
Пахарукова Мария Юрьевна Изучение роли транскрипционных факторов hnf3 и рецепторов ксенобиотиков в механизме видовой специфичности действия гепатоканцерогенных аминоазокрасителей (03. 00. 15 генетика) автореферат
| Вид материала | Автореферат |
СодержаниеМатериалы и методы Содержание работы |
- Автореферат диссертации на соискание ученой степени, 297.23kb.
- «Молекулярные механизмы гормональной регуляции» Общая трудоемкость изучения дисциплины, 11.18kb.
- Автореферат диссертации на соискание ученой степени, 267.26kb.
- Наименование темы урока, 96.06kb.
- Рабочей программы дисциплины Экология растений, животных, микроорганизмов (наименование), 19.6kb.
- Генетика Генетика, 18.18kb.
- «Микология» Общая трудоемкость изучения дисциплины составляет, 15.67kb.
- Темы: общая генетика, генетика человека, медицинская генетика. Вопросы для самоподготовки., 39.67kb.
- Концепция долгосрочной целевой программы «Социальная поддержка населения России», 252.47kb.
- Генетика личностных свойств, 229.93kb.
На правах рукописи
УДК: (616.006:577.171.55:577.218)
Пахарукова Мария Юрьевна
Изучение роли транскрипционных факторов HNF3
и рецепторов ксенобиотиков в механизме
видовой специфичности действия гепатоканцерогенных аминоазокрасителей
(03.00.15 - генетика)
АВТОРЕФЕРАТ
диссертации на соискание ученой степени
кандидата биологических наук
Новосибирск
2008
Работа выполнена в лаборатории регуляции экспрессии генов Института цитологии и генетики СО РАН, Новосибирск
Научный руководитель: доктор биологических наук, профессор
Т.И. Меркулова
Официальные оппоненты: доктор медицинских наук А.Ю. Гришанова
кандидат биологических наук Н.С. Логвиненко
Ведущее учреждение: Московский Государственный Университет им. М.В. Ломоносова, Москва
Защита диссертации состоится «___» _____________2008г. На утреннем заседании диссертационного совета по защите диссертаций на соискание ученой степени кандидата наук (Д-003.011.01) в Институте цитологии и генетики СО РАН, в конференц-зале Института по адресу 630090, г. Новосибирск, 90, Проспект академика Лаврентьева, 10, т/ф (383) 3331278, e-mail: dissov@bionet.nsc.ru
С диссертацией можно ознакомиться в библиотеке Института цитологии и генетики СО РАН.
Автореферат разослан «___» _____________2008г.
Ученый секретарь
диссертационного совета,
доктор биологических наук (А.Д. Груздев)
Актуальность работы. Выяснение механизмов химического канцерогенеза является одной из серьезных фундаментальных проблем теоретической онкологии и молекулярной патофизиологии. Несмотря на почти столетнюю историю исследований, до настоящего времени не имеют объяснения многие особенности действия химических канцерогенов, такие как видовая, линейная и тканевая специфичность, отсутствие соответствия между канцерогенной и мутагенной активностью многих химических соединений, обратимость ранних стадий канцерогенеза. Выяснение причин этих явлений имеет большое значение как для понимания механизма канцерогенного процесса, так и для выявления новых кандидатных генов, ответственных за формирование фенотипа, чувствительного или устойчивого к развитию злокачественных опухолей под действием определенных химических веществ.
Ранее при исследовании механизмов видовой специфичности действия гепатоканцерогенных аминоазокрасителей 3’-метил-N,N-диметил-4-аминоазобензола (3’-МеДАБ) и 2’-3-диметил-4-аминоазобензола (орто-аминоазотолуол, ОАТ) была выявлена связь между опухолеиндуцирующим эффектом соединения и снижением ДНК-связывающей активности транскрипционных факторов HNF3 (hepatocyte nuclear factor), играющих важную роль в процессах дифференцировки и торможения пролиферации гепатоцитов. Было показано, что снижение ДНК-связывающей активности HNF3 в печени крыс происходит в ответ на введение специфичного для крыс гепатоканцерогена 3'-МеДАБ, но не канцерогенного для мышей ОАТ, и, наоборот, в печени мышей активность HNF3 снижается при введении специфичного для мышей гепатоканцерогена ОАТ, но не 3'-МеДАБ, причем это имеет место только у чувствительных к опухолеиндуцирующему действию ОАТ линий [Меркулова и др., 1998; Kropachev et al., 2001; Меркулова и др., 2003]. Однако, каким образом обеспечивается такая избирательность, остается неясным. Дальнейшие исследования показали, что влияние ОАТ и 3'-МеДАБ на HNF3 осуществляется, не напрямую, а, по-видимому, посредством некоего ядерного белка(-ов) [Меркулова и др., 2003]. Представляется вероятным, что либо видоспецифичность связана с избирательным действием этого белка-посредника на HNF3γ мыши и HNF3β крысы, либо видоспецифичность обеспечивается избирательной активацией белка-посредника под действием специфичного для данного вида канцерогена. Было выдвинуто предположение, что в качестве такого белка могут выступать рецепторы ксенобиотиков, связывающие многочисленные химические соединения и опосредующие их плейотропные эффекты на организм.
Цель и задачи исследования. Целью данного исследования было изучение механизмов, обусловливающих видовую специфичность действия гепатоканцерогенных аминоазокрасителей у мышей и крыс, а также поиск ксеносенсорных белков, способных принимать участие в передаче сигнала от ОАТ и 3’МеДАБ на HNF3.
В ходе работы были поставлены следующие задачи:
- Выяснить, связана ли видовая специфичность гепатоканцерогенов ОАТ, 3’МеДАБ с различным действием белков-посредников на транскрипционные факторы HNF3β крысы и HNF3γ мыши или же она обеспечивается избирательной активацией белка-посредника под действием специфичного для данного вида канцерогена.
- Исследовать участие белка-посредника в действии видонеспецифического гепатоканцерогена ДЭНА на HNF3 белки.
- Изучить влияние ОАТ, 3’МеДАБ и ДЭНА на ДНК-связывающую активность рецепторов ксенобиотиков: печеночного Х-рецептора (LXR), прегнанового Х рецептора (PXR) конститутивного рецептора андростанов (CAR), рецептора активаторов пролиферации пероксисом (PPAR), арилгидрокарбонового рецептора (AhR) в печени мышей и крыс.
- Выяснить, обусловлено ли увеличение ДНК-связывающей активности AhR и CAR под действием гепатоканцерогенных аминоазокрасителей изменением содержания этих рецепторов в экстрактах ядер печени.
- Изучить способность ОАТ и 3'-МеДАБ специфически связываться с конститутивным рецептором андростанов (CAR).
Научная новизна и практическая значимость. Установлено, что механизмы влияния гепатоканцерогенных аминоазокрасителей ОАТ и 3'-МеДАБ и диэтилнитрозоамина (ДЭНА) на ДНК-связывающую активность транскрипционных факторов HNF3 различны. Видоспецифичность действия аминоазокрасителей на HNF3 определяется избирательной активацией ядерного белка(-ов) под влиянием гепатоканцерогенного для данного вида вещества (3’МеДАБ для крыс, ОАТ для мышей). Механизм действия неспецифического гепатоканцерогенного соединения другого класса ДЭНА не включает белка-посредника.
Показана принципиальная возможность того, что арилгидрокарбоновый рецептор (AhR) и конститутивный рецептор андростанов (СAR) могут выполнять роль белка, опосредующего действие гепатоканцерогенных аминоазокрасителей на HNF3:
1. Показано, что избирательное увеличение ДНК-связывающей активности рецепторов ксенобиотиков AhR и СAR под действием гепатоканцерогенных аминоазокрасителей ОАТ и 3'-МеДАБ как in vivo, так и in vitro соответствует способности этих веществ снижать ДНК-связывающую активность HNF3 белков.
2. Обнаружено, что при действии видонеспецифического гепатоканцерогена ДЭНА, не активирующего белок-посредник, также не активируются CAR и AhR.
3. Показано, что увеличение ДНК-связывающей активности CAR и AhR не обусловлено изменением содержания этих белков в ядрах печени, и в случае CAR связано, по-видимому, с видоспецифическими различиями в связывании этого рецептора с ОАТ и 3'-МеДАБ.
Полученные результаты имеют большое значение для установления механизмов видоспецифичности гепатоканцерогенного действия ОАТ и 3’МеДАБ, а также способствуют выяснению молекулярно-генетических основ предрасположенности или устойчивости организма к возникновению химически индуцированных злокачественных опухолей печени. Кроме того, результаты проведенного исследования могут быть использованы для разработки новых методов тестирования химических соединений на канцерогенность.
Положения, выносимые на защиту.
1. Видоспецифичность влияния гепатоканцерогенных аминоазокрасителей на транскрипционные факторы HNF3 определяется избирательной стимуляцией белка(-ов)-посредника(-ов), который активируется у крыс гепатоканцерогенным для них 3’МеДАБ, а у мышей гепатоканцерогенным для них ОАТ. Этот белок(-ки)-посредник(-ки) неспецифически снижает ДНК-связывающую активность HNF3γ мыши и HNF3β крысы.
2. Избирательная активация рецепторов ксенобиотиков AhR и СAR под действием гепатоканцерогенных аминоазокрасителей (но не их неканцерогенных аналогов) как in vivo, так и in vitro сопряжена со способностью этих соединений снижать ДНК-связывающую активность HNF3 белков.
3. Неспецифический гепатоканцероген ДЭНА снижает ДНК-связывающую активность HNF3 в отсутствии белка-посредника и не изменяет активность AhR и СAR.
4. Увеличение ДНК-связывающей активности CAR и AhR не обусловлено изменением содержания этих белков в ядрах клеток печени под влиянием аминоазокрасителей.
5. ОАТ и 3'-МеДАБ способны конкурировать с известным лигандом CAR [3Н]-андростенолом за связывание с белками цитозоля печени мышей и крыс. ОАТ является лучшим конкурентом в цитозоле печени мышей, а 3'-МеДАБ – в цитозоле печени крыс.
6. Арилгидрокарбоновый рецептор (AhR) и конститутивный рецептор андростанов (СAR) могут выполнять роль белков, опосредующих действие гепатоканцерогенных аминоазокрасителей на HNF3.
Апробация работы. Основные результаты работы были представлены на II съезде общества клеточной биологии совместно с Юбилейной конференцией, посвященной 50-летию института цитологии РАН (Санкт- Петербург, 2007.), международной конференции «Chemical and biological problems of Proteomics» (Новосибирск, 2004), VIII Дальневосточной школе-конференции по актуальным проблемам химии и биологии (Владивосток, 2004), всероссийской конференции с международным участием «Молекулярная онкология» (Новосибирск, 2008).
Публикации по теме диссертации. По материалам диссертации опубликовано 5 работ.
Структура и объем работы. Диссертация состоит из введения, обзора литературы, описания использованных материалов и методов, изложения и обсуждения собственных экспериментальных данных, заключения, выводов и списка литературы (204 наименования). Работа изложена на 122 страницах машинописного текста и содержит 25 рисунков.
^ МАТЕРИАЛЫ И МЕТОДЫ
Работа выполнена на самцах крыс весом 200г Wistar и 3-х месячных самцах мышей линии A/Не разведения вивария ИЦиГ СО РАН. В экспериментах in vivo ОАТ, 3'-МеДАБ, 4'-МеДАБ растворяли в оливковом масле, ДЭНА в физиологическом растворе и вводили внутрибрюшинно по 1 мл на 100г массы тела животных до конечной концентрации 1мМ [Каледин, Захарова, 1984]. Контрольным животным вводили растворитель. В опытах in vitro канцерогены добавляли в объеме не более 1% от объема реакционной смеси: до концентрации 0,1 мМ ОАТ, 3'-МеДАБ, 4'-МеДАБ в виде растворов в ДМСО (диметилсульфоксид), ДЭНА - в физиологическом растворе в гомогенат или ядерный экстракт [Меркулова и др., 2003]. Экстракты ядер получали согласно методу Горски-Шапиро (Gorsky-Shapiro) [Меркулова и др., 2003]. ДНК-связывающую активность факторов транскрипции оценивали методом задержки в геле [Kropachev et al., 2001] во фракции I, осаждаемой при концентрации сульфата аммония 0,32 г/мл, либо в суммарном экстракте ядер, полученном при 0,46 г/мл сульфата аммония. При проведении смешений фракций выделяли фракции I и II ядерного экстракта последовательным осаждением белков при концентрации 0,32 и 0,46 г/мл сульфата аммония. Двуцепочечные олигонуклеотиды, соответствующие известным сайтам связывания факторов транскрипции, и используемые в качестве ДНК-проб, были синтезированы В.Ф. Кобзевым (ИЦиГ СО РАН). Денситометрию радиоавтографов проводили в программе Gel Pro Analyzer 4.0. Содержание транскрипционных факторов в ядерном экстракте оценивали методом Вестерн-блот-гибридизации. Для визуализации результатов использовали ECL–систему “Amersham”. [3H]-андростенол (5α-андрост-16-ен-3α-ол) с удельной активностью 20-30 Кюри/мМ был получен методом тритиевого обмена Романовой И.В. (ХБиФМ СО РАН). Получение цитозоля клеток печени и изучение связывания [3H]-андростенола с цитозолем проводили по методу описанному ранее [Kropachev et al., 2001]. Статистическую обработку результатов проводили, используя t-критерий Съюдента в программе STATISTICA 5.5.
^ СОДЕРЖАНИЕ РАБОТЫ
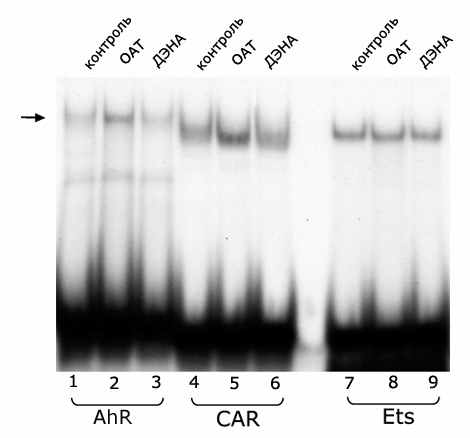
Специфичность действия белков-посредников в механизме влияния ОАТ, 3’МеДАБ на транскрипционные факторы семейства HNF3. Ранее было показано [Kropachev et al., 2001; Меркулова и др., 2003], что, во-первых, в печени мышей и крыс HNF3 ДНК-связывающая активность обеспечивают различные HNF3 белки (HNF3β у крыс и HNF3γ у мышей); а во-вторых, существует некий ядерный белок-посредник, необходимый для снижения ДНК-связывающей активности HNF3 при действии гепатоканцерогенных аминоахокрасителей. В ходе сульфатаммонийного фракционирования при выделении экстракта ядер HNF3 оказывается во фракции I, осаждаемой при концентрации сульфата аммония 0,32 г/мл, а белок-посредник – во фракции II, получаемой при 0,46 г/мл. Это дало возможность получить последовательным осаждением из экстракта ядер печени белковые препараты, содержащие HNF3 и посредник, от контрольных и обработанных гепатоканцерогенами животных. Была разработана схема смешений этих препаратов, позволяющая оценить специфичность действия белков-посредников на HNF3. Для этого смешивали фракцию I от контрольных крыс, содержащую HNF3β, с фракцией II мышей, обработанных ОАТ (Рис. 1, дорожка 6) и фракцией II крыс, обработанных 3’МеДАБ (дорожка 8); а также объединяли фракцию I контрольных мышей, содержащую HNF3γ, с фракцией II крыс, обработанных 3’МеДАБ (дорожка 4) и фракцией II мышей, обработанных ОАТ (дорожка 2).
Рис. 1. ДНК-связывающая активность HNF3 при смешивании фракций I и II экстракта ядер печени контрольных и обработанных канцерогенами мышей и крыс.
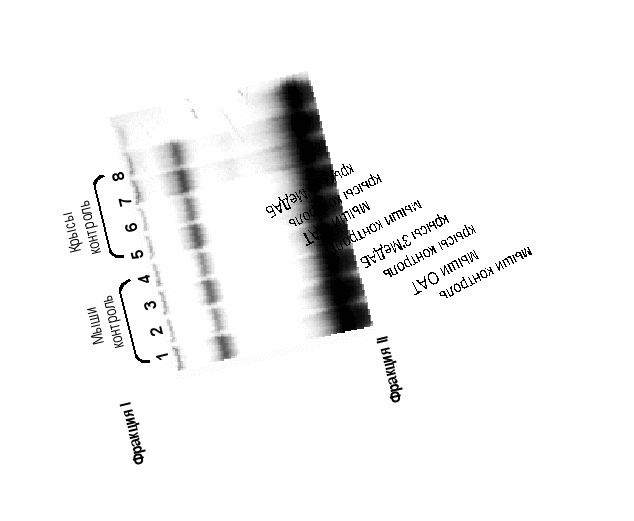
Представлен один из трех независимых экспериментов (в каждый эксперимент брали по 2 мыши и 1-2 крысы).
Оказалось, что белок-посредник мыши, активированный ОАТ, одинаково снижает ДНК-связывающую активность HNF3β крыс и HNF3γ мышей, а белок-посредник крысы, активированный 3МеДАЬ, одинаково действует на HNF3β крыс и HNF3γ мышей (Рис. 1), т.е. белок-посредник действует неспецифично по отношению к разным HNF3 белкам.
И
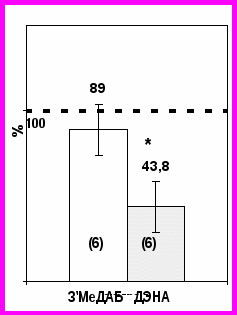 сследование участия белка-посредника в действии ДЭНА. Чтобы исследовать необходимость белка-посредника в действии видонеспецифичного ДЭНА, канцероген добавляли непосредствено к фракции I, не содержащей белка-посредника; для контроля во фракцию I вводили 3'-МеДАБ.
сследование участия белка-посредника в действии ДЭНА. Чтобы исследовать необходимость белка-посредника в действии видонеспецифичного ДЭНА, канцероген добавляли непосредствено к фракции I, не содержащей белка-посредника; для контроля во фракцию I вводили 3'-МеДАБ. Рис. 2. ДНК-связывающая активность HNF3 (в относительных единицах) при добавлении 3'МеДАБ и ДЭНА во фракцию I ядерного экстракта печени крыс.
Результаты представлены в % от интенсивности сигнала в полосе задержки при инкубации с экстрактом от контрольных животных (принята за 100% в каждой электрофореграмме). В скобках указано число экспериментов. В каждый эксперимент брали по одной-две крысы. * - достоверность отличий значений по сравнению с контрольным уровнем составила 99%.
В результате сканирования и обсчета радиоавтографов оказалось, что ДЭНА, в отличие от 3'-МеДАБ, снижает активность HNF3 при добавлении во фракцию I (Рис. 2). Следовательно, для видонеспецифичного ДЭНА посредник не нужен, поскольку фракция I не содержит этого белка, необходимого для действия 3’МеДАБ. Прямо или опосредованно ДЭНА осуществляет свое действие на HNF3 остается неизвестным, однако полученные нами данные показывают, что видонеспецифичный гепатоканцероген ДЭНА реализует иной механизм воздействия на ДНК связывающую активность HNF3, чем видоспецифичные ОАТ и 3'-МеДАБ.
Влияние ОАТ, 3’МеДАБ и ДЭНА на рецепторы ксенобиотиков PPAR, CAR, PXR, LXR, AhR. Представлялось наиболее вероятным, что белок-посредник в действии гепатоканцерогенных аминоазокрасителей может относиться к рецепторам ксенобиотиков. Известно, что эти рецепторы связывают широкий круг соединений различной структуры и участвуют во множестве регуляторных процессов, включая регуляцию клеточного цикла и индукцию опухолей. Поэтому было изучено действие ОАТ и 3’МеДАБ на ДНК-связывающую активность PPAR, CAR, PXR, LXR, AhR методом задержки в геле олигонуклеотидов, соответствующих их специфическим сайтам связывания. Видно (Рис. 3), что под действием и ОАТ и 3’МеДАБ происходит увеличение активности практически всех рецепторов, за исключением PPAR. Однако нас интересовал рецептор, который обнаружил бы избирательность в активации видоспецифическими канцерогенами, и такую избирательность в активации проявили два рецептора - конститутивный рецептор андростанов и арилгидрокарбоновый рецептор.
Рис. 3. ДНК-связывающая активность CAR, Ah-рецептора (AhR), PXR, LXR, PPAR, Ets в суммарной фракции экстракта ядер печени мышей (А) и крыс (Б) в контроле и после введения животным азокрасителей.
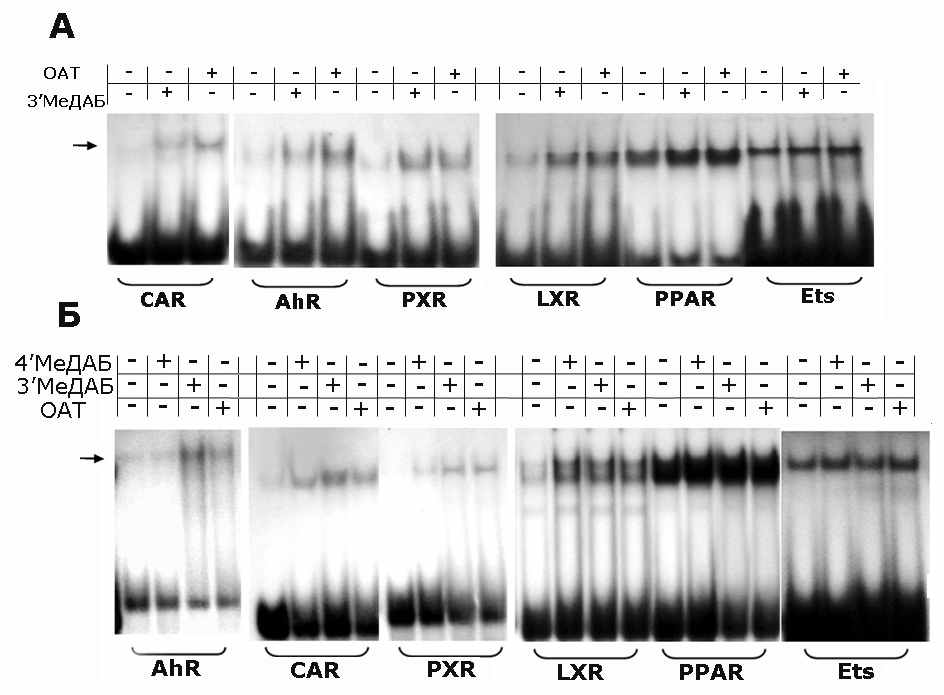
Представлены типичные результаты одного из 5 экспериментов (мыши) и 4 экспериментов (крысы). В каждый эксперимент брали по 1 крысе и 2 мыши.
Рис. 4. Изменение ДНК-связывающей активности арилгидрокарбонового рецептора (AhR) и конститутивного рецептора андростанов (CAR) в суммарной фракции ядер печени мышей и крыс под действием ОАТ и 3’МеДАБ в относительных единицах
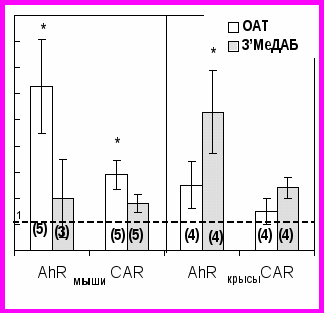 по результатам количественной денситометрии радиоавтографов (Рис. 3), за единицу принята интенсивность сигнала в полосе задержки при инкубации с экстрактом ядер печени животных контрольной группы. В скобках указано число экспериментов. В каждый эксперимент брали по две мыши или по одной крысе. * - достоверность отличий данных “3’МеДАБ” и “ОАТ” составила 95%, при использовании t-критерия Стьюдента.
по результатам количественной денситометрии радиоавтографов (Рис. 3), за единицу принята интенсивность сигнала в полосе задержки при инкубации с экстрактом ядер печени животных контрольной группы. В скобках указано число экспериментов. В каждый эксперимент брали по две мыши или по одной крысе. * - достоверность отличий данных “3’МеДАБ” и “ОАТ” составила 95%, при использовании t-критерия Стьюдента.Результаты количественной обработки радиоавтографов экспериментов по задержке в геле (Рис. 4) показывают, что в ответ на ОАТ у мышей активность AhR увеличивалась в 6,3 раза, а CAR - в 3 раза по сравнению с их исходным уровнем, тогда как в ответ на неканцерогенный для этих животных 3’МеДАБ всего лишь в 2 раза. В печени крыс в ответ на гепатоканцерогенный для них 3’МеДАБ активность AhR увеличивалась в 6,3 раза, а CAR - в 3 раза по сравнению с их исходным уровнем, тогда как в ответ на неканцерогенный ОАТ для этих животных активность AhR всего лишь в 2 раза, CAR – недостоверно изменялась.
Для идентификации ДНК-связывающей активности AhR и CAR были использованы специфические антитела к AhR и CAR. Добавление антител к CAR приводило к исчезновению полосы, соответствующей комплексу белок-зонд, добавление разных препаратов антител к AhR приводило либо к исчезновению соответствующих полос, либо к их супершифту т.е. комплекс АТ-ТФ-зонд демонстрирует меньшую подвижность, чем в отсутствии антител.
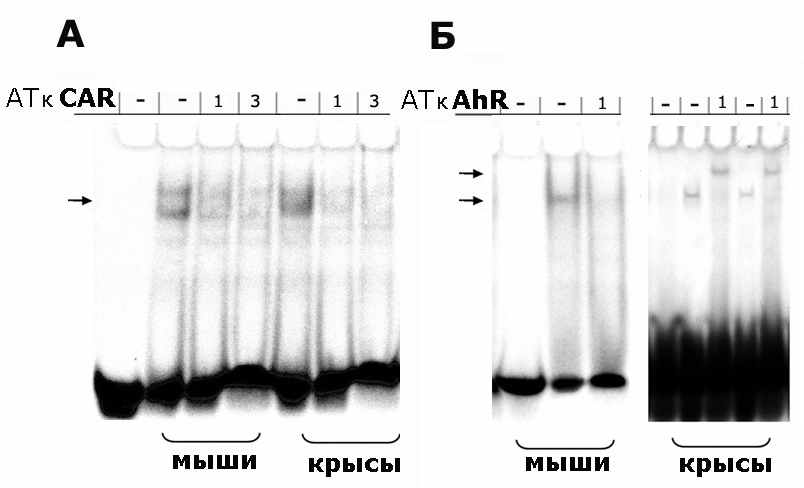
Рис. 5. Идентификация белков с помощью антител (АТ) в ДНК-белковых комплексах, образованных белками ядерного экстракта и ДНК-зондами CAR (А) и AhR (Б). Экстракты ядер прединкубированы в присутствии антител и при добавлении 1% желатина. Использовали АТ к CAR (SantaCruz Biotech. Inc., USA), в случае мышей использовали АТ к AhR (SantaCruz Biotech. Inc., USA), а в случае крыс – АТ к AhR (AbCam, USA).
Введение ДЭНА, вызывающего развитие опухолей и у крыс и у мышей, также сопровождается снижением активности HNF3, также как и введение видоспецифических аминоазокрасителей [Меркулова и др., 2003]. Однако, как мы показали, механизм действия ДЭНА не требует присутствия белка-посредника (Рис. 2). Поэтому можно было предполагать, что если один из рецепторов - потенциальный посредник, обеспечивающий видоспецифичность действия азокрасителей, то он не должен активироваться ДЭНА.
Рис. 6. ДНК-связывающая активность CAR, AhR и Ets в экстрактах ядер печени мышей A/He в контроле и при введении ОАТ и ДЭНА.
 Приведен типичный радиоавтограф одного из 3 независимых экспериментов.
Приведен типичный радиоавтограф одного из 3 независимых экспериментов.Оказалось, что в ответ на ДЭНА (Рис. 6) активации CAR и AhR не наблюдается. Эти результаты хорошо сочетаются с данными о том (Рис. 2), что механизм действия ДЭНА не включает активации белка-посредника, необходимого для действия ОАТ и 3’МеДАБ.
ДНК-связывающая активность белка определяется как количеством молекул этого белка, так и их состоянием. Поэтому была проведена оценка влияния гепатоканцерогенных аминоазокрасителей на содержание белков AhR, CAR методом Вестерн-блот-гибридизации (Рис. 7).
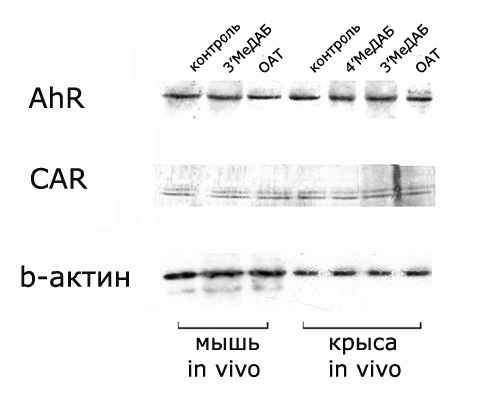 Рис. 7. Вестерн-блот-гибридизация с антителами к AhR и CAR экстрактов ядер клеток печени мышей и крыс до и после обработки аминоазокрасителями ОАТ и 3’МеДАБ.
Рис. 7. Вестерн-блот-гибридизация с антителами к AhR и CAR экстрактов ядер клеток печени мышей и крыс до и после обработки аминоазокрасителями ОАТ и 3’МеДАБ.Приведен типичный результат одного из 3 независимых экспериментов. Параллельно оценивали содержание белка β-актина.
Согласно ранее полученным данным ДНК-связывающая активность AhR и CAR под действием специфичного для данного вида гепатоканцерогена увеличивалась в 5-6 раз и в 3 раза соответственно (Рис. 5). По результатам Вестерн-блот-гибридизации таких многократных изменений содержания CAR и AhR после обработки гепатоканцерогенами не выявлено (Рис. 8). Следовательно, наблюдаемый эффект увеличения ДНК-связывающей активности CAR и AhR под действием ОАТ и 3’МеДАБ не объясняется изменением содержания этих белков в ядерных экстрактах. По-видимому, это происходит за счет изменения их состояния, например, в результате связывания ОАТ и 3’МеДАБ. Известно, что аминоазокрасители являются активаторами AhR человека [Kato et al., 2002], крыс и мышей [Mikhailova et al., 2005; Yang M et al., 1997], что позволяет предполагать, что ОАТ и 3’МеДАБ могут выступать в роли лигандов AhR. Также нельзя исключить и CAR из числа регуляторных белков, связывающих эти соединения.
Изучение связывания гепатоканцерогенных аминоазокрасителей с CAR. Для изучения способности ОАТ и 3’МеДАБ специфически взаимодействовать с конститутивным рецептором андростанов были проведены эксперименты по конкуренции этих канцерогенов с известным лигандом CAR андростенолом (5α-андрост-16-ен-3α-ол) [Forman et al., 1998, Vincent et al., 2005] за связывание с белками цитозоля печени мышей и крыс (Рис.9).
Рис. 9. Связывание 50 мкМ [3Н] – андростенола с цитозолем печени крыс (А) и мышей (Б) в присутствии 25-200(250) кратных избытков немеченого андростенола (кривая 1), 3’МеДАБ (кривая 3), ОАТ (кривая 2), клофибрата (кривая 4), прогестерона (кривая 5).
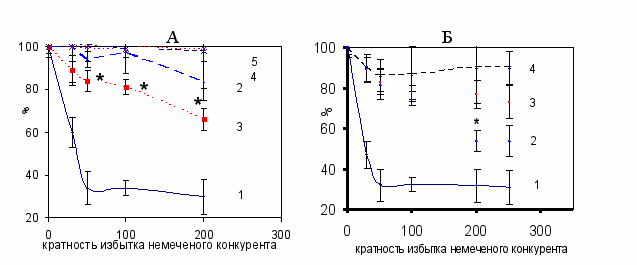
Представлены результаты 3-5 независимых экспериментов. * - разница значений “3’МеДАБ” и “ОАТ” составила 95%.
В качестве негативного контроля были выбраны прогестерон и клофибрат (высокоспецифичный лиганд для PPAR), которые не конкурировали с андростенолом (Рис. 9). Достоверные результаты по конкуренции ОАТ и 3’МеДАБ были получены при использовании этих веществ в 50-200 кратном избытке по сравнению с меченым андростенолом. Соответствующие этим избыткам концентрации находятся в диапазоне доз, в которых эти вещества вызывают развитие опухолей печени при введении животным (1 мМ), и доз, которые были использованы в экспериментах in vitro по оценке влияния гепатоканцерогенов на ДНК-связывающую активность HNF3 (0,1 мМ) [Каледин и Захарова, 1984; Меркулова и др., 2003]. Этот результат свидетельствует о том, что аминоазокрасители являются низкоаффинными лигандами CAR. Важно отметить тот факт, что в цитозоле печени крыс 3’МеДАБ конкурировал с [3Н]-андростенолом сильнее чем ОАТ, и обратная картина наблюдалась в экспериментах с цитозолем печени мышей – ОАТ сильнее конкурировал, чем 3’МеДАБ (Рис. 9). Таким образом, присутствующий в цитозоле печени мышей и крыс белок лучше связывал «свой» канцероген (ОАТ для мышей и 3’МеДАБ для крыс), чем «чужой» канцероген (3’МеДАБ для мышей и ОАТ для крыс). Эти результаты полностью соответствуют различиям в активации CAR под действием аминоазокрасителей (у мышей ОАТ сильнее увеличивает ДНК-связывающую активность CAR, а у крыс - 3’МеДАБ (Рис. 5)). Итак, вероятно гепатоканцерогенные аминоазокрасители могут связываться с CAR, но с различной эффективностью в зависимости от происхождения белка.
В заключение важно отметить, что получены факты, убедительно свидетельствующие о том, что роль белка, который необходим для угнетения функции HNF3 под влиянием гепатоканцерогенных ОАТ и 3’МеДАБ способны выполнять рецепторы ксенобиотиков CAR и AhR. Во-первых, CAR и AhR активируются избирательным образом под действием аминоазокрасителей: у мышей, чувствительных к гепатоканцерогенному действию ОАТ - строго под действием гепатоканцерогенного ОАТ, но не 3’МеДАБ; а у крыс – под действием 3’МеДАБ, но не ОАТ. Во-вторых, мы показали, что ОАТ и 3'МеДАБ способны конкурировать с лигандом CAR – андростенолом, и, по-видимому, являться его низкоаффинными лигандами. Из данных литературы известно, что ОАТ и 3'МеДАБ активируют экспрессию генов-мишеней AhR, и, по-видимому, являются лигандами AhR. Кроме того, нами показано, что неспецифический гепатоканцероген ДЭНА, который снижает активность HNF3 без участия белка-посредника, также и не активирует AhR и CAR. Оба эти рецептора связывают широкий круг химических веществ и участвуют в регуляции многих процессов в организме млекопитающих, в том числе дифференцировки клеток и клеточного роста, индукции ферментов системы детоксикации ксенобиотиков [Carlson&Perdew, 2002; Barouki et al., 2007]. Возможно, именно эти белки играют ключевую роль в распознавании аминоазокрасителей и обеспечении их видоспецифических гепатоканцерогенных эффектов. Детали механизма передачи сигнала от AhR и CAR на HNF3 пока неизвестны, возможно, что AhR и CAR взаимодействуют с каким-либо белком, который также связывается с HNF3 и приводит к снижению его ДНК-связывающих свойств. Известно также, что рецепторы AhR и CAR активируют ряд протеинкиназ, фосфатаз, которые могут оказывать модифицирующее воздействие на HNF3, меняя его ДНК-связывающие свойства. Какой из этих механизмов вовлечен в передачу сигнала от CAR AhR на HNF3, пока не ясно.
Выводы
- Показано, что белок(-ки)-посредник(ки) действия гепатоканцерогенных аминоазокрасителей на транскрипционные факторы HNF3 одинаково снижают ДНК-связывающую активность HNF3γ мыши и HNF3β крысы. Видоспецифичность действия этих соединений обусловлена избирательной активацией белка(-ов)-посредника(-ов) у крыс под действием гепатоканцерогенного для них 3’МеДАБ, а у мышей - гепатоканцерогенного для них ОАТ.
- Выявлена связь между избирательной активацией рецепторов ксенобиотиков AhR и СAR и снижением HNF3 ДНК-связывающей активности под действием гепатоканцерогенных аминоазокрасителей, что наблюдается только у чувствительных к этим соединениям животных (под действием ОАТ у мышей, и под действием 3’МеДАБ у крыс).
- Показано, что видонеспецифичный гепатоканцероген ДЭНА снижает ДНК-связывающую активность HNF3 по иному механизму, чем видоспецифичные ОАТ и 3'-МеДАБ, не включающему белка-посредника. В ответ на ДЭНА активации CAR и AhR не происходит.
- Установлено, что увеличение ДНК-связывающей активности CAR и AhR под действием ОАТ и 3’МеДАБ не вызвано увеличением содержания этих белков в ядрах клеток печени.
- Показано, что ОАТ и 3'-МеДАБ конкурируют с [3Н]-андростенолом (известным лигандом CAR) за связывание с белками цитозоля клеток печени. При этом 3’МеДАБ конкурирует за связывание с белками цитозоля печени крыс лучше, чем ОАТ, а ОАТ является более сильным конкурентом, чем 3’МеДАБ в цитозоле печени мышей.
- Совокупность полученных данных позволяет предполагать, что AhR и СAR могут выполнять роль белков, опосредующих действие гепатоканцерогенных аминоазокрасителей на HNF3.
Основные результаты работы содержатся в следующих публикациях:
- М.Ю. Пахарукова, М.А. Сметанина, В.И. Каледин, В.Ф. Кобзев, И.В. Романова, Т.И. Меркулова. Активация конститутивного рецептора андростанов под действием гепатоканцерогенных аминоазокрасителей в печени мышей и крыс// Бюллетень экспериментальной биологии и медицины. Т. 144. 2007. № 9. С. 313-316.
- М.Ю. Пахарукова, М.А. Сметанина, В.И. Каледин, И.В. Романова, Т.И. Меркулова. Изучение роли ядерных рецепторов и Ah-рецептора в механизме действия гепатоканцерогенных азокрасителей// Цитология. Т. 49. 2007. №9. С. 781-782.
- Merkulova T.I., Kropachev K.Y., Timofeeva O.A., Vasiliev G.V., Levashova Z.V., Ilnitskaya S.I., Kobzev V.F., Pakharukova M.Y., Bryzgalov L.O., Kaledin V.I. Species-specific effects of hepatocarcinogens 3'-methyl-4-dimethyl-aminoazobenzene and ortho-aminoazotoluene in mouse and rat liver// Mol. Carcinogenesis. 44. 2005. № 4. Р. 223-232.
- К.Ю. Кропачев, М.Ю. Пахарукова, Л.О. Брызгалов, В.И. Каледин, В.Ф. Кобзев, Т.И. Меркулова. Неизвестный ядерный белок, снижающий активность HNF3, обеспечивает видоспецифичность действия гепатоканцерогенных аминоазокрасителей// Доклады Академии Наук. Т. 397. 2004. № 5. С. 694-696.
- T.I. Merkulova, V.I. Kaledin, K.Yu. Kropachev, G.V. Vasiliev, V.F. Kobzev, M.Yu. Pakharukova, L.O. Bryzgalov// Abstract book and programme of International conference «Chemical and biological problems of Proteomics». Novosibirsk. Russia. 2004.