
Лекция №11. Кооперативные процессы в эволюции Мы попытаемся разобраться, как могла возникнуть многоклеточность, как многоклеточные потом усложнились
| Вид материала | Лекция |
- Лекция Тип Кишечнополостные (Coelenterata) Подцарство Многоклеточные, 97.31kb.
- Пётр Кропоткин взаимопомощь как фактор эволюции, 899.4kb.
- Взаимопомощь как фактор эволюции, 871.42kb.
- Взаимопомощь как фактор эволюции, 938.72kb.
- Лекция 6-2009 по экологической геохимии 17. 03., 133.18kb.
- Самоконтроль диабета, 809.21kb.
- Доклад Начало и развитие отношениям по договору долевого участия в строительстве было, 72.11kb.
- Лекция №12. Процессы кооперации в социальной эволюции Вспомним схему, которую мы уже, 187.08kb.
- Лекция 1 Введение. Общие сведения об операционных системах, 196.75kb.
- Вряд ли найдется другой современный режиссер, о котором сказано, сколько о Бруке,, 11196.1kb.
Лекция №11. Кооперативные процессы в эволюции |
Мы попытаемся разобраться, как могла возникнуть многоклеточность, как многоклеточные потом усложнились.
Вспомним, каких одноклеточных мы знаем. Об одноклеточных мы уже говорили на предыдущих лекциях. Амеба - простейшее одноклеточное, относящаяся к типу Protozoa. Клетка у амебы выполняет все жизненно необходимые для нее функции. На рисунке показано, как она охотится. Она выпячивает ложноножки, которые охватывают нечто съедобное для амебы.

На обзорной лекции по зоологии было рассказано о том, как образуются колонии. Ниже представлена колония вольвокса (одноклеточные со жгутиками). В принципе, клетка не погибает, если отделить ее от колонии, но при размножении довольно быстро образуется колониальная форма. Колония способна размножаться, то есть внутри колонии образуются более мелкие колонии. До 10 000 одноклеточных может входить в такие колонии. Действия между ними согласуются через контакты между отдельными клетками, то есть все клетки машут жгутиками согласованно, чтобы колония могла перемещаться не беспорядочно.

Более сложно устроено примитивное животное губка. Хотя с виду губки похожи на растения, к перемещению не способны, они отностятся к животным, потому что фотосинтезом не занимаются и имеют животный тип питания. Стенка у губки имеет трехслойное строение: покровные (толстые наружные) клетки, жгутиковые клетки (хоаноциты, внутренние клетки). Пространство между наружным и внутренним слоями клеток заполнено слизистым веществом, в котором находятся клетки амебоциты (похожи на амебу). В отличие от кишечнополостных, во внутренней полости у губок не происходит пищеварение. Эта полость служит только для протока воды. Жгутики машут таким образом, что вода протекает через поры в теле губки и выходит через устье губки. С водой поступают частицы, которые хоаноциты захватывают и поглощают. Покровные клетки и амебоциты питаются частью съеденной хоаноцитами пищи, которую те им передают. С одной стороны, мы видим дифференцированные клетки, но с другой стороны, они дифференцированы не насовсем: хоаноцит может превратиться в амебоцит, потом перебраться на другую сторону и стать покровной клеткой. Таким образом, нельзя сказать, что губки имеют ткани, как у высших животных: хотя у них и имеются слои дифференцируемых клеток, последние специализированы временно. Еще раз подчеркнем, что тут уже имеется разделение функций между клетками, клетки не одинаковые, но это разделение временно.

Каким образом клетки, содержащие одинаковый геном, могут иметь разную форму и выполняют разные функции? Для этого должны синтезироваться разные белки, которые идут и на строительство клеток, и на ферментативные функции. Гены во всех клетках одинаковые, за исключением половых клеток. То есть, гены во всех клетках одинаковые, но при этом клетки имеют разную форму и разные функции. Это объясняется тем, что в каждой клетке работают не все гены, а только те, которые нужны в данный момент. Гены могут включаться и выключаться, то есть, как говорят, активироваться либо быть репрессированными (выключенными).
Сделаем небольшое отступление, чтобы обсудить регуляцию активности генов. Обсудим мы ее на примере бактерий. У них регуляция активности генов несколько проще, чем у высших организмов, но принцип тот же. Мы уже говорили о переключении работы групп генов при помощи изменения сродства РНК-полимеразы к промотору. Напомним. РНК-полимераза – это фермент, который считывает генетическую информацию с ДНК и синтезирует матричную РНК. Как вы помните, в РНК-полимеразе есть σ-фактор – белок, который распознает промотор и помогает РНК-полимеразе на него сесть и начать транскрипцию. Таким образом переключается работа больших групп генов, это такая системная регуляция. Клетка переключается с одной жизненной программы на другую. Сегодня будет рассказано о еще двух системах регуляции. Они называются катаболитная активация (или катаболитная репрессия), и активация-репрессия отдельных оперонов. Оказалось, что у бактерий группы генов, которые кодируют совместно работающие белки, расположены рядом. Ниже приведен пример лактозного оперона (гены, кодирующие белки, которые перерабатывают лактозу). Первый фермент, кодируемый геном lacZ, необходим для расщепления лактозы при извлечении энергии из сахара лактозы; ген lacY кодирует фермент пермеазу, который переносит лактозу из внешней среды внутрь клетки; третий ген также кодирует фермент, метаболизирующий лактозу. Эти три гена находятся рядом друг с другом, и считываются с одного промотора. Гены, транскрибируемые с одного промотора, называются опероном. В данном случае мы говорим о лактозном опероне. Концепция оперона была предложена Жакобом, Моно и Вольманом, за что они получили Нобелевскую премию.
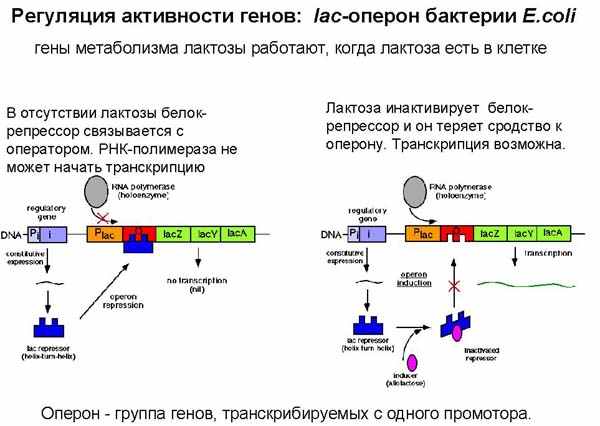
Каким же образом регулируется работа этого оперона? Для того, чтобы он работал, необходимо, чтобы РНК-полимераза распознала промотор и начала синтез матричной РНК. При этом в ней будут считаны сразу три гена, и с полученнрой мРНК будут синтезированы три отдельных белка. Выгодно было бы для кишечной палочки включать работу лактозного оперона, когда лактоза есть, и выключать, когда лактозы нет. Эта система работает следующим образом. После промотора, до начала структурных генов (гены, кодирующие белки), находится участок, который называется оператор. На нем в отсутствии лактозы находится белок, называемый белком-репрессором лактозного оперона. Он кодируется отдельным геном, находящимся рядом с лактозным опероном и постоянно работающим. С этого гена синтезируется своя мРНК, с нее транслируется белок-репрессор, и этот репрессор садится на операторный участок. Когда здесь находится белок-репрессор, РНК-полимераза не может сесть на промотор и начать синтез. Белок-репрессор физически не дает ей этого сделать. Если в среде появляется лактоза, она связывается с белком-репрессором, тот меняет свою конфигурацию и отваливается от оператора. РНК-полимераза может начать свою работу и считать структурные гены. То есть, в присутствии лактозы синтез мРНК лактозного оперона разрешен, в ее отсутствие –запрещен, репрессирован.
В присутствии глюкозы для E. coli не выгодно использовать другие сахара - лактозу, галактозу, мальтозу (период генерации при росте на глюкозе - 50 мин, на лактозе - 80 мин). При использовании лактозы необходимы некоторые дополнительные реакции, переводящие ее в форму, которую клетка может использовать.
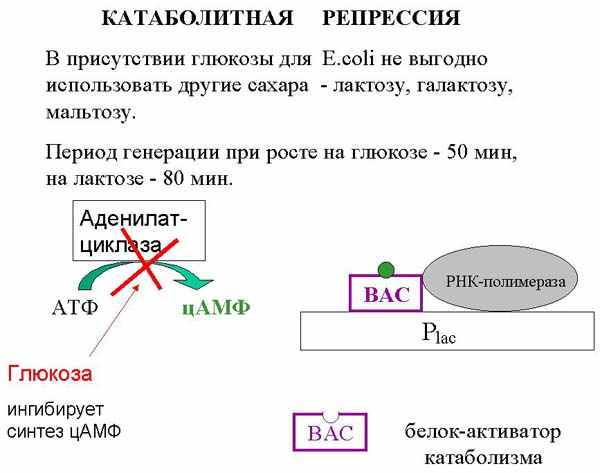
Значит, если во внешней среде имеются одновременно и лактоза, и глюкоза, то запускать обработку лактозы невыгодно; нужно использовать более выгодную для клетки глюкозу. Существует несколько возможных алгоритмов того, как это сделать, некоторые из которых реализуются в других оперонах. Для лактозного оперона природа придумала такое решение. У кишечной палочки есть белок, который вместо того, чтобы репрессировать этот оперон, его активирует. Этот белок называется белок-активатор катаболитных генов (БАК, английская аббревиатура BAC). Дело в том, что лактозный промотор сам по себе не настолько сильный, чтобы с него шла хорошая транскрипция. БАК садится перед промотором и помогает РНК-полимеразе начать транскрипцию. Без этого белка РНК-полимераза не способна запустить транскрипцию с лактозного промотора. А сесть перед промотором белок может только тогда, когда в клетке нет глюкозы.
Как устроена эта система. Дело в том, что БАК способен связаться с ДНК только тогда, когда белку присоединен циклический аденазинмонофосфат (цАМФ). Это вещество образуется из АТФ специальным ферментом аденилатциклазой. В присутствии глюкозы аденилатциклаза блокируется и концентрация цАМФ в клетке падает, и цАМФ больше не может связываться с БАК. БАК диссоциирует с промотра, транскрипция прекращается. Когда глюкоза попала в клетку, она ингибирует работу аденилатциклазы, белок слезает не только с лактозного, но и с некоторых других промоторов, в частности, прекращается метаболизм мальтозы и некоторых других сахаров прекращается, потому что глюкоза – наиболее выгодный продукт.
Таким образом, лактозный белок-репрессор специфичен: у него только одно любимое место во всем геноме кишечной палочки – оператор лактозного оперона, в то время как БАК-белок может сесть на несколько промоторов. Не на все, но на строго заданные - на те, которые не должны работать в присутствии глюкозы. В отсутствие глюкозы он сидит на промоторах и активирует их, а как только глюкоза в клетку попадает, транскрипция с этих промоторов прекращается.

Ниже представлена шариковая модель циклического АМФ. Он – типичный регулятор внутриклеточного метаболизма. Такая система изменения активности аденилатциклазы и, соответственно, концентрации циклического АМФ в клетке, работают не только у бактерий, но и у очень многих организмов, в том числе и у нас с вами. Через аденилатциклазу, регулируя ее активность, действуют некоторые гормоны. Меняя концентрацию циклического АМФ, эти гормоны влияют на внутриклеточные процессы.

Интересно, что циклический АМФ используется не только для регуляции внутриклеточных процессов, но и для межклеточной коммуникации при формировании многоклеточности. Об этом будет рассказано на примере уникального организма – амебы, которая называется Dictyostelium discoideum. Это одноклеточная амеба, которая живет в почве и питается бактериями.
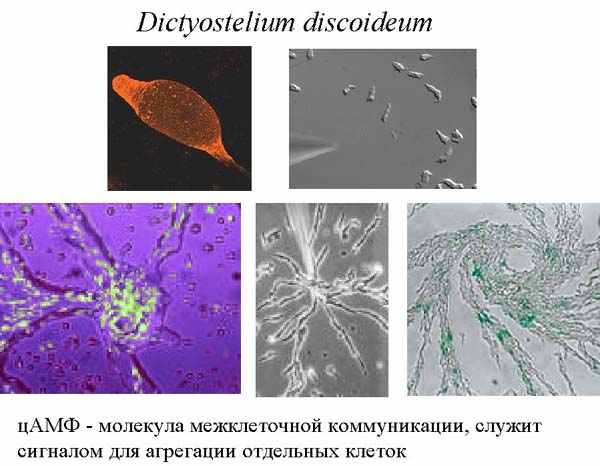
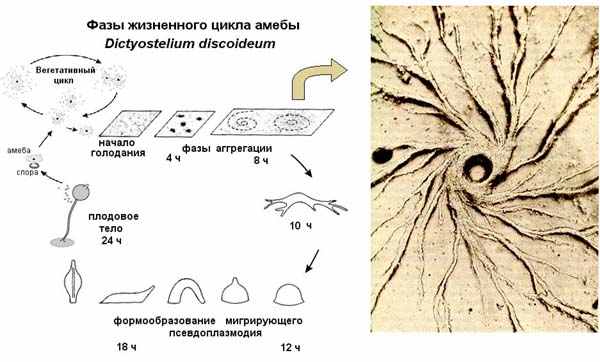
Когда все хорошо, амебы диктиостелиума ползают по своему месту обитания, питаются и делятся время от времени. Но если они голодны, долго не попадалось хорошей еды и их энергозапасы начинают истощаться, они выпускают во внешнюю среду цАМФ, соседние клетки этот сигнал воспринимают. Если клетки сыты, то они на него не реагируют, если же они голодны, то они начинают сползаться в кучу. Вначале они собираются небольшими группами, выпускают циклический АМФ, его концентрация становится больше, поэтому одиночные клетки начинают к ним подползать, образуя агрегат клеток. В итоге к самой большой кучке сползаются остальные группы клеток, и они формируют единый многоклеточный организм, который называется псевдоплазмодий. Он способен ползать, и в отличие от амебы, способен перемещаться на заметные расстояния. Он с довольно заметной скоростью уползает из плохого места. Если ему удается переползти туда, где есть еда, то он опять распадается на отдельные клетки, которые, как приличные одноклеточные амебы, начинает питаться. Если же он ползет – ползет, а хорошая жизнь все не наступает, то он останавливается, примерно 20% клеток ползут вверх, образуя жесткий стебелек, и затем отмирают (то есть, приносят себя в жертву всем остальным). По стебельку остальные клетки переползают на самую верхушку, образуют плодовое тело, в котором созревают споры. Они разбрасываются на некоторое расстояние вокруг, пережидают неблагоприятный период. Когда наступает хорошее время, они прорастают в амебы, и вся история начинается заново. Этот процесс проиллюстрирован на картинке ниже. Эта амеба не является ни одноклеточным, ни многоклеточным организмов. В ее случае мы сталкиваемся с тем, что то, что можно наблюдать в живой природе, сложнее, чем придуманная людьми система классификации. Dictyostelium часть своей жизни одноклеточный, другую часть – многоклеточный, и он способен переходить из одной формы в другую, то есть, так просто его не классифицируешь. Он на стадии псевдоплазмодия имеет дифференцированные клетки. Его передний конец обладает хеморецепцией, он лучше все чувствует, чем задний конец псевдоплазмодия. И образование плодового тела – это процесс уже ярко выраженной дифференциации, при которой 20 % клеток образует стебелек и погибают. Размер генома Dictyostelium’a 3.5x107 bp. Мы можем назвать Dictyostelium факультативно многоклеточным.

Итак, благодаря тому, что не все гены работают в клетке одновременно, клетка может менять программу своей активности, образовывать разные ферменты и иметь разную форму. Это происходит как у одноклеточных, так и у многоклеточных. У многоклеточных регуляция еще более сложная, так как помимо внутриклеточных процессов нужно регулировать еще межклеточные взаимодействия. Но возникает вопрос, как вообще могли образоваться многоклеточные. Как исходно одноклеточные организмы превратились в многоклеточные? У практически всех одноклеточных известны мутанты, не расходящиеся при делении клетки. То есть нормальное деление клетки происходит, все у нее делится: ядра делятся, митохондрии делятся, хлоропласты расходятся по разным дочерним клеткам, но последний этап, когда клетки должны отцепиться друг от друга, у них не происходит, они остаются сцепленными. У некоторых видов эти клетки-мутанты живут хуже, чем нормальные одноклеточные формы, потому что у них нет системы регуляции взаимодействия клеток. Но некоторые виды, видимо, приспособились к этому, эти клетки начинают взаимодействовать друг с другом, у них есть программа согласования своих действий. Нерасхождение при делении встречается не только на уровне одноклеточных, но и на уровне многоклеточных, (например, сиамские близнецы). Не всегда это является уродством, иногда это вариант нормы.
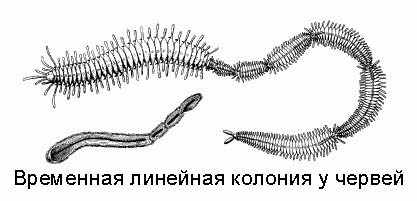
На рисунке вы видите червей, которые после деления не разошлись. Все вместе они образуют то, что называется временной линейной колонией. Если их друг от друга отделить, они будут дальше вполне комфортно жить. Такая колония может потом сама разделиться. Но происходит это не сразу, какое-то время черви живут все вместе.
Существуют колонии так называемых гидроидных полипов. Напомним, что гидра относится к типу Кишечнополостных, подцарство Metazoa. У гидры следующее строение: имеется внешний слой клеток - эктодерма, внутренний слой клеток – эндодерма, (эти клетки могут заниматься пищеварением), есть щупальца, способные захватывать жертву, на которых у нее есть стрекательные клетки, есть рот, внутри гидры есть полость, в которой происходит пищеварение. Гидра прикрепляется ко дну водоема при помощи так называемого базального диска. У гидры имеется сеть из нервных клеток, большое скопление этих клеток находится у щупалец, так как надо реагировать мгновенно на любое прикосновение. Такое строение имеют и остальные представители кишечнополостных.

Многие виды кишечнополостных типа образуют колонии. Ниже представлена колония животных – гидроидных полипов, которая с виду похожа на растение. Каждый "листочек" такого "растения" имеет такое же строение, как гидра.

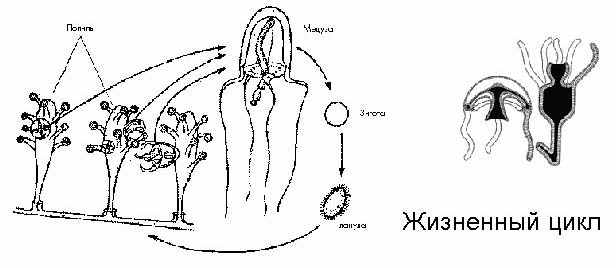
Жизненный цикл у некоторых из гидроидных полипов проходит следующим образом: полипы растут вместе, они объединены общим проводящим каналом, время от времени некоторые из них отрываются, превращаются в медуз (принципиальное строение гидры и медузы одинаковое), которые уплывают и активно размножаются, проходят определенные стадии, образуют половые клетки, которые, сливаясь, дают зиготу, из которой вырастает новый полип. И снова весь цикл повторяется.
Известные вам красивые кораллы – это скелет коралловых полипов. Полипы – похожие на гидру существа, у который есть общий проток воды по общим объединяющим их каналам, они строят для себя известковый скелет. У некоторых этот скелет включает в себя соединения железа, которые окрашивают его в красивый красный цвет. Коралловые полипы живут в море. На следующем рисунке представлены разные формы коралловых полипов.

На следующем рисунке слева представлен мадрепоровый коралл. У него отдельные полипы уже неразличимы, они все сливаются в единое целое, от них остаются только глотки с щупальцами, причем эти щупальца обобществлены, они заглатывают кусочки пищи и направляют ее в общую полость. Справа виден родственник гидры и полипа – мшанка кристателла. У нее тоже отдельные особи сливаются своими основаниями. Каждая особь называется не полипом, а зооидом, то есть это как бы и не отдельное животное, и не орган. Они способны втягиваться в трубку у основания общего тела. Объединенная часть их образует подошву, и, в отличие от кораллов, мшанка способна передвигаться со скоростью 15 мм в сутки.

Другой более отдаленный родственник гидры - физалия - относится к подклассу сифонофор. Это животное называют португальский военный кораблик. У физалии есть сверху парус, который может иметь разную окраску, от голубого до пурпурного. Она довольно ядовитая, у нее есть стрекательные клетки, которые способны сильно отравить даже человека, не говоря уже о мелких животных, которыми она питается. На рисунке представлена схема строения сифонофор. Тело сифонофоры устроено из группы зооидов, разным обарзом модифицированных.
Самый верхний образует плавательный пузырь, внутри него находятся клетки, выделяющие газ, если физалии надо подняться на поверхность. Под плавательным пузырем находятся плавательные колокола – это как бы отдельные индивиды, но здесь они уже превращаются в органы. Ниже находятся индивиды, которые используются как половые органы, они не способны питаться, поэтому другие зооиды – гастрозоиды, их кормят, но не только их, но и весть «организм». Есть защитные зооиды, имеющие стрекательные клетки.
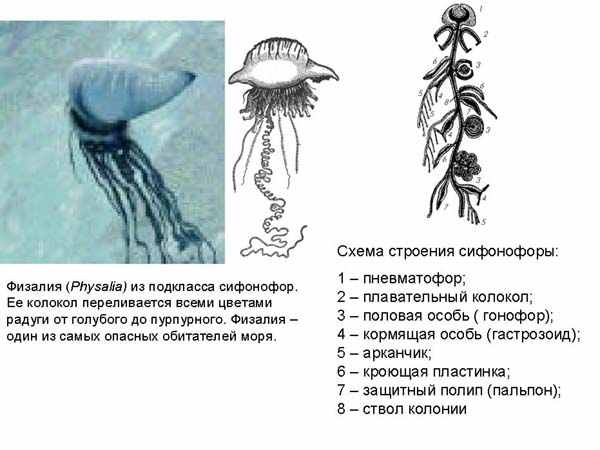
Таким образом, если у колонии коралловых полипов каждый полип равнозначен, то здесь колония может превращаться в достаточно сложноустроенный организм, с дифференцированными полипами, каждый из которых превращается в орган. Таким образом этот организм нельзя однозначно проклассифицировать – является ли он колоний организмов или отдельным организмом.
Аналогичную ситуацию мы можем наблюдать, изучая социальных насекомых – муравьев, пчел, термитов.
У пчел (так же как и у муравьев и термитов) обычно имеется единственная плодоносящая матка. Остальные пчелы – рабочие самки, они неплодоносящие и по сравнению с маткой мелкие. Они кормят матку, которая ничем другим не занимается, кроме как откладывает яйца, из которых появляются рабочие пчелы или трутни.
На картинке показаны термиты.

Они строят огромные термитники. Справа сверху показаны: 2 рабочие особи, солдат и матка. Пока она функционирует, другие особи не могут развиться в матку. Если ее убрать, то одна из рабочих особей превращается в царицу – плодовитую матку. Это регулируется (так же как и у пчел и муравьев) через выделение ферамонов и др. веществ матки. То есть матка выделяет вещества, которые ингибируют (запрещают) трансформацию других самок в матку. Когда матка умирает, все самки начинают выделять эти вещества, и та самка, которая опережает других, тормозит их, становясь маткой. Это означает, что у самок не утеряна способность плодоносить, но она репрессирована.
У термитов, если есть вторая плодоносящая самка, то она улетает в другое место и образует новое сообщество. Одна оплодотворенная матка может создать целое гнездо. Она хранит спермии в определенном резервуаре, которые расходует для оплодотворения яиц очень экономно. После того, как она отложит яйца, она за ними начинает ухаживать, как только появляются личинки, матка их кормит частью яиц, и после этого, личинки превращаются в рабочих особей. Они начинают помогать матке кормить новых личинок вначале яйцами, затем добычей. Часть из рабочих особей ходят за добычей, а часть строят термитник. Таким образом от одной плодоносящей матки может получиться новое гнездо с огромным количеством особей.
У пчел матка может откладывать как оплодотворенные, так и неоплодотворенные яйца. Из первых образуются рабочие пчелы-самки, из вторых – трутни. Трутни могут вылетать в другие ульи и оплодотворять чужую матку, таким образом осуществляя перекрестное оплодотворение. Хотя можно использовать трутней из своего улья, потому что при этом все равно происходит перекомбинация генетического материала.
Здесь следует подчеркнуть, что вегетативное деление клетки вполне обеспечивает увеличение числа клеток. Половой процесс нужен для перекомбинации генетического материала. Есть виды, например среди одноклеточных, для которых описано самооплодотворение. Когда заканчивается обычное деление, проходит половой процесс. К делится, образуя половые клетки, которые потом сами с собой сливаются, то есть потомки одной клетки сливаются друг с другом и образуют зиготу. В данном случае деление не приводит к увеличению числа потомков, но у них перемешивается генетический материал.
Какую можно провести аналогию между термитами и, например, с сифонофорами? Мы здесь видим отдельных индивидов, вроде бы независимых друг от друга, но они образуют колонию, хотя в отличие от сифонофор, отдельные части этой колонии способны самостоятельно передвигаться. Но ясно, что колония – это «организм» с дифференцированными органами. Этими органами являются отдельные термиты. Есть репродуктивный орган – матка, орган защиты – солдаты, есть органы питания и ухода за личинками – рабочие самки, есть система коммуникаций (химическая и поведенческая формы) между органами. Как же социальные насекомые между собой общаются?
В свое время было сенсацией расшифровка языка пчел.

Когда пчела нашла нечто съедобное, она прилетает обратно в свой улей и сообщает остальным, что и где она нашла. Что она нашла, понятно по тому, что прилипло к ее телу. А где она это нашла – сообщает на языке танца. Во время танца пчела движется по вертикальной поверхности. Она совершает движение, схематически отражающее направление и расстояние до места, куда следует лететь за едой.
Солнце в этой схеме по умолчанию находится сверху. Например, если место кормежки расположено прямо по Солнцу, то пчела ползет вверх и делает такую «восьмерку», как показано на рисунке. Таким образом направление она указала. Если место для питания под углом 600 , то пчела поворачивает восьмерку на 600 (рисунок b). Аналогично остальные ситуации. А количеством виляний брюшком она указывает на расстояние. У разных пчел разные диалекты, поэтому у них одно число виляний брюшком показывает разное расстояние. Также пчела может объяснить не только, как лететь по прямой к месту кормежки, но и как облететь, например, гору. Если еда находится рядом с ульем, то пчела выполяет танец, который показан на рисунке слева вверху.
Как же пчела ориентируется на местности, как запоминает ориентиры? Дело в том, что пчелы видят все в поляризованном свете, у них по другому устроено зрительное восприятие, и есть посвященные этому исследования. Интересно знать, что у пчел врожденное владение языком. При жизни они, конечно, совершенствуют свой танец, добавляют новые, свойственные только этому улью элементы танца, но в принципе они изначально могут показать путь к месту, где есть еда. Если взять пчел, говорящих на разных диалектах, то оказывается, они не могут понять друг друга. Возникает вопрос: применяют ли пчелы еще в каких-нибудь ситуациях свой «язык»? Нет. Для остальных ситуаций у пчел хорошо развита система коммуникаций, регулирующая отношения индивида внутри роя, для того, чтобы они делали то, что надо. Но наиболее загадочным и интригующим для ученых была их способность рассказать о том, куда лететь. За работу над расшифровкой языка пчел Карл фон Фриш получил Нобелевскую премию (1973 г.).
Хочется отметить один факт: что у нас, что у пчел общая архитектура нейронов задана наследственно, а не обучением, индивидуальный опыт добавляет то, как они друг с другом соединятся на конечных стадиях. Мы отличаемся от пчел тем, что у нас гораздо большая часть нервных свзяей наследственно не задана, и на них ложится индивидуальный опыт.
Существуют две причины, почему отдельные независимые индивиды (будь это клетки или гидроидные полипы, или приматы) объединяются в колонию или многоклеточный организм. Это наиболее эффективное использование ресурсов и защита. На примере волков объясним первую причину. Летом волки живут небольшими группами, семьями. Они занимаются тем, что ловят разную живность, например мышей, которых летом много, и вполне успешно способны прокормиться. Но зимой мыши и другие мелкие насекомые прячутся под снегом, и приходится ловить крупных животных – лосей или оленей, например. Тогда волки сбиваются в стаи. Стаи могут осуществлять очень сложные виды загонной охоты, некоторые загоняют, некоторые сидят в засаде. Во время охоты волки обмениваются между собой вокальными сигналами, координируя таким образом свои действия. То есть здесь мы сталкиваемся с ситуацией, аналогичной поведению Dictyostelium'а, который часть времени проводит как одноклеточное, а часть времени – как коллектив одноклеточных, объединяющихся в единый организм.
Очень интересен образ жизни приматов. На картинке представлен детеныш шимпанзе бонобо. Как уже говорилось на первых лекциях, шимпанзе бывает двух видов: обыкновенный и бонобо, они разделились примерно 2 млн лет назад. И тот и другой виды равно близки к человеку. Эти виды очень похожи по образу жизни. Живут они в разных зонах, хотя ареалы их расселения перекрываются.

Шимпанзе живут отдельными группами, которые включают в себя несколько десятков особей, они ведут образ жизни охотников-собирателей, питаются они растениями, мелкими животными, объединяются в группы для охоты, несколько молодых самцов способны поймать мелкую добычу и съесть, также они едят муравьев, термитов, и используют орудия для того, чтобы их ловить. Например, для того, чтобы достать термитов, они разжевывают веточку, опускают в термитник, там на нее налипают термиты, и шимпанзе вынимает веточку и снимает их с нее губами. Для того, чтобы достать воду, они способны использовать губку из травы. Они могут разбивать орехи двумя камнями, один из которых – прототип молота, второй – наковальни. В группе существует достаточно строгая иерархия: есть лидирующий самец. Лидер выбирается не по силе и агрессивности, как раньше думали, а по способности к образованию максимального числа дружеских связей и способности к организации своих соплеменников на кооперативные действия. Все остальные члены группы ему подражают, маленьких детенышей воспитывают так, чтобы те приветствовали лидера и не вели себя плохо в его присутствии. Взаимодействия членов группы построены на некоторых видах поведения, например, вычесывание шерсти (груминг).

У человека сохранились от наших родственников приматов некоторые формы поведения, о которых будет рассказано на следующей лекции.