
О структуре комплекса зеленых лягушек в раифском участке волжско-камского заповедника р. И. Замалетдинов
| Вид материала | Документы |
- Организация паромно-транспортных логистических систем (на примере волжско-камского, 330.99kb.
- Комитета Государственной Думы Федерального собрания Российской Федерации по природным, 66.82kb.
- Федеральная служба по надзору в сфере транспорта волжское управление государственного, 422.88kb.
- О. В. Динамика орнитофауны Ульяновской области (Россия, Среднее Поволжье) в конце, 23.85kb.
- Урок 28 «Война лягушек и мышей», 1155.42kb.
- X-man среди лягушек, 9.8kb.
- Молодость старого заповедника, 58.33kb.
- Выставка работает до 12 июня 2011, 100.5kb.
- Оборудование: овощи, выращенные на участке, на собственных огородах, фотовыставка «Урожай, 135.25kb.
- Ноу впо российская международная академия туризма волжско Камский филиал, 925.63kb.
О СТРУКТУРЕ КОМПЛЕКСА ЗЕЛЕНЫХ ЛЯГУШЕК
В РАИФСКОМ УЧАСТКЕ
ВОЛЖСКО-КАМСКОГО ЗАПОВЕДНИКА
Р.И. Замалетдинов,1 Л.Я. Боркин,2 С.Н. Литвинчук,3 Ю.М. Розанов3
Zamaletdinov R.I., Borkin L.Ya., Litvinchuk S.N., Rosanov Yu.M.
On the green frogs complex structure in VOLZHSKO-KAMSKY
Zapovednik Raifa part
The Rana esculenta complex was analyzed by flow-cytometry on the materials of 2002 and 2003 years. It is established that R. ridibunda Pall., R. lessonae Camer. and R. kl. esculenta L. inhabit the zapovednik and its buffer zone. Data on species ecology and biotopic distribution are presented; 4 types green frogs population systems are found.
Европейские зеленые лягушки (комплекс Rana esculenta) изучаются биологами на протяжение уже более чем двух столетий. Особый интерес к этой группе земноводных проявился в последние сорок лет, с момента открытия польским зоологом Л. Бергером (Berger, 1967; Gűnther 1990) формы гибридогенного происхождения, которая, первоначально, еще в 1758 г., была описана К. Линнеем как Rana esculenta. Своеобразие генетических способов воспроизводства этой формы, а также встречаемость у нее полиплоидии и необычные популяционные особенности привлекают к себе внимание многих исследователей из разных стран, а сама съедобная лягушка стала рассматриваться как интересная модель неортодоксального видообразования (Боркин, Даревский, 1980).
В настоящее время считается, что зеленые лягушки в Восточной Европе представлены тремя таксонами (Боркин, 1998; Кузьмин, 1999): лягушки озерная (R. ridibunda Pallas, 1771) и прудовая (R. lessonae Camerano, 1882) являются «хорошими», «менделевскими» видами, а съедобная лягушка (R. kl. esculenta Linneus, 1758) представляет собой особую форму, произошедшую от гибридизации указанных выше родительских видов; она имеет большой современный ареал – от Франции до Поволжья, – и большинством специалистов признается в качестве самостоятельного вида.
Проблема полевых исследований комплекса зеленых лягушек во многом связана с их точной видовой диагностикой. Принято считать, что между видами существуют более или менее четкие морфологические различия (Банников и др., 1977; Таращук, 1985; Борисовский и др., 2000; Коцержинская, 2000 и др.). Однако точность видовой идентификации при использовании только морфологических показателей вызывает в ряде случаев законное сомнение, так как она не всегда подтверждается более точными методами, например, такими как проточная ДНК-цитометрия (Боркин и др., 1987; Виноградов и др., 1988) или электрофорез белков (Tűnner, 1970).
В свете гибридогенной концепции, значительная часть ареала зеленых лягушек, расположенная в европейской части России, до настоящего времени остается относительно мало изученной. Однако именно здесь все три вида могут встречаться не только симпатрично (т.е. с перекрыванием своих ареалов), но и синтопично (т.е. в одном и том же водоеме).
На территории Татарстана до недавнего времени видовую идентификацию зеленых лягушек проводили только на основе морфологических показателей (Гаранин, 1983; Корчагина, Гаранин, 1996; Garanin, 2000; Павлов и др., 2001; Галеева и др., 2002). В связи с этим вопрос о видовом составе комплекса оставался до конца не решенным. Впервые точную видовую идентификацию зеленых лягушек, обитающих на территории Татарстана, удалось провести лишь в 2002 г. с помощью метода проточной ДНК-цитометрии в Институте цитологии РАН (Боркин и др., 2003). В этой работе был использованы сборы лягушек и из Волжско-Камского заповедника.
В настоящем сообщение мы дополняем и обобщаем данные по видовому составу, пространственному размещению и популяционным системам зеленых лягушек Раифы.
Материалы и методика
Материал был собран на территории Раифского участка Волжско-Камского заповедника и его охранной зоны в течение полевых сезонов 2002 и 2003 гг. на семи озерах: Белое, Раифское, Ильинское, Илантово, Гнилое, Линево и Круглое. Указанные водоемы различаются между собой по размерам, происхождению, по положению в ландшафте.
Первичная видовая идентификация лягушек в природе проводилась по внешним признакам. Нами использовались следующие морфологические признаки: сочленение голеностопного сустава, форма внутреннего пяточного бугра (рис. 1), а также цвет резонаторов у самцов. Все перечисленные признаки являются общепринятыми при определении зеленых лягушек (Банников и др., 1977).
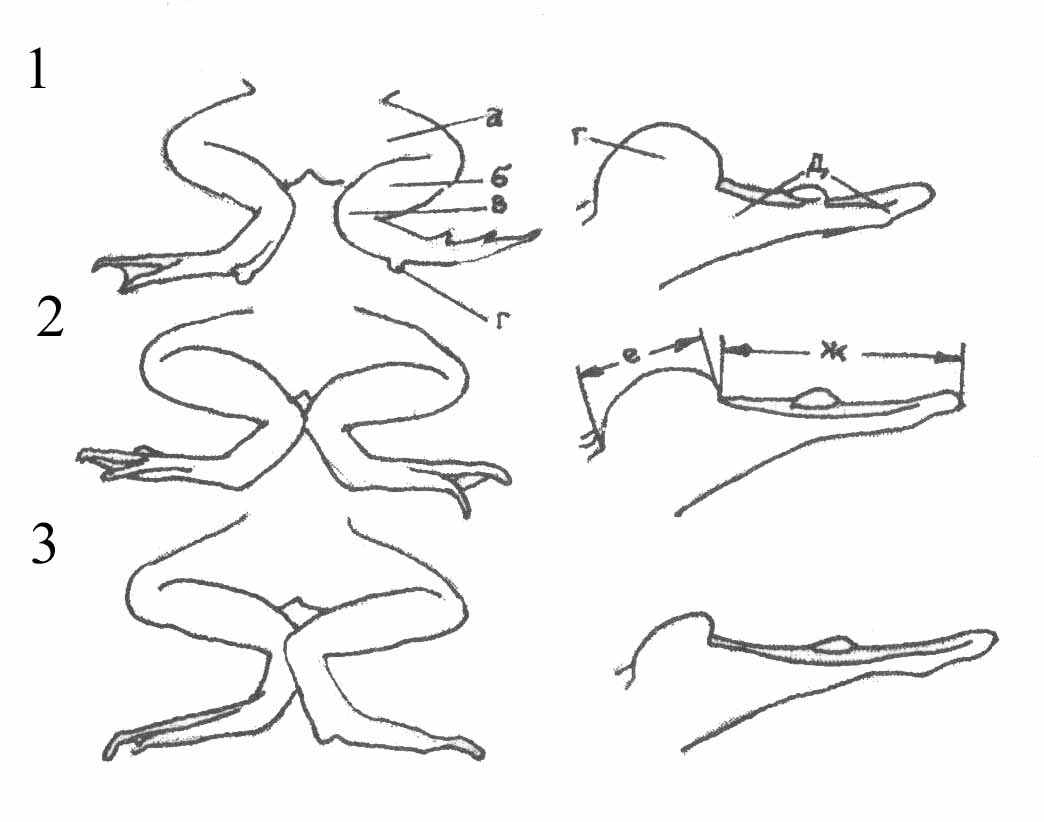
Рис. 1. Внешний вид сочленения голеностопного сустава и внутреннего
пяточного бугра у прудовой (1), съедобной (2) и озерной (3) лягушек
(по А.Г. Банников и др., 1977).
Условные обозначения: а – бедро; б – голень; в – голеностопное сочленение; г – внутренний
пяточный бугор; д – первый палец задней конечности; е – длина внутреннего пяточного бугра; ж – длина первого пальца задней конечности.
На основе части отловленных лягушек первичное определение было проверено в Институте цитологии РАН (г. Санкт-Петербург) методом проточной ДНК-цитометрии. В основе метода проточной ДНК-цитометрии лежит измерение массы генома клеток крови или клеток семенников. Установлено, что в пределах изучаемого комплекса видов существует четкая дифференциация по данному показателю. Так, масса ядерной ДНК у прудовой лягушки меньше, у озерной лягушки больше, а у съедобной лягушки этот показатель имеет промежуточные значения (Боркин и др., 1987).
Краткая характеристика озер
Озера Раифского участка Волжско-Камского заповедника и его охранной зоны приурочены к долинам рек Сумка и Сер-Булак. Все водоемы Раифы образуют единую гидросистему (рис. 2). По гидрохимической классификации О.А. Алекина воды озер Раифы относятся к гидрокарбонатному классу кальциевой группы с малой и средней минерализацией. Гидрохимический режим основных ионов характеризуется относительным постоянством (Унковская и др., 2002). По ландшафтной приуроченности условно можно выделить: типичные лесные озера, озера открытых пространств и опушечные озера.
Рис. 2. Карта-схема расположения озер Раифы.
Озеро Белое – открытый водоем, площадью 6,4 га, имеет суффозионно-карстовое происхождение (Тайсин, 1996). Через озеро протекает р. Сумка. Озеро расположено в охранной зоне заповедника.
Озеро Раифское – самый крупный водоем Раифского участка заповедника, его площадь 31,9 га, происхождение также суффозионно-карстовое. В озеро впадает р. Сер-Булак, через озеро протекает р. Сумка. В своей южной части, ниже линии уреза р. Сер-Булак и около выхода р. Сумка, озеро заболочено. Вода характеризуется невысокой прозрачностью в летний период. Температурный режим озера соответствует режиму глубокого стратифицированного водоема.
Озеро Ильинское (Уразлино) – крупный открытый водоем, площадью 21,7 га. Через озеро протекает р. Сумка. В северо-западной части озеро заболочено. Вода характеризуются относительно высокой прозрачностью. Температурный режим водоема аналогичен температурному режиму оз. Раифское. Озеро расположено в охранной зоне заповедника.
Озеро Илантово – интенсивно зарастающий лесной водоем, площадью 4,8 га. Физико-химические показатели его воды близки к болотным водам. Даже в летний период, при нормальном содержании кислорода в поверхностном слое воды, у дна отмечается его дефицит (Унковская и др., 2002). Озеро возникло на месте заболоченного леса путем просадки грунта (Тайсин, 1969; 1996).
Озеро Гнилое – лесной водоем, площадью 0,8 га, представляющий собой «окно» в сфагновой сплавине. До последнего времени озеро подвергалось загрязнению со стороны прилегавшей к нему зверофермы. Максимальная глубина, включая донные отложения, достигала 11 м (Тайсин, 1969); в настоящее время водная толща составляет 4,9 м (Унковская и др., 2002).
Озеро Линево – лесной водоем, площадью 6,9 га. Через озеро протекает р. Сер-Булак. Северный и юго-западный берега озера заболочены. Озеро возникло на месте хвойного леса путем образования глубокого провала. Мощность озерных отложений достигает 5 метров и практически равна толще воды (Тайсин, 1969; 1996; Унковская и др., 2002).
Озеро Круглое – небольшой водоем, площадью всего 0,5 га, расположенный на опушке леса около пос. Садовый. Озеро испытывает определенный антропогенный пресс со стороны прилегающих садовых участков (Унковская и др., 2002).
Видовой состав и биотопическое распределение
зеленых лягушек
На территории Раифы отмечены все три вида европейских зеленых лягушек. Первоначальное определение видов на основе морфологических признаков в большинстве случаев было подтверждено методом проточной ДНК-цитометрии (табл. 1). Вместе с тем, зафиксированные в ряде случаев ошибки первоначального определения дают основание для сомнения в надежности определения видов зеленых лягушек по морфологическим признакам, что отмечается и в литературе (Pagano, Joly, 1998; Борисовский и др., 2000; 2001).
Таблица 1
Распределение массы генома (пг) у зеленых лягушек,
обитающих на территории Раифы
| Вид | Масса генома | ||
| n | M±m | min-max | |
| Rana lessonae | 16 | 13,88±0,018 | 13,76-14,00 |
| Rana kl. esculenta | 8 | 14,89±0,021 | 14,79-14,98 |
| Rana ridibunda | 13 | 16,12±0,028 | 15,98-16,28 |
Согласно целому ряду проведенных исследований, принято считать, что родительские виды R. kl. esculenta, т.е. озерная и прудовая лягушки, достаточно хорошо различаются по своей биотопической приуроченности (Berger, 1976; Гаранин, 1983; Лада, 1995; Боркин, 1998; Кузьмин, 1999 и др.). Озерная лягушка заселяет преимущественно крупные глубокие водоемы, как непроточные, так и проточные, но в открытых ландшафтах. Исключение составляют популяции этого вида, обитающие в условиях сильного антропогенного загрязнения среды, при которых озерные лягушки могут заселять и небольшие водоемы. Прудовая лягушка, напротив, обитает преимущественно в небольших лесных замкнутых водоемах, в том числе пересыхающих и заболачивающихся. Съедобная лягушка приурочена, как правило, к биотопам промежуточного характера, а именно – к озерам лесных опушек (Лада, 1995; Корчагина, Гаранин, 1996). Также она может встречаться и в водоемах, характерных для одного из родительских видов, совместно с ним, заселяя, в основном, более характерные для себя биотопические микроучастки. Съедобная лягушка избегает как крупных открытых, так и мелких заболачивающихся водоемов.
Установлено, что биотопическая дифференциация зеленых лягушек связана с особенностями мест их зимовок. Озерная лягушка зимует преимущественно на дне водоемов, зарывшись в ил (в связи с чем, пяточный бугор у нее низкий, приплюснутый). Прудовая лягушка зимует на суше (для закапывания в грунт в качестве приспособления она имеет пяточный бугор высокий, «лопатовидный»). Съедобная лягушка способна зимовать как на дне водоема, так и на суше (соответственно пяточный бугор у нее, как правило, имеет промежуточную форму).
На территории Раифы озерная лягушка была отмечена в характерных для нее биотопах: на открытых участках рек Сумка и Сер-Булак, а также на крупных и открытых озерах – Раифское, Ильинское и Белое. На оз. Раифское вид обитает вместе с прудовой и съедобной лягушками, придерживаясь открытых участков.
Прудовая лягушка встречается по рекам заповедника и на лесных озерах – Илантово, Гнилое, Линево, которые подвержены заболачиванию. Кроме того, этот вид был выявлен в озерах Раифское и Круглое. На оз. Раифское прудовая лягушка придерживается преимущественно мелководных и заросших участков.
Съедобная лягушка достоверно обнаружена только в озерах Круглое и Раифское. В обоих водоемах съедобная лягушка обитает совместно с одним (оз. Круглое) или с обоими (оз. Раифское) родительскими видами, занимая в биотопическом отношение промежуточное по отношению к озерной и прудовой лягушкам место.
Популяционные системы зеленых лягушек
Совместное обитание на одном водоеме разных видов зеленых лягушек обусловливает тот или иной тип популяционной системы. В ареале зеленых лягушек возможны как «чистые» (содержащие только один вид), так и смешанные (содержащие два или три вида) популяционные системы. В настоящее время на территории России выделяется 7 типов таких систем, среди которых 3 «чистых» и 4 смешанных (Лада, 1995; Lada et al., 1995; Боркин, 1999).
На территории Раифы обнаружены 4 типа популяционных систем, два из которых смешанные:
R-тип, или «чистые» популяции озерной лягушки отмечены на озерах Ильинское и Белое, а также в долинах рек Сер-Булак и Сумка. Последняя впадает в Волгу; считается, что именно с Волги по Сумке озерная лягушка проникла на территорию Раифы (Корчагина, Гаранин, 1996);
L-тип, или «чистые» популяции прудовой лягушки отмечены на озерах Илантово, Гнилое и Линево. На территории Раифского участка заповедника и его охранной зоны это самый обычный вид зеленых лягушек (Корчагина, Гаранин, 1996).
L-E-тип отмечен только на оз. Круглое. Считается самым распространенным типом смешанной популяционной системы на территории Волжского бассейна, где отмечаются гибриды (Лада, 1995; Lada et al., 1995; Боркин, 1998; Борисовский, 2001; Borkin et al., 2002; Боркин и др., 2003);
R-E-L-тип отмечен на озере Раифское, где характерна биотопическая дифференциация всех трех видов. На территории Волжского бассейна эта популяционная система распространена достаточно широко, и характерна для крупных водоемов (Лада, 1995; Lada et al., 1995; Борисовский и др., 2001; Borkin et al., 2002; Боркин и др., 2003).
Об охране зеленых лягушек
В настоящее время европейский комплекс Rana esculenta привлекает к себе внимание не только как интересный объект исследований. Изучение его структуры ставит новые вопросы в плане сохранения биологического разнообразия. Это связано, в первую очередь, с новыми данными о распространении съедобной лягушки на восточной периферии ареала. Недостаточные знания по экологии и численности этого вида дают основания для включения его в региональные Красные книги (Борисовский, 2001).
Наши материалы являются одними из первых достоверных данных о распространении всех трех видов зеленых лягушек на относительно небольшой территории Татарстана. Недостаточная изученность комплекса на всей территории республики, а также географически периферийное положение R. kl. esculenta дают основания рекомендовать этот вид для включения во второе издание Красной книги Республики Татарстан.
Авторы выражают признательность сотрудникам Волжско-Камского заповедника за поддержку в проведение исследований, а также доценту КГУ В.И. Гаранину за консультации и ценные замечания при обсуждении работы.
Работа выполнена при финансовой поддержке РФФИ (проект № 02-04-49631), ФЦП «Интеграция» (Э-0121) и АН РТ (НИОКР № 09-9.4-193).
Литература
Банников А.Г., Даревский И.С., Ищенко В.Г., Рустамов А.К., Щербак Н.Н. Определитель земноводных и пресмыкающихся фауны СССР. М., 1977. 415 с.
Борисовский А.Г. Земноводные – Amphibia // Красная книга Удмуртской Республики. Животные. Ижевск, 2001. С. 66-69.
Борисовский А.Г., Боркин Л.Я., Литвинчук С.Н., Розанов Ю.М. Распространение зеленых лягушек (комплекс Rana esculenta) в Удмуртии // Вестн. Удм. ун-та. Биол. Ижевск, 2001. № 5. С. 51-63.
Борисовский А.Г., Боркин Л.Я., Литвинчук С.Н., Розанов Ю.М. Морфометрическая характеристика зеленых лягушек (комплекс Rana esculenta) Удмуртии // Вестн. Удм. ун-та. Биол. Ижевск, 2000. № 5. С. 70-75.
Боркин Л.Я. Отряд Бесхвостые // Ананьева Н.Б., Боркин Л.Я., Даревский И.С., Орлов Н.Л. Земноводные и пресмыкающиеся. Энциклопедия природы России. М., 1998. С. 19-174.
Боркин Л.Я. Проблема видообразования у амфибий // Методологические проблемы развития зоологии. СПб., 1999. С. 13-15.
Боркин Л.Я., Виноградов А.Е., Розанов Ю.М., Цауне И.А. Полуклональное наследование в гибридогенном комплексе Rana esculenta: доказательство методом проточной ДНК-цитометрии // Докл. АН СССР, 1987. Т. 295. № 5. С. 1261-1264.
Боркин Л.Я., Даревский И.С. Сетчатое (гибридогенное) видообразование у позвоночных // Журн. общ. биол., 1980. Т. 41. № 4. С. 485-506.
Боркин Л.Я, Литвинчук С.Н., Розанов Ю.М., Лада Г.А., Ручин А.Б., Файзулин А.И, Замалетдинов Р.И. Гибридогенный комплекс Rana esculenta: существует ли «волжский парадокс» // Третья конференция герпетологов Поволжья. Тольятти, 2003. С. 7-12.
Виноградов А.Я., Розанов Ю.М., Цауне И.А., Боркин Л.Я. Элиминация генома одного из родителей до предмейотического синтеза ДНК у гибридогенного вида Rana esculenta // Цитология, 1988. Т. 30. № 6. С. 691-698.
Галеева Д.Н., Гаранин В.И., Замалетдинов Р.И., Павлов А.В. Материалы к кадастру земноводных и пресмыкающихся Республики Татарстан // Материалы к кадастру амфибий и рептилий бассейна Средней Волги. Н. Новгород, 2002. С. 186-221.
Гаранин В.И. Земноводные и пресмыкающиеся Волжско-Камского края. М., 1983. 175 с.
Корчагина Т.А., Гаранин В.И. К эколого-морфологической дифференциации зеленых лягушек Приказанья // Актуальные проблемы герпетологии и токсикологи. Тольятти, 1996. Вып. 2. С. 28-30.
Коцержинская И.М. Применение метода комплексного вариационного анализа для идентификации лягушек Rana esculenta complex // Современная герпетология. Саратов, 2000. Вып. 1. С. 71-72.
Кузьмин С.Л. Земноводные бывшего СССР. М., 1999. 298 с.
Лада Г.А. Среднеевропейские зеленые лягушки (гибридогенный комплекс Rana esculenta): введение в проблему // Флора и фауна Черноземья. Тамбов, 1995. С. 88-109.
Павлов А.В., Замалетдинов Р.И., Солодников М.В. Возможности охраны амфибий и рептилий в Предволжье и Западном Предкамье Республики Татарстан // Актуальные проблемы герпетологии и токсинологии. Тольятти, 2001. Вып. 5. С. 74-79.
Тайсин А.С. Рельеф и воды // Волжско-Камский государственный заповедник. Казань, 1969. С. 26-51.
Тайсин А.С. Антропогенная активизация и динамики озер Приказанского района: Дис. в виде науч. докл. … канд. географ. наук. Казань, 1996. 59 с.
Таращук С.Л. К методам определения европейских зеленых лягушек Rana esculenta (Amphibia, Ranidae) // Вестник зоологии, 1985. № 3. С. 83-85.
Унковская Е.Н., Мингазова Н.М., Павлова Л.Р. Гидрологическая и гидрохимическая характеристика водоемов Раифы // Тр. Волж.-Камс. гос. природ. зап-ка. Казань, 2002. Вып. 5. С. 9-36.
Berger L. Embrional and larval development of F1 generation green frog different combinations // Acta zool. Cracov, 1967. Vol. 12. № 7. P. 123-160.
Borkin L.J., Litvinchuk S.N., Mannapova E.I., Pestov M.V., Rosanov J.M. The distribution of green frogs (Rana esculenta complex) in Nizhny Novgorod province, Central European Russia // Rus. Journal of Herpetology, 2002. Vol. 9. № 3. P. 195-208.
Garanin V.I. The distribution of amphibians in the Volga-Kama Region. // Advances in Amphibian Research in the Former Soviet Union. Sofia-Moscow, 2000. Vol. 5. P. 79-132.
Gűnther R. Die Wasserfrösche Europas (Anura-Forschlurche). Wittenberg-Lutherstadt A. Ziemsen Verlag, 1990. 288 s. (Die Neue Brehm-Bucherei. Bd. 600).
Lada G.A., Borkin L.J., Vinogradov A.E. Distribution, population system and reproductive behavior of green frog (hybridogenic Rana esculenta complex) in the Central Chernozem territory of Russia // Rus. Journal of Herpetology, 1995. Vol. 2. № 1. P. 46-57.
Pagano A., Joly P. Limits of the morphometric method for taxanomic field identification of water frog. Alytes, 1998. Vol. 16. P. 3-4.
Tűnner H.D. Das Serumeiweissbild einheimischer Wasserfrösche und Hybridcharakter von Rana esculenta // Verh. Deutsch. Zool. Ges., 1970. № 23. S. 352-358.
1 Институт экологии природных систем АН РТ
2 Зоологический институт РАН
3 Институт цитологии РАН