
Материя и мышление
| Вид материала | Документы |
- Жан-пьер шанжё, ален конн материя и мышление, 6789.87kb.
- Пространство и материя, 398.24kb.
- Мышление и его патология Мышление, 686.03kb.
- Понятие «критическое мышление» и его характеристики, 304.37kb.
- План: Понятие мышления в психологии. Виды мышления, 126.54kb.
- Триглав Природы Материя, 120.3kb.
- Заключение. Живая материя, 669.86kb.
- Материя. Сотворение. Этика и религия. Мокша. Общая оценка}, 934.61kb.
- Центр "синтез" Н. И. Сиянов. Триумф Виджл-воина. (Конспект), 693.84kb.
- I. Мышление как высший психологический процесс, 10.28kb.
4. От элементарных систем к мысленным объектам
Рассмотрим теперь следующий, более высокий, уровень организации — уровень нейронных систем (рис. 19). Очевидно, что нейроны могут объединяться друг с другом, а эти объединения могут специализироваться на таких особых функциях, как рефлекторные действия: ходьба, первые этапы зрительного восприятия... Например, сетчатка — это очень сложная сеть, которая, основываясь на данных об уловленных фоторецепторами фотонах, вырабатывает первое представление о внешнем мире. На втором уровне, во всей полноте реализованном только у беспозвоночных — таких, например, как земляной червь или слизень — происходит то, что этологи называют «закрепленными действиями»: отыскивать пищу, летать, спариваться, хватать добычу.
А. К.: То есть не важно, являются ли те или иные действия врожденными или приобретенным — мы все равно говорим, что это второй уровень организации.
Ж.-П. LLL: Это уровень элементарных контуров спинного мозга, позвоночного столба, так сказать, «мини-мозга». Малые совокупности нейронов у беспозвоночных собираются в нервные узлы, которые соединяются друг с другом многочисленными связями.
Далее идет другой уровень. Некоторым беспозвоночным, прошедшим особенно большой путь эволюции — таким, например, как спруты, поведенческие реакции которых близки к поведенческим реакциям позвоночных, — а также высшим позвоночным
4. ОТ ЭЛЕМЕНТАРНЫХ СИСТЕМ К МЫСЛЕННЫМ ОБЪЕКТАМ 107
Социальная жизнь Архитектура
Homo sapiens Млекопитающие Приматы à
Птицы
7Г Рыбы
Пчелы
Насекомые
омы Муравьи
Термиты — -—
(взаимосвязь совокупностей...)
_« Понимание»
(совокупность
нейронов)
Нейронные контуры (рефлекторные дуги, локальные контуры)
Нейроны
Молекулы
(нейромедиаторы нейропептиды, рецепторы, каналы, ферменты...)
Атомы (углерод, водород, кислород, азот...)
Рис. 19. Уровни организации нервной системы.
и, в особенности, человеку свойственно выстраивать так называемые «репрезентации». Их нервная система обладает способностью собирать нервные клетки вместе и тем или иным образом их кодировать — например, навык управления транспортным средством можно рассматривать как частный случай «моторной» репрезентации. Однако существуют также и репрезентации сенсорного типа, репрезентации более «абстрактные». Иерархическая организация в многочисленных нейронных контурах, о которых мы уже говорили, заменяется при этом параллельной организацией.
Нейронные основы кода, используемого в таких репрезентациях, очень подробно изучены Георгопулосом [38] на примере указывания кистью руки у развитой обезьяны. Он регистриро-
108
НЕЙРОННЫЙ МАТЕМАТИК
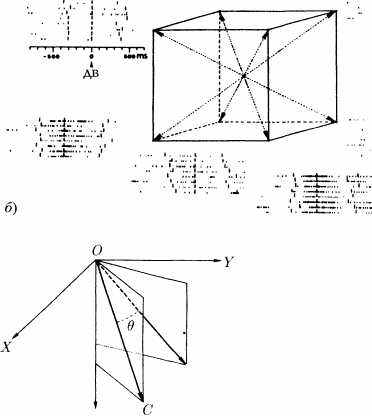
вал индивидуальную активность нескольких сотен нейронов двигательной зоны коры, в то время как обезьяна указывала рукой в заданном направлении. Георгопулос пытался определить, каким образом двигательная программа кодируется (или «репрезентируется») на уровне популяции регистрируемых нейронов. Он смог показать, что каждая клетка этой популяции проявляет максимальную активность, когда обезьяна указывает рукой в некоем особом, или предпочтительном, направлении, отмечая тем самым его специфичность (см. рис. 20). Для каждого нейрона определяется вектор, ориентация которого соответствует оптимальному направ-
а)
M
-z
4. ОТ ЭЛЕМЕНТАРНЫХ СИСТЕМ К МЫСЛЕННЫМ ОБЪЕКТАМ 109
лению, а длина определяется активностью этого самого нейрона в тот момент, когда обезьяна протягивает руку в каком-то определенном направлении. Эта длина меняется при изменении направления, в котором указывает рука. Она соответствует, в некотором роде, «голосу» этого конкретного нейрона или «налогу», который нейрон выплачивает ансамблю популяции за процесс кодирования движения руки в соответствующем направлении. Более того, направление, в котором указывает рука обезьяны, представляет собой (с менее чем 10%-ной погрешностью) векторную сумму элементарных «нейронаных» векторов. Вектор направления, в котором укажет рука, равен векторной сумме «голосов» популяции нейронов.
А. К.: Значит, сложение? Просто потрясающе. Можно представить все это в декартовых координатах... !
Ж.-П.Ш.: Да, через индивидуальные активности нейронов. Векторная сумма этих «микроскопических» активностей очень близко соотносится с вполне «макроскопическим» направлением,
Рис. 20. Состояние активности одного и того же нейрона двигательной зоны коры головного мозга бодрствующей обезьяны в тот момент, когда она пыталась дотянуться рукой до цели, помещаемой последовательно в каждом из восьми направлений трехмерного пространства, представленных на рисунке стрелками.
а) — Зарегистрированная электрическая активность представлена короткими вертикальными штрихами, каждый из которых соответствует одному нервному импульсу. Каждая линия из штрихов соответствует одной из попыток обезьяны. Общая для всех записей вертикальная линия (ДВ) отмечает начало движения. Рассмотрение всего множества записей показывает, что данный нейрон отвечает оптимальным способом (наиболее плотное расположение штрихов), когда обезьяна направляет свою руку в нужном направлении: в данном случае по направлению на 4 часа 30 минут в плоскости рисунка.
б) — Амплитуда отклика (частота разряда), когда обезьяна указывает рукой в заданном направлении (М) есть линейная функция косинуса угла между направлением движения и предпочтительным направлением (С) нейрона. Эту амплитуду можно рассматривать как «налог», который нейрон платит за кодирование движения всему ансамблю популяции. (Рисунок из работы [37])
110
НЕЙРОННЫЙ МАТЕМАТИК
90°

Вектор популяции
Клеточный вектор
Направление движения Векторная гипотеза
Траектория движения
Интервал доверия вектора популяции
Рис. 21. Кодирование популяцией нейронов двигательной коры направления движения на полдень (90°) на двумерной рабочей плоскости. Хорошо тренированная обезьяна реализует целое семейство достаточно сжатых траекторий движения. В соответствии с векторной гипотезой, сумма «голосов» каждого регистрируемого нейрона (общим счетом 241 клетка) или клеточных векторов, представленных здесь непрерывными линиями, дает вектор популяции (пунктирная линия), который сориентирован в направлении движения с интервалом доверия 99%. (Рисунок из работы [37].)
в котором указывает рука обезьяны. Происходит кодирование ансамблем нейронов, и я думаю, что эта модель носит общий характер (см. рис. 21). На определенном уровне сложности центральной нервной системы возникают «репрезентации» или «мысленные объекты» [9], которые можно описать как состояния активности нейронов популяции, так и по графам этих нейронов. Каждый нейрон популяции отличается от своего соседа. Каждый в рамках этого множества обладает функциональной специфичностью, индивидуальностью, «сингулярностью».
А. К.: Ты говорил, что такая способность свойственна и спруту, а ведь она вряд ли может быть врожденной.
4. ОТ ЭЛЕМЕНТАРНЫХ СИСТЕМ К МЫСЛЕННЫМ ОБЪЕКТАМ 111
Ж.-П.Ш.: Не будем сваливать все в одну кучу. В данный момент я пытаюсь определить уровни организации, и я еще не закончил.
Мы подходим, таким образом, к уровню, который можно назвать символическим или уровнем понимания. На этом уровне организации можно определить мысленные репрезентации в физических терминах. На более высоком уровне, который я называю уровнем разума, формируются цепочки представлений. «Совокупности совокупностей» эволюционируют во времени. Здесь становится крайне важным временной фактор, который мы еще не обсуждали достаточно подробно.
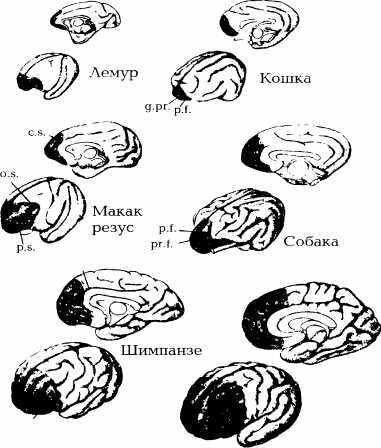
Самая передняя область головного мозга, фронтальная кора (рис. 22), по всей вероятности, выполняет так или иначе именно эту функцию. В качестве попытки иллюстрации этого утверждения приведу пример, взятый мною из клинических исследований пациентов, страдающих от поражения лобной доли. Классический пример — результат теста Милнера и Петридеса [81]. Исследователь предлагает испытуемому рассортировать несколько карт в соответствии с определенным правилом. Например, по цвету. Если три карты — красные, то и четвертая должна быть такой же, иначе исследователь отмечает ошибку. Затем, продолжают с тремя другими картами, тоже красными, и испытуемый выбирает красную карточку. Он все время следует одному и тому же правилу. Неожиданно, не предупреждая испытуемого, исследователь меняет стратегию. Правило будет, к примеру, касаться номинала карт: скажем, только тузы... Испытуемый сначала делает ошибки и продолжает выбирать красные карты, на что исследователь ему каждый раз указывает. Сделав несколько ошибок, испытуемый осознает, что стратегия изменилась. Больной, страдающий поражением фронтальной коры, к такому осознанию не приходит. Он упорно продолжает ошибаться. Но Милнеру и Нетридесу, испытуемый больной не формулирует предположений, которые позволяют продвигаться дальше в рамках этого теста. Он лишен элементарной, но характерной для уровня разума функции.
А. К.: Значит, наличие функции зависит от определенной зоны.
Ж.-Н. Ш.: Да. В начале у субъекта проявляются нейрологиче-ские нарушения — например, вследствие повреждения сосудов. Он консультируется у нейролога, который исследует его мозг путем сканирования. Определенные области лобной доли показы-
112
НЕЙРОННЫЙ МАТЕМАТИК
Человек
ip.f.
Рис. 22. Эволюция относительной площади поверхности префронтальной (выделена черным) коры головного мозга млекопитающих. Двигаясь от примитивных млекопитающих к Homo sapiens, можно наблюдать увеличение относительной площади поверхности префронтальной коры. У кошки она составляет 3,5%, у собаки — 7%, у лемура — 8,5%, у макака — 11%, у шимпанзе — 17% и у человека — 29%. По форме борозд и щелей префронтальной коры можно определить вид млекопитающего: р. f. = пре-сильвиева щель, Lp.f. = нижняя предцентральная щель. (Рисунок из работы [34])
вают нарушения. Тогда больной может быть подвергнут тестам, которые позволят определить функциональный дефицит, связанный с этими поражениями. И наоборот, нейролог может, исследуя пациента, распознать аномальный результат какого-либо теста
5. НЕЙРОПСИХОЛОГИЯ МАТЕМАТИКИ ИЗ
и диагностировать фронтальное поражение, которое затем подтвердится на сканограмме. Таким образом, лобная доля вносит определенный вклад в то, что я называю нейронными архитектурами разума [10]. Английский нейропсихолог Шаллис [91, 92] замечательно выразил суть этого различия. Он разделяет так называемые «рутинные» функции от функций «внимательного наблюдения», принимающих, по его мнению, участие в обнаружении ошибок, формулировании новых гипотез и изобретении новых стратегий. Таким образом, можно вполне резонно предположить, что в головном мозге существуют особые области, участвующие в производстве рациональной мысли. В поддержку этого заключения можно привести следующий замечательный факт: относительная площадь поверхности лобной доли в сравнении с остальной поверхностью новой коры (неокортекса) в ходе эволюции растет, причем у обезьян эта относительная площадь значительно больше, чем у крыс, а у людей — значительно больше, чем у обезьян.
А. К.: Это в точности соответствует моему описанию второго уровня.
Ж.-П. Ш.: В данном конкретном случае, — да. Но здесь может быть замешан также и третий уровень, хотя тесты, о которых шла речь, не показывают этого с достаточной очевидностью.
А. К.: Разделение второго и третьего уровня следует сохранить. Твои замечания относительно лобной доли, отвечающей за функции второго уровня, меня убедили. Что касается третьего уровня, то об этом я ничего не знаю.
Ж.-П. Ш.: Лобная доля играет важную роль в генерации предположений. Предположения, выдвигаемые при игре в карты Мил-нера и Петридеса, являются рудиментарными. Несомненно, на своем уровне осуществляется порождение и гораздо более сложных гипотез, но их очень сложно зарегистрировать. Для этого нам потребовалось бы поместить мозг Архимеда под камеру наблюдения за несколько долей секунды до возгласа «Эврика»!
5. Нейропсихология математики
Ж.-П.Ш.: В соответствии с различием уровней организации в функциях мозга, локализованные поражения головного мозга позволяют «добраться» и до математических способностей. Вели-
114
НЕЙРОННЫЙ МАТЕМАТИК
кий французский нейропсихолог Экаэн [51], с которым ты, может быть, знаком, различает несколько категорий дефицита.
В случае «цифровой алексии и/или аграфии» субъект не в состоянии читать и/или писать цифры, но может сохранять умение пользоваться буквами. Экаэн смог показать, что за чтение и написания цифр отвечает левое полушарие — точнее, теменная доля левого полушария.
Пациенты, страдающие от «пространственной акалькулии», не могут строить ряды из цифр. Этот дефицит, по-видимому, связан с системой визуально-двигательной ориентации, которая позволяет одновременно читать и упорядочивать цифры. В этом случае за управление движением глаз отвечает, скорее всего, правое полушарие.
Третий дефицит называется «анарифметия». Это дефицит собственно способности к счету. Пациент не в состоянии совершать арифметические действия, независимо от своей способности читать, писать и правильно располагать числа. Во всех случаях речь идет о дефицитах, связанных с тем первым уровнем, о котором мы говорили выше.
Интересное подразделение делает Лурия. Он полагает, что все перечисленные дефициты связаны с теменно-затылочной областью коры. Они, по его мнению, отличны от тех, что поражают височную долю и вызывают нарушение памяти. Поражения этого типа приводят к тому, что субъект перестает запоминать то, что он только что сделал. Ему не удается следовать за нитью своих вычислений.
Больные, страдающие поражением лобной доли, имеют нарушения другой природы. Они не воспринимают задачу, которую им необходимо решить. Они теряют нить размышления, им не удается формулировать последовательное рассуждение, они дают импульсивные ответы, в какой-то мере случайные, и упорствуют в своей ошибке. Для обнаружения фронтальных поражений применяется среди многих других тестов и цепочка последовательных вычитаний. Можно также предположить, что лобная доля отвечает за последовательное выполнение математических операций, за решение и даже постановку задач. Возможно, это соответствует второму или третьему уровню.
А. К.: Только не третьему.
Ж.-П.Ш.: Используемые в данном случае тесты являются по необходимости элементарными.
5. НЕЙРОПСИХОЛОГИЯ МАТЕМАТИКИ 115
А. К.: И все равно невозможно разработать тест для третьего уровня.
Ж.-П.Ш.: Почему же невозможно? Вообрази себе, например, такой тест, который могли бы использовать не-математики. Еще один тест, предлагаемый больным с поражением лобной доли, состоит в том, что их просят прочитать, а затем пересказать какую-нибудь историю. Им читают, например, «Красную Шапочку» (Лер-митт) или «Золотого Петушка» (Лурия), а затем просят восстановить историю.
А. К.: Здесь мы все еще остаемся на втором уровне.
Ж.-П.Ш.: Нет, элементы истории восстанавливаются, но целый текст бывает несвязным. Конец перемещается в начало, порядок следования эпизодов путается...
А. К.: Но это же порядок организации, воображение в этом не участвует.
Ж.-П.Ш.: Это верно. Однако найди мне объективный тест воображения. Нейропсихологи будут тебе очень благодарны.
А. К.: Я не знаю такого теста. Однако хочу задать тебе вопрос: часто говорят, что математики теряют творческий потенциал, когда стареют. И это довольно известное явление. Что ты об этом думаешь?
Ж.-П.Ш.: Лобная кора подвержена относительно быстрому старению — в частности, при болезни Альцгеймера. Больные, страдающие этим заболеванием, действительно, могут очень быстро потерять память и способность к счету. То, что они при этом теряют научный творческий потенциал — также вполне правдоподобно. ..
А. К.: Наверняка возможно более четкое различение второго и третьего уровней...
Ж.-П.Ш.: Очень сложно, особенно в рамках обычных операций. Больной с фронтальными нарушениями — какой описан, например, у Лермитта [74] — это больной, который «вписывается в окружающий мир». Когда ему дают какой-либо предмет, он его использует. Ему дают ручку, он ей пишет. Очки он надевает на нос. Молотком забивает гвоздь. Он находится в прямой связи с внешним миром, сохраняя владение речью. Он нормально выполняет рутинные виды деятельности, но ему не удается решать задачи, которые относятся к новым ситуациям. Непредвиденность — это значительное препятствие для человека, страдающего некоторыми видами нарушений лобной доли.
116
НЕЙРОННЫЙ МАТЕМАТИК
6. Переход с уровня на уровень посредством вариации-селекции
Ж.-П. Ш.: Думаю, для уточнения различий между вторым и третьим уровнями нам следует задать новый вопрос. Как мы переходим с уровня на уровень? В течение нескольких лет [8, 12, 13, 15] я разрабатывал предположение (аналогичные идеи можно найти и у других авторов [31, 30, 65] ), суть которого заключается в том, что переход с уровня на уровень (причем какой угодно уровень) можно описать в терминах этакого «обобщенного дарвинизма» (см. рис. 23). Переход с некоторого уровня на следующий нуждается в двух фундаментальных составляющих — генераторе разнообразия и системе отбора. Элементы начального уровня комбинируются между собой, меняются случайным образом и вырабатывают некие переходные «формы», в зависимости от следующего более высокого уровня организации. Эти формы порождаются
Доступ к верхним уровням
(социальным и т.д.)
уровень Z
Стабилизация и
усиление
| | | | | | | | | | |
| | ι ιμα.ογυ\α. | Opj. СииНОСЩИ·*! | " | " " | А | J г | | | |
| | правила | И устойчивость | l· | Регрессия 1 1 Регулирование между | уровнями | | |||
| | | À | | | | 1 | | | |
| | | 1 | | | | Y | — \mrmp>WT, | Y | |
| | | | | | | | | | |
Укоренение '
в нижних уровнях
(атомном и т.д.)
время
Рис. 23. Обобщенный дарвинизм
6. ПЕРЕХОД с УРОВНЯ НА УРОВЕНЬ ПОСРЕДСТВОМ ВАРИАЦИИ. .. 117
уже структурированными элементами, т.е. вовсе не обязательно атомами. Стало быть, имеет место производство «дарвиновских» вариаций, которые способны получить временный доступ к высшему уровню организации. Далее механизм селекции стабилизирует некоторые из этих переходных состояний и таким образом порождает более высокий уровень организации.
А. К.: Какой именно механизм селекции?
Ж.-П.Ш.: Общая модель выглядит примерно так:
материя —»- форма —- функция (вариации) Ψ
(стабилизация)
Функция действует ретроактивно на переход «материя-форма». Критерий отбора связан, таким образом, с «новой» функцией, определяемой переходной формой, произведенной генератором разнообразия. Если эта новая функция соответствует такому воздействию на внешний мир, которое благоприятствует выживанию организма, то она отбирается.
А. К.: Внутри мозга или вне его?
Ж.-П.Ш.: Я для начала попытался представить очень общую формальную модель, которая, надеюсь, действительна, каков бы ни был рассматриваемый уровень организации в первоначальном состоянии. Попробуем теперь ее применить. Самый простой и самый известный случай — эволюция видов. Генератор разнообразия находится на уровне генома, или хромосомной ДНК. «Дарвиновские» вариации — мутации, рекомбинации, дупликация генов, перенос хромосомного материала — представляют собой случайные, но редкие события, которые вызывают уже вторичные модификации «фенотипа» организма, которые могут сопровождаться «адаптацией» к тем или иным особым условиям окружения. Также может иметь место выделение особых генетических комбинаций, однако отбора при этом не происходит вследствие географической изоляции: этот процесс называют недарвиновской эволюцией, но мне этот термин не очень нравится. Некоторые «нейтральные» элементы впоследствии сохраняются, другие же исчезают.
Нервная система представляет собой точно такой же орган, как и другие. Однако она имеет особый статус. Соединения между нервными клетками, синапсы, не возникают в мгновение ока —
118 НЕЙРОННЫЙ МАТЕМАТИК
они образуются в результате долгого и сложного процесса развития, который продолжается у человека вплоть до наступления половой зрелости. То есть в нервной системе происходит внутренняя по отношению к организму эволюция. Более того, различается, по крайней мере, два типа внутренней эволюции: эволюция количества связей, происходящая в процессе развития организма, и эволюция эффективности связей между нейронами, т. е. их состояний активности, что, впрочем, протекает гораздо легче, чем изменение связности.
Рассмотрим сначала первый тип эволюции — эволюцию посредством «эпигенеза», происходящую во время эмбрионального и постнатального развития организации мозга. Для начала предположим, что «человечность» формирующегося мозга (в смысле его отличия от мозга, скажем, обезьяны) определяется неким глубочайшим генетическим детерминизмом церебральной организации. Участвующие в этом развитии гены сейчас активно исследуются у позвоночных. Очень подробно они были исследованы у мухи-дрозофилы, которая, пусть и всего лишь муха, имеет, как и мы, голову, грудную клетку, брюшную полость и конечности. Не так давно [36, 86] были идентифицированы некоторые из генетических детерминант, фиксирующих декартовы координаты эмбриона (по осям «голова-хвост», «спина-живот»), регулирующих сегментацию тела (т. е. тело формируется в виде последовательных сегментов — примерно как у червей) и, наконец, идентифицирующих собственно сегменты (головной, с антеннами и жвалами; грудной, с крыльями и лапками; брюшной, с генитальны-ми органами и т.д.). В течение эмбрионального и постнатального развития эти три совокупности генов проявляют себя дифференциально и последовательно. Результатом этого чередования генетических проявлений является организм, обладающий целостной архитектурой, планом организации, который в рамках одного вида остается неизменным (или почти неизменным) от одного индивида к другому.
Можно предположить, что увеличение площади фронтальной коры, которое мы наблюдаем у млекопитающих (от мыши к человеку) происходит под влиянием некоторых из упомянутых генов. Их число, вероятно, не очень велико. В самом деле, ДНК шимпанзе на 99% идентична ДНК человека. Если какие-то из этих генов остаются активными в передней части эмбрионального зачатка мозга более длительное время, то следствием этого будет дифференци-
6. ПЕРЕХОД с УРОВНЯ НА УРОВЕНЬ ПОСРЕДСТВОМ ВАРИАЦИИ. .. 119
альное увеличение площади поверхности фронтальной коры. Таким образом, общая организация головного мозга человека, основной части нашей церебральной архитектуры, находится во власти генов.
Тем не менее, власть генов имеет свои пределы. Как же их обнаружить? Можно для начала сравнить связность одного и того же нейрона у двух генетически идентичных индивидах (например, у двух однояйцевых близнецов); идентифицировать нейрон можно по его форме и расположению. Опыт был осуществлен Левента-лем [50] на партеногенетических ракообразных дафниях — водяных блохах, обладающих упрощенной нервной системой с фиксированным числом нейронов, причем все эти нейроны располагаются в пространстве всегда одинаково (или почти одинаково). С помощью партеногенеза можно легко получить несколько генетически идентичных индивидов, называемых «изогенными». После чего достаточно разрезать их на тонкие слои, исследовать под электронным микроскопом и сравнить у этих индивидов полное аксонное дендритообразование одного и того же нейрона. Результаты исследования показывают, что главные линии связности у разных индивидов сохраняются неизменными, однако в деталях, на уровне распределения синаптических контактов, появляется определенная вариантность.
Второе «доказательство»: исследование эволюции церебральной связности как функции от объема приобретенного опыта. Маленького котенка или новорожденную обезьяну помещают на так называемый «чувствительный» постнатальный период в искусственное визуальное окружение, отличное от того, в каком обычно развиваются эти животные. В результате у взрослого животного очень сильно и часто необратимо нарушается функциональная специализация индивидуальных нейронов зрительной коры (специфика ориентации, бинокулярность зрения и т.д.). Человек также может по несчастному стечению обстоятельств приобрести подобный «опыт» — например, вследствие врожденной катаракты. Непрозрачность хрусталика на ранних стадиях развития организма приводит к визуальному дефициту и даже к слепоте, сохраняющейся и после операции катаракты (если она была проведена после окончания вышеупомянутого чувствительного периода), т.е. к слепоте на уровне коры головного мозга. Эти опыты, наряду с многими другими, говорят о том, что «установление» взрослой связности обусловлено активностью нервной системы
120
НЕЙРОННЫЙ МАТЕМАТИК
в процессе ее развития. Вместе с Филиппом Курежем и Антуа-ном Даншеном [12] мы предложили формальную модель эволюции связности системы нейронов в процессе ее развития по дарвиновской схеме в рамках данной модели; структура генетического материала при этом не изменяется, что позволило нам квалифицировать нашу модель как «эпигенез» посредством селективной стабилизации синапсов. Основная идея заключается в том, что генетические детерминанты, ответственные за распознавание отдельных нейронов, входящих в обе группы клеточных партнеров, одинаковы или почти одинаковы. Для того, чтобы закодировать эту способность к распознаванию, достаточно небольшого числа генов. На определенной критической стадии развития (в чувствительный период) два ансамбля нейронов входят в контакт. Я говорю здесь не о сформировавшихся контактах между нейроном χ первой группы и нейроном у второй группы, но о процессе «установления контакта», бурном, нечетком, множественном и перекрывающемся. На этой стадии можно наблюдать впечатляющее разнообразие всевозможных связей. В действие вступает генератор «дарвиновских вариаций»! Далее происходит «доводка», позволяющая, посредством стабилизации некоторых соединений и удаления других, установить взрослый тип связности. С помощью такой модели можно моделировать простые ситуации обучения, а также более сложные ситуации, возникающие в процессе развития, в частности, человека. В человеческом мозге последовательные волны формирования и отбора синапсов следуют друг за другом, сливаются, накладываются друг на друга последовательными притоками и оттоками... в течение длительного времени после рождения. Необходимо, разумеется, уточнить биологические ограничения, которые ведут к отбору предпочтительных соединений. Эти правила отбора должны зависеть от организма в его целостности и во взаимодействии с внешним миром.
А. К.: А почему бы не использовать все эти связи, коль скоро они уже сформированы? Их просто необходимо использовать. Чем объясняется отбор?
Ж.-П. Ш.: Свой вклад в моделирование конечного состояния сети вносит присущая системе активность. А именно она у нейронов не идентична в точности. В предложенной модели эволюция того или иного синапса определяется локальными правилами эволюции в зависимости от собственного состояния активности
6. ПЕРЕХОД с УРОВНЯ НА УРОВЕНЬ ПОСРЕДСТВОМ ВАРИАЦИИ. .. 121
синапса и от состояния клетки, на которую эта активность проецируется. Например, как я уже говорил, совпадение активности двух контактирующих клеток может вызвать стабилизацию этого контакта. Обучение приводит к новому отношению «вход/выход». После обучения одинаковый сигнал на входе всегда дает одинаковый сигнал на выходе, тогда как до обучения сигналы на выходе от опыта к опыту варьировались.
Описанная формальная модель обладает интересным математическим свойством, которое можно сформулировать в виде теоремы изменчивости. Согласно этой теореме, одинаковое отношение «вход/выход» после обучения можно получить даже в тех случаях, когда в результате отбора сохраняются разные связности. Это вполне согласуется с наблюдениями изменчивости связности, о которых я только что говорил. Известно также, что хотя у большинства людей речевые центры расположены в левом полушарии, есть люди, у которых эти центры размещаются в правом полушарии или распределены по обоим полушариям. Причем по одной лишь манере речи отличить одних от других еще никому не удавалось. Нейрофенотип лее, несмотря на очевидную схожесть функций, изменяется очень сильно. Мы подходим, таким образом, к заключению, крайне значимому в рамках нашего обсуждения. Несмотря на существенные различия в тонкой церебральной организации отдельных математиков, им как-то удается одинаково воспринимать своим мозгом одинаковые математические объекты.
Завершив рассмотрение нейронного дарвинизма эволюции связности, коснемся и другой эволюции — эволюции более высокого уровня, которую можно охарактеризовать как дарвинизм ментальный [14] или психологический. (Это понятие встречается еще у Фрейда: см. [98, с. 244].) Нейронный дарвинизм в ходе развития организма проявляется, главным образом, в самом раннем детстве или еще в эмбриональном состоянии. Эмбрион активен, он проявляет спонтанную активность, которая может вмешаться во «внутренний» отбор синапсов, обеспечивающих координацию между различными нервными центрами. Что до ментального дарвинизма, то он имеет отношение, по большей части, к взрослому мозгу — как на уровне понимания, так и на уровне разума. На шкале психологического времени он производит, скорее, изменения синоптической эффективности, чем эволюцию количества связей. Единицами отбора здесь служат не просто связи
122
НЕЙРОННЫЙ МАТЕМАТИК
и элементарные контуры, но совокупности нейронов, способных на координированную активность, Они выбираются из элементов, уже отобранных в результате нейронной эволюции. Генератор разнообразия является здесь не результатом изменчивости связей в процессе развития, но результатом спонтанного и неустойчивого начала активности совокупностей нейронов — так называемой «пре-репрезентации». Возникает активность комбинаторного типа, предвосхищающая взаимодействие с внешним миром. Если в результате этого процесса достигается некая «конгруэнтность», «резонанс» между внутренним и внешним состояниями системы, то пре-репрезентации стабилизируются, «запасаются» в сети. Если резонанса нет, то никакого запечатлевания не происходит. Такое запоминание изменяет величины синаптической эффективности, составляющие в совокупности сохраняемую конфигурацию системы.
Последовательность ментальных репрезентаций, вызываемых процессом размышления в «рабочем разделе» краткосрочной памяти, имеет в своей основе аналогичный ментальный дарвинизм. Применима ли такая модель к работе математика? Можно предположить, что в течение периода «инкубации» различные репрезентации математических объектов сменяются, переходя друг в друга и следуя друг за другом достаточно произвольно. Затем среди репрезентаций или пре-репрезентации происходит нечто вроде внутреннего отбора посредством резонанса. Этот отбор приводит к «объекту-результату», который согласуется с поставленной задачей, т.е. с «интенцией», на которую следовало должным образом отреагировать. На той стадии, на которой мы сейчас находимся, эти идеи носят пока еще очень схематический характер, и я не думаю, что в данный момент их можно сформулировать сколько-нибудь точнее.
Совместно со Станисласом Деэном и Жан-Пьером Надалем мы разработали модель [20], пока еще очень элементарную, сети нейронов, расположенных последовательными слоями, которая способна распознавать порядок следования символов или мелодию, запоминать их и впоследствии воспроизводить. Эта модель дает хорошее объяснение процессу обучения некоторых птиц пению (см. рис. 24). Сейчас мы вплотную приближаемся к возможности детально моделировать некоторые этапы протекания мысли, однако до достижения успеха предстоит еще немало работы.
6. ПЕРЕХОД с УРОВНЯ НА УРОВЕНЬ ПОСРЕДСТВОМ ВАРИАЦИИ. .. 123
Стадия
Начальные вокализации
6 4 VII
ί
IV
IV
б
4 III
Пластическое пение

слог 4
слог 4
слог 5
слог 2
46ί· viV'TnT1'1'
4+ * · * V \ xiJv t»*N * ι
IV
слог 1 слог
б+ ' ΥΠΜΤΓ 'lYIMvn'î'l' ΊΥ>
4+ JjJJJJJJJJJJJJJs JJJ
слог 3
слог 3 Кристаллизованное пение

слоги для обучения
Рис. 24. Обучение болотного воробья пению. Линиями представлена частота производимых звуков в зависимости от времени. Внизу справа показаны слоги, которым обучается воробей. Птенец слышит и запоминает эти слоги в период между 22-м и 62-м днями после появления на свет. Примерно через 200 дней птенец производит первые вокализации и выстраивает их в слоги, услышанные за семь месяцев до этого. В песне взрослой птицы остается только слог № 3. Кристаллизация пения сопровождается потерей (или отсевом) слогов, что свидетельствует о «селекционном» характере обучения. (Рисунок из работы [77])
124
НЕЙРОННЫЙ МАТЕМАТИК
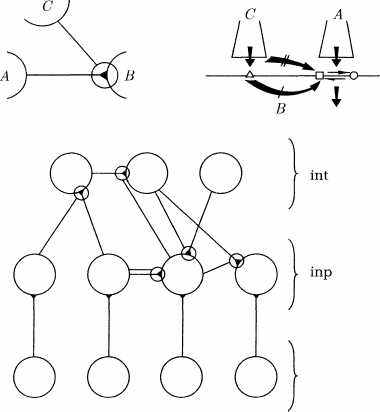
sens
Рис. 24 α. Формальная нейронная сеть, способная распознавать, воспроизводить и записывать в памяти посредством отбора временные последовательности «репрезентаций». Архитектура очень проста: три слоя нейронов (сенсорные, обозначенные на рисунке сокращением sens, входные — inp и внутренние — int), разделенные на группы самовозбуждающихся нейронов (обозначенные кругами), кодирующих «репрезентации». Между собой нейроны соединяются тройками синапсов ЛВС, эффективность которых модулируется химически. Подробнее см. работу [20].