кислоту используют в клинической практике для лечения бактериальных инфекций.
Репликация ДНК эукариот начинается одновременно во многих точках начала репликации, от каждой точки движутся две репликативные вилки в противоположных направлениях. Скорость синтеза молекул ДНК эукариот составляет 50 нукл./сек.
Репликон – фрагмент молекулы ДНК, репликация которого происходит под контролем одной точки начала репликации. Кольцевая хромосома прокариот имеет 1 репликон. Геномы эукариот содержат сотни и тысячи репликонов.
Инициация - образование репликативной вилки. Нити ДНК разделяются благодаря действию специальных ферментов и белков.
Хеликаза (от англ. helix – спираль) - основной фермент, расплетающий цепи ДНК. У прокариот он называется белок DnaB. Хеликаза разрывает водородные связи между комплементарными основаниями, используя энергию АТФ.
Топоизомеразы – ферменты, которые устраняют положительные сверхвитки перед репликативной вилкой. Эти ферменты временно разрывают нити материнской ДНК в двойной спирали перед репликативной вилкой, после завершения процесса нити ДНК восстанавливают целостность.
SSB - белки – это белки, которые связываются с одноцепочечной ДНК и удерживают матрицу. В результате образуется репликативная вилка, где и происходит синтез новых цепей ДНК.
Механизм действия ДНК-полимераз. Ферменты, катализирующие процесс синтеза новых цепей ДНК называются ДНК-зависисмые ДНК-полимеразы. В клетках бактерий существует три типа ДНК-полимераз: ДНК-полимераза I, II, III.
Синтез ДНК в репликативной вилке катализирует ДНК-полимераза III. ДНК-полимераза I играет важную роль в синтезе отстающей цепи и репарации ДНК. ДНК-полимераза II участвует в репарации ДНК.
Реакция синтеза. ДНК-полимеразы могут только удлинять (элонгировать) уже существующую полинуклеотидную нить, которую называют затравкой или праймером. В клетках роль затравки играет олигонуклеотид РНК (РНК-праймер), который комплементарен матрице и образует с ней двухспиральный комплекс матрица-затравка. Синтез РНК праймера осуществляет фермент праймаза. Затем ДНК-полимераза синтезирует цепь ДНК, используя 3’-OH конец праймера. После окончания синтеза ДНК РНК-праймеры удаляются с помощью ферментов – экзонуклеаз.
ДНК-полимераза имеет 2 субстрата:
Комплекс матрица – затравка и
- дезоксинуклеозидтрифосфаты (АТФ, ГТФ,ЦТФ и ТТФ)
Реакция осуществляется как удлинение цепи с 3’- конца затравки
Необходимы ионы Mg 2+ (3’ – OH) конец полинуклеотидной цепи атакует α-фосфат дНТФ, образуя фосфодиэфирную связь
Матрица копируется точно на основе принципа комплементарности
Кроме полимеризующей активности, ДНК-полимеразы обладают корректирующей активностью: они дважды проверяют нуклеотид, который добавляют в растущую цепь ДНК. Корректирующая активность ДНК-полимераз проявляется в способности отщеплять нуклеотид, некомплементарный матрице, и замещать его комплеметарным нуклеотидом.
Асимметричность репликативной вилки. Поскольку ДНК-полимераза наращивает цепь ДНК в направлении 5’ → 3’, синтез дочерних цепей идет в противоположном направлении. Одна цепь синтезируется непрерывно (лидирующая), а другая синтезируется прерывисто, в виде фрагментов Оказаки (отстающая). Фрагменты Оказаки – это короткие цепочки ДНК вместе с РНК-праймерами, расположенные на отстающей цепи.
Синтез отстающей цепи. ДНК-полимераза III останавливается перед РНК-праймером предшествующего фрагмента Оказаки. Здесь подключается ДНК-полимераза I, которая удаляет РНК-праймеры предшествующего фрагмента Оказаки и одновременно наращивает 3’-OH – конец растущей цепи ДНК, заполняя образующуюся брешь. Фермент лигаза соединяет два фрагмента Оказаки, используя АТФ. Таким образом, на отстающей цепи также синтезируется непрерывная цепь ДНК.
Функции теломеразы.
Проблема недорепликации 3’-концов линейных молекул ДНК заключается в том, что удаление последних РНК-праймеров с 5’-концов обеих цепей дочерней ДНК, приводит к тому, что дочерние цепи оказываются короче материнской на 10-20 нуклеотидов (у организмов разных видов размер РНК-праймеров различен). Проблема недорепликации 3’-концов линейных молекул ДНК эукариот решается с помощью специального фермента – теломеразы.
В случае репликации кольцевых молекул ДНК этой проблемы не существует, т.к. первые РНК-затравки удаляются ДНК-полимеразой I, которая одновременно наращивает 3’-OH – конец растущей цепи ДНК и заполняет образующуюся брешь.
Ограничение числа клеточных делений. Лимит Хейфлика.
Американский ученый Л. Хейфлик в начале 60-х годов 20 века показал, что культивируемые клетки новорожденных детей могут пройти 80-90 делений, в то время как соматические клетки 70-летних людей делятся 20-30 раз. Такое явление ограничения числа клеточных делений называют “лимитом Хейфлика”.
В 1971 г. А.М. Оловников в своей теории маргинотомии (от лат. marginalis – краевой, toma - сечение) предположил, что последовательное укорочение ДНК хромосом во время репликации лежит в основе ограниченного числа делений, которое наблюдается у нормальных соматических клеток, растущих в культуре in vitro. Позже был обнаружен фермент теломераза, активный в стволовых, половых и раковых клетках, которые способны к бесконечному делению.
Функции теломеразы.
Фермент теломераза впервые был обнаружен в 1985 году у равноресничной инфузории Tetrahymena thermophila, а позже – в клетках дрожжей, растений и животных, в том числе в яичниках человека и иммортализованных (бессмертных) линиях раковых клеток HeLa.
Согласно номенклатуре этот фермент называют теломерной терминальной трансферазой. Теломераза выполняет функции ДНК-полимеразы, которая достраивает 3’-концы линейных молекул ДНК короткими повторяющимися последовательностями нуклеотидов (у позвоночных – ТТАГГГ) в отсутствии ДНК матрицы. В качестве матрицы для синтеза цепи ДНК теломераза использует молекулу РНК, которая является составной частью этого фермента.
Строение теломеразы. Теломераза, кроме белковой части, содержит молекулу РНК, которая исполняет роль матрицы для наращивания 3’-конца ДНК-матрицы. Молекула теломеразной РНК содержит от 150 нуклеотидов у простейших до 1400 нуклеотидов у дрожжей, у человека она содержит 450 нуклеотидов. Поскольку РНК является матрицей для синтеза ДНК, теломеразу можно отнести к обратной транскриптазе, т.е. ферменту, синтезирующему ДНК на матрице РНК.