Ученье свет, а неученье тьма народная мудрость
| Вид материала | Документы |
- Ученье свет, а неученье тьма народная мудрость, 3885.89kb.
- Каер Жанна Алексеевна моу вознесенская сош 10 класс сочинение, 63.71kb.
- «Ученье – свет, а неученье – тьма», 41.62kb.
- Внеклассное мероприятие по литературному чтению Долмашкина А. В.,Моу сош №44, 60.21kb.
- Задание Придумайте название для своей команды. Составьте небольшой рассказ о своей, 338.38kb.
- Ученье свет, а неученье жди проверки, 103.98kb.
- City of ember официальный дистрибьютор в России кинокомпания «Вест», 1964.97kb.
- Анхель де Куатьэ Тайна печатей (книги 1-6), 6473.48kb.
- Лекция 1-2, 1001.75kb.
- Говорят, нет дыма без огня, а, как известно, народная мудрость редко ошибается., 100.75kb.
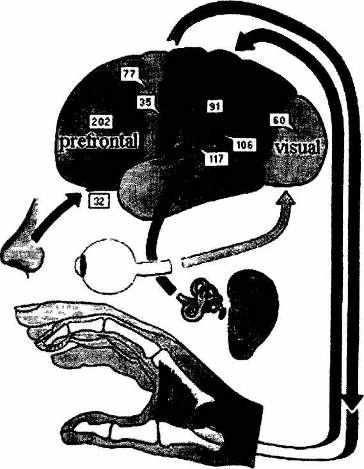
На рис. 2.14 показано зарегистрированное с помощью этого метода изменение активности различных участков коры испытуемого, пытающегося вспомнить название столицы Южной Кореи. Особенно сильно выраженный участок активации в префронтальных областях левого полушария может интерпретироваться как типичное проявление интенции на воспроизведение информации из безличной семантической памяти (при попытках припоминания автобиографических сведений активация наблюдается в правых префронтальных структурах — см. 5.3.2). Одновременная активация заднетеменных областей коры свидетельствует о том, что эта задача, по-видимому, решается при помощи фоновых координации, связанных с мысленным просмотром пространственных репрезентаций, или «когнитивных карт» (см. 6.3.1).
Как мы отмечали выше, магнитные поля могут использоваться не только для диагностики, но и для активного изменения динамики физиологических процессов. Для этого применяются мощные магнитные воздействия, которые ведут к кратковременному нарушению работы определенных участков мозга. Сам этот метод получил название трансчерепной магнитной стимуляции, или Τ MC {transcranial magnetic stimulation, TMS). Один из интереснейших результатов, полученных с помощью данного метода, явно противоречит предсказаниям и духу концепции модулярности Фодора (см. 2.3.2). Оказалось, что затылочные (то есть с анатомической точки зрения зрительные) отделы коры слепых испытуемых постепенно меняют свою специализацию и принимают на себя часть функций по переработке тактильной информации. Они не только систематически активируются в случае тактильных за-
149
дач, но их временное «выключение» ведет к возникновению ошибок узнавания воспринимаемых на ощупь объектов, например, при чтении шрифта Брайля.
Одним из самых мощных методов современной когнитивной нейронауки является магнитоэнцефалография (МЭГ). В ней применяется регистрация магнитных полей, сопровождающих активность нейронов. Поскольку эти эффекты примерно в 100 млн. раз слабее фонового магнитного поля Земли, их регистрация чрезвычайно трудоемка и требует использования специальных сверхпроводниковых детекторов (superconducting quantum interference devices — SQUIDs). Это самый быстрый из всех неинвазивных методов, конкурирующий даже с микроэлектродным отведением. МЭГ с успехом используется для уточнения функции отдельных регионов мозга, а также для описания временной развертки активации. Так, с ее помощью было установлено, что зрительные поля V5, участвующие в переработке информации о движении, включаются в работу через 20—30 мс после активации первичных зрительных полей VI (Andersen et al., 1997). К сожалению, МЭГ имеет слабое пространственное разрешение, поэтому обычно ее применяют в сочетании с фМРТ. Для проведения подобных исследований нужно участие междисциплинарного коллектива в составе физиков, психологов, нейрофизиологов и медиков, владеющих навыками работы с дорогостоящей физической аппаратурой, способных интерпретировать получаемые данные и объяснять их друг другу20.
Стоимость необходимой технической инфраструктуры и уникальная квалификация сотрудников — не единственная проблема мозгового картирования. Большинство этих методов основано на регистрации метаболических процессов, лишь сопутствующих собственно нейрофизиологической активности. Характер этой связи не всегда понятен в деталях. Далее, получаемая информация, давая трехмерную модель активированных областей, обычно ничего не говорит о взаимоотношениях между этими областями в процессе решения задачи. В этом смысле известные преимущества имеет классический метод ЭЭГ, так как он дает возможность определять корреляцию (когерентность) колебаний электрических потенциалов в различных областях поверхности коры и, таким образом, судить о типе взаимоотношений этих областей. Наконец, серьезные сомнения начинает вызывать так называемая процедура вычитания, с которой практически всегда связано использование мозгового картирования
2
 0 В настоящее время апробируется так называемая оптическая томография, при которой применяется близкое к инфракрасной зоне спектра световое облучение {Near Infra-Red Spectroscopy, Ν 1RS). Хотя значительная часть фотонов поглощается при этом тканями кожи, черепа и мозга, некоторая часть выходит наружу, что позволяет судить о состоянии активации нейронных структур и степени кровенаполнения. По-видимому, речь идет о создании аналога фМРТ, но более дешевого и удобного в применении. Ведется работа и над простыми акустическими методами мозгового картирования, использующими для оп-150 ределения скорости кровотока эффект Допплера.
0 В настоящее время апробируется так называемая оптическая томография, при которой применяется близкое к инфракрасной зоне спектра световое облучение {Near Infra-Red Spectroscopy, Ν 1RS). Хотя значительная часть фотонов поглощается при этом тканями кожи, черепа и мозга, некоторая часть выходит наружу, что позволяет судить о состоянии активации нейронных структур и степени кровенаполнения. По-видимому, речь идет о создании аналога фМРТ, но более дешевого и удобного в применении. Ведется работа и над простыми акустическими методами мозгового картирования, использующими для оп-150 ределения скорости кровотока эффект Допплера.в экспериментальных исследованиях. При этом из картины активации мозга в экспериментальном условии вычитается картина активации в контрольном или некотором другом референтном условии.
Так, чтобы узнать, в чем состоит особенность нейрофизиологических механизмов семантической обработки материала по сравнению с перцептивной обработкой, из результирующего «образа активации» в задаче на семантическую категоризацию вычитается «образ активации» в задаче на анализ перцептивных признаков материала. При этом, естественно, все области мозга, которые были примерно одинаково активированы в обеих задачах, выпадают из рассмотрения. Но что если активированные области — это не просто мозаика независимых друг от друга модулей и целое в работе мозга, как подчеркивали гештальтпсихологи, всегда «больше суммы своих частей»? Тогда процедура вычитания может не только не помочь, но даже осложнить интерпретацию полученных данных. В формальном отношении это старая проблема, которая впервые возникла в 19-м веке в связи с дискуссиями, развернувшимися в связи с методом вычитания Дондерса (см. 1.2.1 и 2.2.3). Наметившееся решение применительно к методикам трехмерного мозгового картирования заключается в использовании статистических процедур факторного анализа, выявляющих информацию о взаимодействии (обычно в форме корреляций) процессов активации различных структур мозга.
Перед тем как перейти к рассмотрению нового поколения теоретических моделей когнитивной психологии, необходимо упомянуть еще одну группу методов, использование которых становится обязательным атрибутом все большего числа исследований. Речь идет о регистрации движений глаз, основанной на быстрой обработке видеоизображений. Хотя история анализа движений глаз насчитывает более 100 лет, значительная часть ранних работ была направлена на классификацию типов движений глаз и на описание их биомеханических характеристик. Безусловно релевантными для психологии были лишь исследования чтения и зрительного поиска. Они позволили описать характерный узор очень быстрых саккадических движений и относительно неподвижных фиксаций, а также выявить то обстоятельствог что глазодвигательная активность обычно несколько опережает все формы отчета (включая интроспекцию!) о выполняемой деятельности (см. 3.4.1).
В конце 1960-х годов широкую международную известность получили работы отечественных исследователей, прежде всего биофизика А.Л. Ярбуса (1965). Он продемонстрировал выраженную зависимость траектории движений глаз при рассматривании сложных осмысленных изображений от стоящей перед субъектом задачи. Насколько всеобщим было восхищение результатами Ярбуса, настолько незначительным оказалось желание их повторить. Дело в том, что его методика была основана на использовании резиновой присоски с укрепленным на ней зеркальцем (позднее радиоантенной). Присоска ставилась непосредственно на поверхность (склеру) глаза испытуемого, голова которого жестко фиксировалась в металлической рамке. Часто испытуемый еще 151
должен был в течение эксперимента держать прикушенной специальную соединенную с рамкой и вылитую по форме его зубов пластиковую пластину. Такой эксперимент мог продолжаться не более нескольких минут, обработка же данных занимала недели и месяцы.
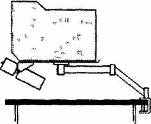
Ситуация стала быстро меняться в середине 1990-х годов, когда началось гражданское использование технологии автоматического наведения ракет по видеоизображениям цели и ее непосредственного окружения. При регистрации движений глаз роль отслеживаемой «цели» выполняют зрачки, снимаемые в инфракрасном, остающемся невидимым для испытуемого свете быстрыми (с частотой смены кадра до 500 Гц) и очень маленькими (примерно 5 мм) телекамерами. Одновременно таким же образом регистрируются движения головы. Объединение информации о положении головы и зрачков позволяет чрезвычайно быстро (с задержкой в несколько миллисекунд) определять абсолютное направление взора в окружающем пространстве. Эти новые методы могут быть совершенно бесконтактными (камеры устанавливаются только в окружении, например, монтируются над компьютерным монитором). Они также позволяют испытуемому одновременно с регистрацией в известных пределах двигать головой, взаимодействовать с техническими устройствами и общаться с другими людьми (рис. 2.15).
Значение этих методов выходит за рамки исследований собственно зрительного восприятия, поскольку направление взора служит наиболее надежным и практически мгновенным индикатором положения фокуса внимания, а следовательно, и выполняемых действий (см. 4.1.1). Кроме того, продолжительность зрительных фиксаций и общее время пребывания взора в определенной области окружения позволяют судить о характере и уровне обработки информации (Vehchkovsky, 1999). Существенно, что данные о направлении взора и окуломоторных собы-
инфракрасная видеокамера
компьютер
152
Рис. 2.15. Бесконтактная методика регистрации движений глаз А — инфракрасное освещение, выделяющее зрачки для компьютерной видеообработки и слежения, Б — возможное расположение аппаратуры (по Joos, Rotting & Vehchkovsky, 2003)
тиях, включающих саккады и моргания, обрабатываются за сотые доли секунды. Это дает возможность менять параметры ситуации прямо по ходу эксперимента, в зависимости от параметров движений глаз (gaze-contingent experiments). Более того, специальные процедуры, близкие коннекционистским моделям, делают возможным по узору саккад и фиксаций идентифицировать намерения человека, что необходимо для адаптивной поддержки операторов сложных технических систем (см. 2.1.2 и 7.4.3). Если мозговое картирование радикально изменило характер работ в области нейропсихологии, регистрации движений глаз, судя по всему, предстоит революционизировать многие практические приложения когнитивных исследований.
2.4.3 Нейробиологические модели познания
В потоке новых данных, порожденных использованием методов трехмерного картирования мозга, стало не всегда просто узнавать аналоги традиционных психологических понятий и те компоненты ранних когнитивных моделей, которые строились на базе традиционной компьютерной метафоры. Даже если пытаться и далее описывать мозг как машину, занимающуюся вычислениями, ясно, что это машина совершенно другого рода, чем все известные нам на сегодняшний день рукотворные технические устройства. Главное отличие состоит, несомненно, в эволюционном и онтогенетическом развитии, а также в обилии элементов — по некоторым данным общее число нейронов головного мозга превышает 100 миллиардов, число же их специализированных соединений, синапсов, оказывается на два-три порядка больше (см. 9.4.2). Все это ведет к массивной параллельности нейрофизиологических процессов, сочетающейся, впрочем, с определенной анатомической дискретностью и функциональной специализацией мозговых структур.
Многие авторы пытались дать описание такой глобальной специа
лизации. Одним из самых известных примеров служит схема А. Р. Лурия
(1975), в которой выделено три основных функциональных блока мозга.
Так называемый энергетический блок (стволовые отделы мозга, древняя
кора и медиобазальные отделы лобной коры) обеспечивает длительное
бодрствование и осуществление форм деятельности, требующих кон
центрации внимания. Второй блок (задние височные и теменно-заты-
лочные отделы коры) обеспечивает получение, переработку и хранение
информации. Это гностический или познавательный блок. Наконец,
третий, исполнительный блок (лобные доли) необходим для программи
рования, регуляции и контроля текущей деятельности. Взгляды Лурия
продолжают сегодня интенсивно развиваться в нейрокогнитивных мо
делях внимания, которые выявляют также различные генетические и
биохимические основания для каждого из этих функциональных блоков
(см. 4.3.3). 153
Надо сказать, что это была далеко не первая и не последняя трехуровневая классификация. Самая первая из них была предложена Джоном Хьюлинг-Джексоном, выдающимся английским неврологом конца 19-го века. Опираясь на эволюционное учение Дарвина, он выделил лобные доли коры как высшую инстанцию поведенческого контроля у человека. Кроме того, проведенный им в ряде работ анализ расстройств речи (афазий) привел его к выводу, что в реализации речевых функций принимают участие не только традиционно рассматриваемые в этом контексте структуры левого полушария (уже хорошо известные к тому времени зоны Брока и Вернике — см. 7.1.1 ), но и механизмы правого полушария, а также субкортикальные центры. Он сформулировал идеи координации активности нескольких разноуровневых механизмов в процессе решения задач, близкие понятию «функциональная система». Эти работы послужили одним из оснований для предпринятой H.A. Бернштейном 50 лет спустя попытки теоретического синтеза накопленных к середине 20-го века поведенческих и нейропсихологических данных о механизмах регуляции движений (см. 1.4.2).
Практически одновременно с Лурия американский нейрофизиолог Пол Маклин (например. McLean, 1973) предложил несколько иную концепцию тройственной организации работы мозга. Наибольший интерес в ней представляет описание среднего уровня, включающего связанную с пространственной памятью и аффективным поведением лим-бическую кору — высшую инстанцию мозга у древних млекопитающих. Лимбическая система, в свою очередь, развивается из субкортикальных базалъных ганглиев, ответственных за контроль поведения у рептилий и птиц. По мнению Маклина, агрессивность территориального поведения отдельных людей, групп и целых государств — наследие этого периода филогенеза. Хотя человек интеллектуально и покинул царство биологических закономерностей, эмоционально он не в состоянии контролировать свой, в его основе все еще «рептильный мозг» {reptilian brain)21.
Все эти авторы — Хьюлинг-Джексон, Бернштейн, Лурия и Маклин, — по сути дела, решали одну и ту же общую задачу, которая называется в геологии стратификацией, то есть выделением в общем массиве породы отдельных слоев, или уровней, относящихся к различным предысторическим эпохам (Velichkovsky & Challis, 1999). Стратификация выполняет в геологии чрезвычайно полезную роль, позволяя сразу переносить на найденный в некотором слое материал общие характеристики соответствующей эпохи, например, кембрия или триасса. Если
2
1 Речь идет о довольно смелом, критикуемом многими авторами предположении. Оно частично подтверждается современными исследованиями эволюции мозга, свидетельствующими о том, что одна из ответственных за эмоциональные реакции млекопитающих (а тем самым, и человека — см. 9.4.3) структура, так называемая миндалина (амигдала), развивается непосредственно из зачатков коры древних рептилий (Abolotz, Morales & Montiel, 154 2003).бы нам удалось построить уровнево-эволюционную модель психофизиологических процессов, то она могла бы выполнять по отношению к частным находкам и феноменам когнитивной науки аналогичную эвристическую функцию (см. 1.4.1 и 8.4.3).
Характеризуя актуальное состояние исследований, следует отметить их известную противоречивость, вызванную быстрой сменой характера объяснительных моделей. Волна технических инноваций стремительно меняет в последнее время сам облик когнитивной психологии как научной дисциплины в направлении комплексных когнитивных нейроисследований. Современные модели психологических процессов испытывают особенно сильное влияние «модулярной» нейропсихологии. При этом из-за обилия данных и противоречивости теоретических взглядов зачастую оказывается забытой более общая эволюционная и функциональная перспектива исследований. Конец 20-го века, когда многие когнитивные психологи — порой довольно неожиданно для самих себя — стали нейропсихологами и психофизиологами, оказался временем выдающихся открытий (часть из них будет обсуждаться в последующих главах), но зачастую и псевдооткрытий, в традициях так называемой френологии — популярной в начале 19-го века салонной игры на ощупывание формы черепа с целью определения бугров и впадин, предположительно связанных со степенью развития патриотизма, памяти или, например, литературных способностей.
Так, несколько лет назад одновременно в Северной Америке и Европе появились сообщения двух видных авторов об открытии с помощью ПЭТ-сканирования «центра мышления» — любое умственное усилие действительно вызывало активацию этой узколокализованной области в основании фронтальных долей. При последующем анализе этот «центр» оказался, однако, всего лишь крупным узлом питающих передние отделы коры мозга кровеносных сосудов. По сегодняшний день в профессиональной литературе обсуждается другое недавнее предположение (Tulving, 1998), согласно которому кодирование информации о различных событиях собственной биографии при запоминании (так называемая эпизодическая память — см. 5.3.2) преимущественно осуществляется фронтальными отделами левого полушария, а ее извлечение при припоминании — префронтальными структурами правого. Трудно прежде всего представить эволюционные основания для подобной специализации полушарий, ведь запоминание и припоминание обычно представляют собой моменты единого мнестического действия. Не случайно это предположение оказалось скорректированным в ходе психологических и нейропсихологических исследований памяти (см. 5.3.3).
Одновременно накапливаются новые данные о множественности механизмов и чрезвычайно широких взаимосвязях таких глобальных когнитивных функций, как, например, речь и память. В частности, в случае речи (как это предполагал уже Хьюлинг-Джексон) было установлено, что мозговая локализация основных механизмов не ограничена только левым полушарием коры, но включает также структуры правого
155
полушария и различные субкортикальные механизмы (см. 7.3.3). В результате подобного обилия взаимосвязей изучаемых структур наиболее успешными практически во всех областях когнитивных исследований сегодня оказываются нейросетевые, или коннекционистские модели.
В связи с распространением моделей, основанных на использовании искусственных нейронных сетей, следует заметить, что реальные мозговые механизмы, по-видимому, демонстрируют несколько различных принципов эволюционной организации (Striedter, 2004). Наряду с формами организации, напоминающими относительно автономные «модули» («ядра» или «центры» — они находятся, главным образом, в субкортикальных структурах), значительная часть коры, ее так называемые ассоциативные отделы, демонстрирует более распределенный, сетевой тип организации, когда каждая группа нейронов связана со всеми остальными. Это различение, впрочем, не является очень строгим. Сетевой тип организации демонстрирует и так называемая восходящая ретикулярная активирующая система {Ascending Reticular Activating System, ARAS), локализованная в глубоких субкортикальных структурах ствола, а также среднего и промежуточного мозга. Ее функция состоит в широкой активации коры, без чего состояния бодрствования и внимания оказываются невозможны (см. 4.4.1)22. Модулярные и коннекционистские подходы в когнитивных исследованиях опираются на данные о подобной горизонтальной организации самого мозгового субстрата.
Помимо и сверх того, мозговые структуры сохраняют следы их эволюционного происхождения, которые обуславливают моменты иерархических отношений между ними. Разумеется, эти иерархические, межуровневые отношения выражены не так явно, как в случае структуры бюрократических учреждений (типа какого-нибудь министерства или крупного университета). Часто они оставляют впечатление скорее градуальных переходов (Гольдберг, 2004). Существуют три основных градиента эволюционного развития мозга. Первый и наиболее ранний из них связан с переходом от древнейших, субкортикальных к кортикальным структурам (Grillner et al., 2005). Внутри коры больших полушарий, которая, кстати говоря, имеется в развитом виде только у млекопитающих (Aboitiz, Morales & Montiel, 2003), возник второй выраженный градиент развития — от задних к более новым, передним (лобным, или фронтальным) структурам. Данные эволюционные изменения продолжались вплоть до начала антропогенеза (6—7 млн. лет назад), который иногда даже называют в литературе «эпохой лобных долей» (рис. 2.16). На этой последней фазе эволюции внутри самих передних отделов коры наметился еще один градиент роста, сопровождающийся ускоренным разви-
2
 2 В последние десятилетия было, впрочем, обнаружено, что в действительности эта система состоит из нескольких параллельных подсистем со своими начальными структурами в субкортикальных областях и конечными в коре Более того, отдельные подсисте-156 мы имеют различную биохимическую основу (см в особенности 9 4 3)
2 В последние десятилетия было, впрочем, обнаружено, что в действительности эта система состоит из нескольких параллельных подсистем со своими начальными структурами в субкортикальных областях и конечными в коре Более того, отдельные подсисте-156 мы имеют различную биохимическую основу (см в особенности 9 4 3)