
Экология и биоэнергетика некоторых зимующих видов птиц Якутии 03. 00. 16 Экология
| Вид материала | Автореферат диссертации |
- Программа курса Методика организации курса \"Экология Тамбовской области\", 78.43kb.
- Экология Вопросы к зачёту, 25.45kb.
- Лекция 1 Что такое экология. Разделы экологии Термин «экология», 148.25kb.
- Динамика видов зимующих птиц парка. Мониторинг, 539.69kb.
- Тематика лекций (Наименование тем, содержание, объём в часах) № п/п, 74.97kb.
- Экология как наука Экология, 193.74kb.
- e-sila ru/projects/intro html Виктор Дольник, 2175.5kb.
- Задачи: в различных источниках информации определить птиц, зимующих в нашем городе., 395.76kb.
- Промышленная экология) учебно-методический комплекс по дисциплине Прикладная экология, 384.12kb.
- Пространственно-биотопическое размещение и особенности экологии петрофильных видов, 564.31kb.
На правах рукописи
МОРДОСОВА НАДЕЖДА ИННОКЕНТЬЕВНА
Экология и биоэнергетика некоторых
зимующих видов птиц Якутии
03.00.16 – Экология
Автореферат
диссертации на соискание ученой степени
кандидата биологических наук
Якутск - 2009
Работа выполнена на кафедре зоологии биолого-географического факультета Якутского государственного университета им. М.К. Аммосова и в Институте биологических проблем криолитозоны СО РАН
| Научный руководитель: | доктор биологических наук Ануфриев Андрей Иванович |
| Официальные оппоненты: | доктор биологических наук Соловьев Филипп Петрович, Якутский государственный инженерно-технический институт |
| | кандидат биологических наук, доцент Ларионов Анатолий Геннадьевич, Якутская государственная сельскохозяйственная академия |
| Ведущая организация: | Институт систематики и экологии животных СО РАН г. Новосибирск |
Защита диссертации состоится 2 октября 2009г. в ___ час. на заседании диссертационного совета Д 212.306.03 при ГОУ ВПО «Якутский государственный университет им. М.К. Аммосова» по адресу:
677000, г. Якутск, ул. Белинского, 58. Факс: 8(4112)33-68-76.
E-mail: sparrow_passer@rambler.ru
С диссертацией можно ознакомиться в научной библиотеке ГОУ ВПО «Якутский государственный университет им. М.К. Аммосова» по адресу:
г. Якутск, ул. Белинского, 58.
Автореферат разослан: сентября 2009г.
Ученый секретарь
диссертационного совета, д.б.н. Н.С. Данилова
ОБЩАЯ ХАРАКТЕРИСТИКА РАБОТЫ
Актуальность темы. Энергетический обмен живого организма – одна из наиболее важных проблем современной биологии, являющихся предметом всестороннего изучения. Существенное место в этой проблеме занимает изучение закономерностей теплового обмена животных в естественных условиях, который находится под постоянным воздействием сложного комплекса внешних факторов (Шилов, 1968). Основными целями 100-летнего изучения энергетических процессов класса птиц было понять «устойчивость» птицы как энергетической машины и выяснить количество энергии затрачиваемой птицей при жизни в природе (Дольник, 1995). Исследования биоэнергетики птиц в их естественном окружении активизировались после предложенного Кенди (Kendeigh, 1944) метода прямой калориметрии, введения показателя «энергия существования», и установления возможности расчета аллометрических уравнений (Kendeigh et all., 1977). В настоящее время исследованиями ряда отечественных и зарубежных специалистов накоплен значительный основополагающий материал, характеризующий биоэнергетику птиц в условиях их естественного окружения (Шилов, 1968; Дольник, 1975; 1978; 1982; 1995; Дольник, Гаврилов, 1982; Дольник, Гаврилов, Постников, 1982; Гаврилов, 1972; 1974; 1995а, б; 2000а; Гаврилов, Дольник, 1976; Андреев, 1977; 1978; 1980; Kendeigh, 1944; Kendeigh et all., 1977; Pearson, 1954; Orians, 1961; Shartz, Zimmerman, 1971; Utter, 1971; Wolf, Heinsworth, 1972). Подобные исследования продолжаются и в настоящее время (Reich et all., 2006; Chown, 2007; Seibel, 2007; Makarieva et all., 2004; 2006; 2008). Изучение особенностей поддержания энергетического баланса некоторых видов оседлых воробьиных птиц Якутии проводилось Н.А. Находкиным (1988а). Его исследования проводились при различной температуре среды в течение годового цикла жизнедеятельности птиц. В основном он изучал пищевую активность и баланс энергии некоторых видов воробьиных птиц. Однако многие вопросы экологии, терморегуляционные возможности, уровень метаболизма, температура тела и их изменение в годовом цикле жизнедеятельности, активность и другие важные вопросы адаптации птиц к суровым условиям Центральной Якутии оставались неизученными. Совершенно не были изучены биоэнергетика крупных тетеревиных и врановых птиц в этом регионе. Решение этих вопросов весьма актуально в изучении энергетического обмена животных высоких широт.
Цель и задачи исследования. Основная цель работы - выявить эколого-физиологические механизмы адаптации у ряда видов зимующих в Центральной Якутии воробьинообразных и курообразных птиц, обеспечивающие успешное их выживание в условиях резкоконтинентального климата. В процессе работы выполнялись следующие задачи:
- Изучить особенности терморегуляции в условиях естественного зимнего диапазона температур.
- Определить оптимальный температурный режим существования в зимний период.
- Изучить динамику температуры тела в годовом цикле жизнедеятельности и определить его зависимость от температуры среды.
- Изучить динамику двигательной активности мелких видов воробьинообразных в годовом жизненном цикле и определить его зависимость от температуры среды.
Научная новизна и теоретическая значимость. В результате выполненных исследований впервые установлено, что мелкие и крупные по размерам тела виды птиц, зимующие в Якутии, обладают набором специфических эколого-физиологических адаптаций, позволяющих им успешно существовать в условиях холодного климата. Все исследованные виды обладают развитой химической терморегуляцией. Базальный метаболизм у птиц в зимнее время находится на относительно невысоком уровне, и даже ниже, чем в летнее время. У крупных зимующих видов птиц (врановых и тетеревиных) установлена разная стратегия температурных адаптаций. Врановые, питаясь высококалорийными кормами, постоянно поддерживают высокий уровень метаболизма и температуру тела. У тетеревиных, питающихся низкокалорийной пищей и использующих убежища в снегу в холодное время года отмечена сезонная и суточная гипотермия. У мелких видов зимующих воробьиных и у тетеревиных наблюдается выраженная суточная и сезонная гипотермия, связанная с изменением температуры среды обитания. Двигательная активность у изучаемых видов мелких воробьиных в годовом жизненном цикле имеет отчетливую сезонную изменчивость. Результаты выполненных исследований являются основой для дальнейшего изучения функционирования различных физиологических систем у зимующих птиц на клеточном и молекулярном уровнях.
Практическая значимость. Зимующие воробьиные и тетеревиные птицы Центральной Якутии могут быть модельным объектом для изучения физиологических механизмов температурных адаптаций. Полученные материалы могут быть использованы при разработке мер по охране зимующих птиц в критических условиях зимней жизни, а также при чтении лекций по экологии и экологической физиологии студентам биологических и сельскохозяйственных специальностей. Исследования проводились при финансовой поддержке РФФИ, проект 08-04-00047-а (2008-2010), полученные результаты используются при выполнении Интеграционного проекта СО РАН № 137 «Адаптации популяций и формирование сообществ птиц в условиях криолитозоны» (2009 - 2011).
Основные положения, выносимые на защиту:
- У зимующих видов птиц при переживании зимних холодов используются общебиологические и специфические адаптационные механизмы, приобретенные в суровых условиях Центральной Якутии.
- Врановые и тетеревиные птицы используют различные способы выживания в условиях холодного климата севера.
Апробация работы: По теме диссертационной работы опубликованы 3 статьи, одна из которых опубликована в журнале из списка ВАК, а также 5 кратких сообщений, опубликованных в материалах конференций и съездов, в т.ч. в Международном журнале Integrative Zoology.
Структура и объем работы: Диссертация состоит из введения, пяти глав, выводов и списка литературы, включающего 129 отечественных и 34 зарубежных источника. Изложена на 153 стр. м.п., содержит 16 таблиц и 41 рисунков.
Глава 1. Энергетический метаболизм у птиц, зимующих в условиях холодного климата (обзор литературы)
В данной главе обсуждаются имеющиеся в литературе данные по морфологическим, экологическим приспособлениям, особенностям терморегуляции и значение снежного покрова в жизни птиц высоких широт.
В главе рассматриваются: 1. Основные понятия и термины; 2. Основные формы активности; 3. Энергетика птиц различной экологической специализации; 4. Самоподдержание или существование; 5. Характеристика основных эколого-физиологических механизмов адаптаций зимующих птиц к условиям среды; 6. Зимующие птицы Якутии; 7. Терморегуляция в жизни северных птиц; 8. Морфологические приспособления птиц к холоду; 9. Особенности питания в зимнее время; 10. Роль снежного покрова в жизни птиц.
Базальный метаболизм или основной обмен принято понимать как минимальное количество энергии, необходимое для нормального существования организма и поддержания основных жизненных потребностей в покое (Дольник, 1995). Расход энергии при выбранной стандартной температуре среды в 00С, при которой птицы заведомо находятся в состоянии холодовой нагрузки – стандартный метаболизм. Диапазон температур среды, в котором расход энергии остается на неизменном уровне – термонейтральная зона. Расход энергии в этой зоне – базальный метаболизм (Гаврилов, 1979). Проникновение вида в новую для него среду перестраивает его морфологию и поведение таким образом, чтобы снять избыточную нагрузку на бюджет энергии. Затраты видов, обитающих в суровых условиях сопоставимы с затратами видов, живущих в мягких условиях.
У птиц, обитающих в резко-континентальном климате сезонные вариации базального метаболизма более выражены, чем у птиц, обитающих в субтропиках и тропиках. Сезонная цикличность базального метаболизма чаще встречается и сильнее выражена у мелких видов птиц, чем у крупных: у мелких обычно происходит повышение уровня базального метаболизма в зимний период, а у крупных (массой тела более 200 г) – понижение. Воробьиные птицы, обитающие в умеренном и холодном климате, каким-то образом осуществляют повышение базального метаболизма на зиму, адаптивно по отношению к холодным зимним условиям (Дольник, 1995).
Врановые птицы в значительно большем диапазоне температур среды способны сохранять неизменную теплопродукцию, изменяя теплоотдачу. Изменение теплоотдачи происходит за счет хорошо развитой циркуляторной системы, изменения положения перьевого покрова, позы птицы. Крупные виды врановых хорошо сохраняют тепловой баланс при низких температурах среды (Гаврилов, 1979).
Воробьиные птицы имеют иные характеристики метаболизма, чем неворобьиные (Lasiewsky, Dawson, 1967; Kendeigh, 1969, 1970, 1972; Дольник, 1969). Различия касаются химической терморегуляции – скорости теплопродукции, которая у воробьиных птиц выше. Особенности метаболизма адаптивны для мелких размеров, но не адаптивны для крупных. Врановые сумели преодолеть отрицательные последствия крупных размеров тела путем физической терморегуляции (Дольник, 1974).Впервые в отечественной литературе проблему сохранения энергетического баланса организма как основу адаптации к существованию в определенной среде обитания сформулировал Н.И. Калабухов (1946). Он первым провел оценку влияния условий существования на организм в целом и сформулировал заключение, что интеграция приспособлений определяет общую характеристику организма – его энергетический баланс. Влияние внешней среды вызывает реакции организма, направленные на поддержание и сохранение этого баланса.
Глава 2. Материал и методики исследований
Исследования проводились в 2005 - 2009гг. на следующих видах: обыкновенная чечетка (Acanthis flammea Linnaeus, 1758), большая синица (Parus major Linnaeus, 1758), полевой воробей (Passer montanus Linnaeus, 1758), обыкновенный снегирь (Pyrrhula pyrrhula Linnaeus, 1758), свиристель обыкновенный (Bombycilla garrulus Linnaeus, 1758), дикуша азиатская (Falcipennis falcipennis Hartlaub, 1855), обыкновенный тетерев (Lyrurus tetrix Linnaeus, 1758), каменный глухарь (Tetrao parvirostris Bonaparte, 1856), черная ворона (Corvus corone Linnaeus, 1758) и ворон (Corvus corax Linnaeus, 1758). Птиц отлавливали осенью во время массового лета, кочевок и миграций с помощью орнитологических сетей и манных птиц. Снегири были отловлены специальной ловушкой сконструированной по типу «хлопушки». В экспериментальной работе было отловлено и использовано 115 особей. Каменный глухарь и тетерев были отловлены птенцами в июле 2006г. в Олекминском и Хангаласском улусах республики. Дикуша в возрасте 3-х лет получена из Карасукского питомника Института систематики и экологии животных СО РАН, (Новосибирская обл.) в 2007г. Ворона и ворону отловили в окрестностях г. Якутска. Крупные зимующие птицы содержались в республиканском зоопарке «Орто-Дойду» в условиях естественного температурного фона - в открытых вольерах. В ночное время тетеревиные укрывались в вольерах в утепленных убежищах. На время экспериментальных работ по газообмену птиц привозили в виварий Ботанического сада ИБПК СО РАН. Объем исследованного материала приведен в табл. 1.
Таблица 1
Объем исследованного материала
| Вид работы | Вид птицы | Количество опытов или анализов | Итого | |
| Активность в течение суток (число срабатываний контактной площадки) | Чечетка* | 26 868 | 353 459 | |
| Полевой воробей* | 134 440 | |||
| Снегирь * | 163 094 | |||
| Свиристель* | 29 057 | |||
| Потребление кислорода (опыт) | 2778 определений | 2778 | ||
| Температура тела (количество измерений) | Большая синица | 34 | 49 565 | |
| Полевой воробей | 174 | |||
| Снегирь обыкновенный | 129 | |||
| Свиристель обыкновенный | 116 | |||
| Дикуша азиатская | 8192 | |||
| Обыкновенный тетерев (2 экз.) | 16 364 | |||
| Каменный глухарь (2 экз.) | 16 364 | |||
| Ворон | 8129 | |||
* - прыжков птицы
Измерение температуры тела у крупных птиц. Крупным зимующим птицам (обыкновенный тетерев, каменный глухарь, ворон) в межлопаточной области под кожу были имплантированы термографы DS 1922 L-F5 («Dallas semiconductor Corp.»). DS 1922 L/T (temperature logger iButtons) – является самодостаточной системой, представляет собой плоский цилиндр диаметром 17,4 мм, толщиной 6 мм, массой около 3 г. Перед вживлением приборы программировались таким образом, чтобы измерения температуры тела проводились с частотой 1 раз в 60 или в 120 мин. Данные снимали с извлеченных датчиков при помощи однопроводного адаптера компьютера.
Измерение температуры тела у мелких воробьиных птиц. Измерение температуры тела проводили при помощи имплантируемого под кожу птицы в подмышечной области температурного датчика. Этот датчик длиной в 1см, округлый, в сечении 1мм диаметром, разработан по технологии Био-Термо зарегистрированной торговой марки Digital Angel Corporation (производство Япония). Измерения выполнялись ручным (карманным) сканирующим устройством, считывающим измеряемую величину при приближении сканера на расстоянии 2 - 4 см от датчика. Данные снимали в течение светлого времени суток на улице в естественных температурных условиях и для сравнения в теплом помещении. При проведении измерения температуры в течение суток, исследуемые птицы находились в клетках на улице. Операции по вживлению датчиков проводились к.с-х.н. В.Ф. Ядрихинским в клинике ветеринарного факультета ЯГСХА (г. Якутск).
Суточная активность. Регистрировали путем подсчета прыжков испытуемой птицы на жердочке. Устройство работает при любой температуре и может устанавливаться вблизи клетки. В качестве регистратора использовались персональные компьютеры с программой, написанной инженером ИБПК СО РАН Г.Г. Архиповым.
Уровень метаболизма и терморегуляция. Потребление кислорода измеряли у птиц по описанной ранее методике (Ахременко, Ануфриев, 1984), с использованием переносного автоматического газоанализатора ПГА-12. Для стабилизации температуры внешней среды использовали термостатируемую камеру, в которой поддерживалась заданная температура в интервале от -400С до +350 – +400С. Респираторные камеры изготавливали из фанеры и обклеивали клейкой пленкой. Для изучения газообмена чечетки, большой синицы и полевого воробья использовались камеры меньшего размера, чем для снегиря и свиристеля. Для крупных птиц использовали респираторные камеры объемом от 10 до 40 литров. Обработку полученных данных производили с использованием компьютерной программы, разработанной инженером ИБПК СО РАН Г.Г. Архиповым. Статистическую обработку проводили общепринятыми методами.
Глава 3. Терморегуляция и уровень метаболизма у зимующих птиц
Терморегуляция в полной мере наблюдается только у полностью гомойотермных организмов. Реакции организма на охлаждение в отдельных физиологических состояниях различны и не всегда могут быть уложены в рамки представлений о постоянстве внутренней среды (Слоним, 1986). Мелкие воробьиные имеют самую высокую температуру тела среди всех воробьинообразных и более того, среди всех гомойотермных животных (Гаврилов, 1995; Дольник, 1995).
Обыкновенная чечетка (Acanthis flammea Linnaeus, 1758). Исследование терморегуляторных реакций у чечетки в зимний период в диапазоне от -400С до 300 - 350С показало наличие выраженных реакций на изменение температуры среды. У чечеток в связи с более мелкими размерами и массой тела, уровень метаболизма более высокий, чем у других мелких воробьиных. При снижении температуры среды от 200С до минус 40 - 50С метаболизм увеличился на 15% со скоростью 0,03 мл О2/г·ч. При снижении температуры от 50 - 80С до минус 400 - 450С, метаболизм возрос на 35 - 40% со скоростью 0,375 мл О2/г·ч. Динамика суточных изменений метаболизма в зимнее время совпадает с изменениями температуры тела у мелких воробьиных - повышенный уровень метаболизма в утренние и дневные часы с понижением в ночное время. Измерения проводились в декабре, когда температура среды в ночные часы опускалась до минус 350 - 400С. В летнее время, при изменении температуры среды от +150С до +300С потребление кислорода у чечеток существенно не менялось.
Большая синица (Parus major Linnaeus, 1758). В зимнее время, как и у остальных видов мелких воробьиных, отмечена активная реакция на изменение температуры окружающей среды. Так при снижении температуры с +280С до -220С потребление кислорода увеличивается в два раза. В диапазоне температур от минус 300С до минус 400С потребление кислорода возрастает в 2,5 раза со скоростью 0,047 мл О2/г·ч. Скорость роста значительно выше чем у чечетки (0,03 мл О2/г·ч), но ниже чем у полевого воробья (0,06 мл О2/г·ч).
Полевой воробей (Passer montanus Linnaeus, 1758). У полевого воробья в зимнее время при температурах выше 250 установлен наиболее низкий уровень метаболизма. При снижении температуры среды до минус 200 - 250С потребление кислорода увеличивается в три раза. Рост метаболизма начинался при достижении температуры 100, и продолжался до температуры минус 250 - 300С. Максимальный уровень метаболизма у неподвижно сидящей птицы достигал 5 - 6 мл О2/г·ч. При снижении температуры среды от 200С до минус 300С интенсивность роста уровня метаболизма составляла 0,06 мл О2/г·ч. Это на 30% ниже данного показателя у снегиря. У этих двух видов максимальные уровни метаболизма при низких температурах среды близки по абсолютному значению, но у снегиря уровень минимального обмена значительно выше, чем у полевого воробья. В ночное время (декабрь), у спящих воробьев, при температуре среды минус 200С - 300С уровень метаболизма был относительно стабилен. Увеличение уровня метаболизма зарегистрировано в ранние утренние часы, за 1 - 2 часа до рассвета. В летнее время у полевого воробья в пределах термонейтральной зоны до 260С уровень метаболизма изменяется незначительно, далее происходит резкое увеличение потребления кислорода. Потребление кислорода в пределах температурного оптимума составляет 2,5 - 3,5 мл О2/г·ч.
Снегирь (Pyrrhula pyrrhula Linnaeus, 1758). Во время трехчасовой экспозиции (дневное время) в респираторной камере, при снижении температуры среды от плюс 280С до минус 280C, уровень метаболизма возрос на 28% с интенсивностью 0,04 мл О2/г·ч на 10С. Реакция на изменение температуры происходит моментально. Изучение температурной зависимости метаболизма показало, что в зимнее время нижняя граница термонейтральной зоны находится в диапазоне 50 - 60С, при снижении температуры среды до минус 350С потребление кислорода возрастало на 20% с интенсивностью 0,02 - 0,03 мл О2/г·ч на 10С.
Рост метаболизма у неподвижной птицы происходил до температур минус 300 - 350С, при более низких температурах происходила либо стабилизация роста, либо снижение потребления кислорода. В ряде случаев отмечено: при низких температурах, когда птицы начинали двигаться происходил значительный рост метаболизма. При температурах среды выше 250С также отмечено увеличение потребления кислорода. Летом уровень метаболизма в зоне температурного оптимума немного ниже, чем в декабре - январе. При температуре среды 200 - 250С (термонейтральная зона) потребление кислорода у птиц в неподвижном состоянии изменяется незначительно. При температуре выше 250С, происходит рост уровня метаболизма.
Свиристель (Bombycilla garrulus Linnaeus, 1758). У данного вида установлены выраженные метаболические реакции на изменение температуры среды. В экспериментах при снижении температуры среды от +250 до -300С, метаболизм возрос более чем на 70%. В этом диапазоне скорость роста метаболизма составила 0,013 мл О2/г·ч, что было значительно выше, чем у полевого воробья и снегиря. Изучение терморегуляторных реакций в зимнее время показало, что у этого вида нижняя граница термонейтральной зоны находится в диапазоне температур 80 - 100С, верхняя граница около 300 - 350С. В летний период терморегуляция свиристеля претерпевает закономерные изменения. В июне, в гнездовой период, птицы становятся более чувствительными к изменению окружающей температуры. В диапазоне температур 50 - 100С уровень метаболизма относительно постоянен. В целом, у свиристеля в летний период в диапазоне температур от 60 до 300С изменение уровня метаболизма составляет 40 - 50%.
Дикуша (Falcipennis falcipennis Hartlaub, 1855). Зона оптимальных для дикуши температур расположена в интервале от 50 - 70С до 170 - 200С. В границах приведенной температурной зоны потребление кислорода у взрослого самца массой около 700 г составила 1,1 - 1,2 мл О2/г·ч. При снижении температуры среды до -200С потребление кислорода возрастает почти в два раза. При температурах среды выше 200С также отмечен рост уровня метаболизма. В температурном интервале от плюс 100 до минус 200С рост уровня метаболизма на 10С составил 0,043 мл О2/г·ч (табл. 2).
Таблица 2
Показатели интенсивности уровня метаболизма у тетеревиных и врановых
| Вид | Минимальный уровень потребления кислорода (мл О2/г·ч) | Максимальный уровень потребления кислорода (мл О2/г·ч) | Величина роста метаболизма (мл О2/г·ч)/% | Интенсивность роста метаболизма (мл О2/г·ч на 10 С) |
| Глухарь | 0,6 | 1,05 | 0,45/42,8 | 0,013 |
| Тетерев | 0,8 | 2,0 | 1,2/60 | 0,043 |
| Дикуша | 1,0 | 2,2 | 1,2/54,5 | 0,043 |
| Ворон | 1,8 | 3,6 | 1,8/50,0 | 0,06 |
| Ворона | 2,6 | 4,7 | 2,1/44,7 | 0,07 |
Обыкновенный тетерев (Lyrurus tetrix Linnaeus, 1758). У взрослого самца, массой 1210 г минимальный уровень метаболизма отмечен в интервале температур от 50 до 200С и составил 0,9 - 1,1 мл О2/г·ч. При температуре минус 180 - 200С возрастал до 2,0 - 2,1 мл О2/г·ч. При повышении температуры среды до плюс 200 - 250С, также установлен рост потребления кислорода. Несмотря на то, что размеры и масса тела тетерева превышают таковые у дикуши, ряд показателей метаболизма у этих видов сходны. Так, величина и интенсивность роста метаболизма на 10С у них близки по абсолютной величине (табл. 2).
Каменный глухарь (Tetrao parvirostris Bonaparte, 1856). У взрослого самца, массой 3480 г минимальный уровень метаболизма определен в интервале температур от 50 до 150С, и составил 0,6 - 0,7 мл О2/г·ч. При снижении температуры воздуха до минус 280 - 300С потребление кислорода возрастает на 40 - 50%, а рост уровня метаболизма был значительно ниже, чем у дикуши и тетерева, как по абсолютной величине, так и в процентном выражении (табл. 2). Таким же низким был и рост метаболизма при снижении температуры воздуха на каждый градус. У этого вида, имеющего крупные размеры и массу тела по сравнению с другими исследованными нами тетеревиными птицами, уровень метаболизма значительно ниже.
Черная ворона (Corvus corone Linnaeus, 1758). Зона оптимальной температуры находится в интервале от 50 - 70С до 150 - 170С. При снижении температуры среды до -250С отмечен рост потребления кислорода почти на 45% (табл. 2). У вороны, по сравнению с другими рассматриваемыми видами, отмечен наибольший рост интенсивности метаболизма при снижении температуры среды на 10С - 0,07 мл О2/г·ч.
Ворон (Corvus corax Linnaeus, 1758). У ворона (самец - 1290 г) термонейтральная зона в зимнее время смещена в область более низких температур по сравнению с представителями тетеревиных птиц. Химическая терморегуляция включается при температурах ниже 50С. Рост метаболизма отмечен также при температурах выше 170 - 200С. При снижении температуры среды до минус 250С потребление кислорода возрастало на 50%, а интенсивность химической терморегуляции составляет 0,06 мл О2/г·ч на каждый 10С.
Глава 4. Температура тела зимующих птиц
У полевого воробья температура тела в зимнее время (декабрь - январь) при температуре внешней среды минус 400 - 450С изменялась в пределах 38,90 - 40,70С (табл. 3).
Таблица 3
Дневная температура тела у полевого воробья (декабрь - январь)
в первой половине дня (10 - 15 ч)
| Показатель/среда | Естественная температура | Комнатная температура | |
| Температура среды, 0С | n(M±m) | 9 (-42,58±2,35) | 9 (20,80±0,49) |
| Min-Max | -26 – -48 | 19,5 – 22,5 | |
| Температура тела, 0С | n(M±m) | 9 (39,88±0,18) | 9 (41,36±0,32) |
| Min-Max | 38,9 – 40,7 | 38,4 – 41,7 | |
n - количество измерений
Как видно из материалов табл. 3. температура тела у воробья в условиях оптимальных температур (200C) среды колебалась в пределах 38,40 - 41,70C. В естественной среде усредненные значения температуры их достоверно ниже, чем у птиц находящихся при оптимальных температурах. У большой синицы в этот же временной период при естественных температурах среды температура тела изменялась в пределах (39,70 – 42,50С), а в условиях температурного оптимума (41,40 – 42,60С) (табл. 4). Разница усредненных значений достигала 10С. У снегиря в январе в дневное время при температуре окружающей среды минус 400 – 420С температура тела изменялась в диапазоне (39,4 – 41,30С), в оптимальной температурной зоне (200С) температура тела изменялась в пределах (41,80 – 43,90С), в среднем была на 1,80C выше. В июне, при внешних температурах около 200C температура тела снегиря изменялась в дневное время в пределах (41,8 – 44,9)0С, а усредненные значения практически не отличались от температуры его тела зимой в оптимальных условиях (табл. 5). Наблюдение суточной динамики температуры тела полевого воробья и обыкновенного снегиря в зимние месяцы показали, что в период с 21 ч до 4 – 5ч у полевого воробья наблюдается снижение его до -400C. Период с повышенной температурой тела продолжался с 9 до 17 ч, и по времени полностью совпадал с периодом двигательной активности птиц. При этом установлена положительная корреляция между температурой среды и тела, коэффициент корреляции 0,7. У снегиря температура тела ниже 400C была установлена в вечерне-ночные часы. После 18 ч температура тела снегиря не поднималась выше 39,50C до утра. Наиболее высокая температура тела отмечена в утренние часы, затем она постепенно снижается. Как и у полевого воробья, у снегиря присутствует положительная корреляция между температурой тела и среды. Коэффициент корреляции также был равен 0,7.
Таблица 4
Дневная температура тела большой синицы (декабрь - январь)
в первой половине дня (10 - 15 ч)
| Показатель/среда | Естественная температура | Комнатная температура | |
| Температура среды, 0С | n(M±m) | 9 (-42,58±2,35) | 9 (20,80±0,49) |
| Min-Max | -26 – -48 | 19,5 – 22,5 | |
| Температура тела, 0С | n(M±m) | 7 (41,13±0,35) | 9 (42,08±0,28) |
| Min-Max | 39,7 – 42,5 | 41,4 – 42,6 | |
n - количество измерений
Таблица 5
Дневная температура тела снегиря (январь, июнь)
в первой половине дня (10 - 12 ч)
| Показатель/среда | Январь (в естественной среде) | Январь (в помещении) | Июнь (в естественной среде) | |
| Температура среды, 0С | n(M±m) | 9 (-41,32±0,10) | 9 (20,80±0,49) | 24 (19,54±0,78) |
| Min-Max | -40,9 – -41,9 | 19,5 – 22,5 | 15,0 – 25,0 | |
| Температура тела, 0С | n(M±m) | 9 (40,66±0,22) | 7 (42,44±0,28) | 23 (42,57±0,13) |
| Min-Max | 39,4 – 41,3 | 41,4 – 42,6 | 41,8 – 44,9 | |
n - количество измерений
Ворон (Corvus corax Linnaeus, 1758). Температура тела ворона на протяжении годового цикла относительно стабильна и поддерживается на достаточно высоком уровне (рис. 1.).
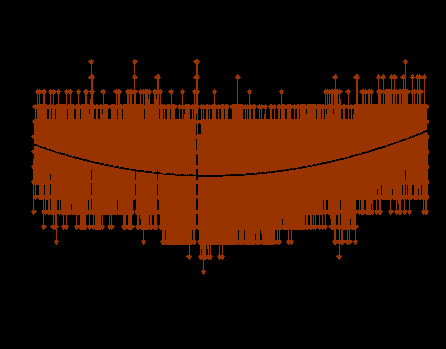
Рис. 1. Динамика температуры тела у ворона в годовом цикле
Наблюдаются колебания среднесуточной температуры тела, наиболее высокая температура тела зарегистрирована в летние месяцы: май (39,6±0,04)0C, июнь (39,9±0,04)0C, июль (39,6±0,09)0C, далее она начинает снижаться, достигая минимальных значений в январе (38,5±0,06)0С. Абсолютно низкие температуры его тела +35,50 – +360C отмечены в ночное время с декабря по февраль, и наиболее высокие в дневное время суток – до +42,00 – +42,50C встречались в течение всего года. Амплитуда изменений температуры тела по абсолютным значениям в течение года составила 70С (Ануфриев и др., 2008а, 2008б). Среднесуточная температура тела в течение года была однонаправлена с температурой окружающей среды, коэффициент корреляции +0,95.Диапазон изменений среднемесячной температуры тела относительно невелик, по сравнению с таковыми у других видов животных, подвергнутых подобного рода исследованиям (бурундук, суслик, заяц-беляк, псовые: Ануфриев и др., 2007б; Соломонов и др., 2008; Ядрихинский, Ануфриев, 2008). Суточная динамика температуры тела на протяжении всего года сходная: повышение в дневное время суток и понижение в ночное время. Коэффициент корреляции температуры тела и температуры среды 17 июля был +0,96, 27 декабря +0,65. На протяжении первой половины года у ворона среднесуточная температура тела возрастает, на протяжении второй снижается. Годовой диапазон изменений среднесуточной температуры составляет 1,40С, а по абсолютной величине 70С. Таким образом, относительно высокий уровень метаболизма позволяет ворону даже в периоды низких годовых температур стабильно поддерживать высокую температуру тела. В зимнее время отсутствовали даже признаки гипотермии.
Обыкновенный тетерев (Lyrurus tetrix Linnaeus, 1758). У тетеревов колебания среднесуточной температуры тела в течение года более выражены, чем у ворона. Наиболее высокие среднегодовые температуры тела у тетеревов установлены в летние месяцы 390 – 400С, в последующие месяцы она начинает постепенно снижаться, и достигает минимальных годовых значений в декабре – январе (рис. 2.). В эти месяцы у него зарегистрированы абсолютно низкие температуры тела. У одной из птиц понижение температуры тела доходили до 280С, у другой - 330С. В целом, минимальная среднемесячная температура отличалась от максимальной у одной птицы на 3,340С, у второй на 3,00С, у ворона это значение составляет 1,380С. Среднесуточная температура тела у тетерева в годовом жизненном цикле имеет отчетливую зависимость от температуры среды. Она более выражена, чем у ворона. В течение суток температура тела тетеревов изменялась следующим образом, в летний период пониженные температуры тела (390 - 39,50С) наблюдались в ночные часы. С восходом солнца и началом активности птиц температура тела нарастала, достигая значений 400С. Коэффициент корреляции температуры тела и температуры среды 1 августа был равен 0,64. В зимний период в темное время суток температура тела поддерживается на уровне 350 - 370С, в светлое время суток она повышалась до 380 - 390С. В период зимних холодов зависимость температуры тела от температуры окружающей среды, несмотря на использование естественных убежищ (снега) для ночевки, возрастает. Так, 18 января коэффициент корреляции температуры тела и температуры среды составлял 0,8.
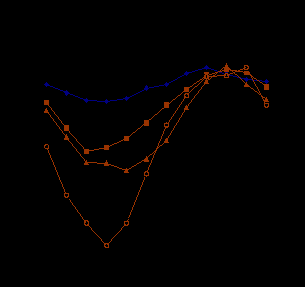
Рис. 2. Динамика среднесуточной температуры тела ворона (1), тетерева(2), глухаря (3) и среднесуточной температуры среды (4)
Каменный глухарь (Tetrao parvirostris Bonaparte, 1856). Динамика температуры тела глухаря была сходной с таковой у тетерева, однако среднесуточная температура его тела во все сезоны года была ниже, чем у тетерева. Относительно высокая масса (свыше 3 кг) и размеры тела глухаря позволяют ему быть энергетически более экономичным. В зимний и в летний периоды зависимость температуры тела от изменений температуры среды одинаковы. Коэффициент корреляции 27 июля и 2 февраля составил 0,71.
Глава 5. Суточная активность у мелких воробьиных птиц
в годовом жизненном цикле
Полевой воробей (Passer montanus Linnaeus, 1758). В начале весны двигательная активность полевого воробья равна световому дню, летом при круглосуточном освещении она несколько меньше (табл. 6).
Таблица 6
Продолжительность активности полевого воробья и длина светового дня
| Дата | Длина периода активности | Длина светового дня | Разность |
| 07.12 | 07 ч 45мин | 7 ч 15 мин | 30 мин |
| 28.12 | 07 ч.30 мин | 5ч 25 мин | 2ч 05 мин |
| 07.01 | 07 ч30 мин | 6 ч 45мин | 45 мин |
| 12 .01 | 09 ч 00 мин | 6 ч 55 мин | 2 ч 05 мин |
| 13.07 | 19 ч 53 мин | 24 ч | -4 ч 07 мин |
| 15.07 | 18 ч 19мин | 24 ч | -5 ч 41 мин |
Минимальное время активности (полетов и перемещений) у полевого воробья в зимний период составляет не менее 7 ч 30 мин., вероятно, данное время является минимально необходимым для запасания достаточного количества энергии для успешной ночевки. Зимой полевые воробьи проявляют только те формы активности и в том минимально возможном количестве, какое необходимо для их существования. Такие формы активности, как драки, «активное безделье» и другие, требующие дополнительных затрат энергии, практически полностью отсутствуют, а имеющиеся формы активности сведены до минимума. Например, полет составляет всего около 12 минут в сутки. Вся их активность в это время года связана в основном с добыванием пищи. В течение года основные характеристики двигательной активности полевого воробья значительно колеблются. Низкий уровень активности характерен на протяжении всей зимы. Двигательная активность полевого воробья значительно изменяется в различные сезоны года. Нами установлено, что время его активности в зимнее время длиннее светового дня и составляет 7,30 – 9,00 ч (табл. 6.). По-видимому, этот промежуток времени является минимально необходимым для запасания достаточного количества энергии для успешного существования в течение длительного ночного времени. В это время года воробей проявляет активность, связанную с поиском пищи, а другие формы активности сведены до минимума и проявляются в том количестве, какое необходимо для существования. Такая низкая активность характерна не только в зимние месяцы, но и в начале весны – в марте-апреле. Затем она начинает увеличиваться и достигает максимума в мае. Это совпадает с временем начала размножения и, по-видимому, не позволяет считать ее за миграционную активность. В этот период времени активность его равна длине светового дня. При наступлении круглосуточного освещения в июне-июле активность птиц составляет 18 ч 19 мин – 19 ч 53 мин. В течение летне-осеннего периода происходит сокращение двигательной активности, что связано с сокращением длины светового дня. В сентябре она составляет 10 ч 40 мин. В это время количество прыжков составило 651, или 54 прыжка за 1 ч. В последующие месяцы происходит дальнейшее снижение активности и в декабре-январе составляет 400 - 600 прыжков, что на порядок ниже, чем летом – 6000 - 8000 прыжков.
Cнегирь (Pyrrhula pyrrhula Linnaeus, 1758). Двигательная активность снегиря изменялась в течение года и зависела от продолжительности светлой части суток и температуры окружающей среды. Во все сезоны года графическая кривая активности снегирей двухвершинная. В период круглосуточной освещенности первые срабатывания контактной жердочки зарегистрированы между 3 и 4 часами. С 4 до 10 ч активность птиц максимальная. В это время птицы интенсивно кормились, ухаживали за перьями, совершали перелеты. Затем в период наивысшего солнцестояния активность резко снижалась и находилась на относительно низком уровне в период высоких суточных температур воздуха. Второй, менее значительный, пик активности отмечается в вечерне-ночное время, после 20 ч. В это время птицы выбирали затемненные части вольера, спали или сидели с разведенными в стороны крыльями. Зимой в период наиболее низких годовых температур сохраняется двухвершинный тип суточной активности на фоне общего снижения активности (рис. 3.).
Обыкновенная чечетка (Acanthis flammea Linnaeus, 1758). У этого вида в зимний период двигательная и кормовая активность весьма низкая. Ночной покой у птиц в конце января занимает 15,72 ч в сутки, в дневное время в покое птицы находятся не более 0,14 ч. Поза готовности к действию в покое только 0,14 ч. Основная часть форм активности не требовала больших энергетических затрат: поза готовности к действию 3,17 часа, чистка оперения - 2,25 ч полет - 0,11 часа в сутки.
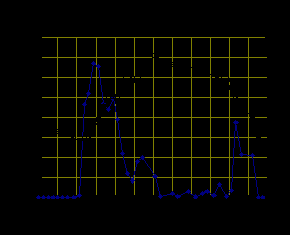
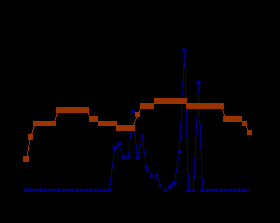
Рис. 3. Суточная активность снегиря в летний и зимний периоды
Время полетов относительно небольшое, но продолжительнее, чем у других исследованных воробьиных птиц. Время приема пищи составляет 2 ч, из них 1,5 ч птицы кормятся семенами подсолнечника и только 0,5 ч - пшеном. Двигательная активность у чечеток, также как и у других видов воробьиных птиц, изменяется в течение года и зависит от продолжительности светлой части суток и температуры окружающей среды. В периоды устойчивых зимних холодов и короткого светового дня, их двигательная активность по сравнению с летними месяцами снижается на порядок. Уменьшается время, на протяжении которого птицы активны, снижаются и количественные показатели (число прыжков). Зимой чечеткам для поддержания энергетического баланса необходимо потреблять достаточное количество пищи за относительно короткий светлый период, и иметь соответствующую наполненность желудочно-кишечного тракта перед долгим, до 19 ч, темным периодом.
Выводы
- Мелкие и крупные по размерам тела виды птиц, зимующие в Якутии, обладают рядом специфических эколого-физиологических адаптаций, позволяющих им успешно существовать в условиях холодного климата.
- Изучение уровня метаболизма и терморегуляторных реакций у мелких зимующих воробьиных птиц Якутии показало, что все исследованные виды обладают достаточно развитой химической терморегуляцией. В зимние месяцы нижняя граница термонейтральной зоны находится в интервале температур 00С - 100С. В периоды зимних холодов в ночное время в состоянии сна уровень метаболизма понижается и возрастает в утренние часы.
- Базальный метаболизм у птиц в зимний период находится на относительно невысоком уровне, и даже ниже, чем у птиц в летнее время. Это может быть связано с зимним увеличением массы тела, улучшением теплоизоляционных свойств перьевого покрова, со снижением общего уровня гормональной активности в зимний период и с понижением общей двигательной активности.
- У крупных зимующих видов птиц Якутии отчетливо просматривается разная стратегия температурных адаптаций. Врановые птицы, питаясь высококалорийными кормами, постоянно поддерживают высокий уровень метаболизма и температуры тела. Тетеревиные птицы, питание которых менее калорийно, имеют отчетливую зависимость от температуры среды. Установлено наличие сезонной и суточной гипотермии.
- Полученные данные свидетельствуют о том, что у мелких видов воробьиных птиц в условиях Якутии наблюдается выраженная суточная и сезонная гипотермия. В период ночного сна, с понижением уровня метаболизма, экономия энергии у птиц достигается путем использования механизмов понижения температуры периферических частей тела.
- Суточная активность у мелких видов зимующих воробьинообразных в годовом жизненном цикле имеет выраженную сезонную изменчивость. Активность их минимальна в декабре-январе, в феврале-марте начинает возрастать, достигая годового максимума в мае - июне. Уже в августе, с уменьшением продолжительности светлого времени суток, величина активности птиц снижается.
Список работ, опубликованных по теме диссертации
- Мордосова Н.И. Изменение температуры тела в годовом цикле и уровень метаболизма у ворона при зимних температурах среды / А.И. Ануфриев, Н.Г. Соломонов, А.П.Исаев, В.Ф. Ядрихинский, Н.И. Мордосова //Доклады Академии Наук. - 2008. - Т. 422. - №4. - С. 568-570.
2. Мордосова Н.И. Исследование изменения активности домового воробья (Passer domesticus L.) по сезонам года / Н.И. Мордосова // Вестник Якутского госуниверситета. – Якутск, 2006. - Т. 3. - №3. –С. 145-148.
3. Мордосова Н.И. Две стратегии адаптации крупных зимующих птиц Якутии к низким температурам / А.И.Ануфриев, Н.Г.Соломонов, А.П. Исаев, Н.И. Мордосова // Вестник Якутского госуниверситета. – Якутск, 2008. - Т. 5 - №4. – С. 10-15.
4. Мордосова Н.И. Некоторые пути адаптации домового (Passer domesticus L.) и полевого (Passer montanus L.) воробьев к условиям существования в Якутии / Н.И. Мордосова //VI Межрегиональная научно-практическая конференция молодых ученых, аспирантов и студентов. – Нерюнгри, 2005. – С. 54-55.
5. Мордосова Н.И. Некоторые эколого-физиологические направления адаптации новых видов птиц в условиях г. Якутска / Н.И. Мордосова // Международные научные чтения «Приморские зори – 2005», посвященные 10-летию со дня основания ТАНЭБ. – Владивосток, 2005. - Вып. 1. – C. 34-36.
6. Мордосова Н.И. Некоторые пути адаптации домового (Passer domesticus L.) и полевого (Passer montanus L.) воробьев к условиям существования в Якутии / Н.И. Мордосова // IX Лаврентьевские чтения. – Якутск, 2005. – С. 67-74.
7. Мордосова Н.И. Изменение температуры тела у CORBUS CORAX в годовом жизненном цикле / А.И.Ануфриев, И.А. Сокольников, Н.И. Мордосова //III Международная конференция по мигрирующим птицам Севера Тихоокеанского региона. Тез. докл. - Якутск, 2007. - Ч. 1. – С.12.
8. Mordosova N.I. Ravens (Corvus corax) body temperature change during annual life cycle / N.I. Mordosova, A.I. Anufriev // Integrative Zoology - 2008. – Paris, 2008. - P.73.