
Физиологическая адаптация американских норок (Mustela vizon schr.) и серебристо-черных лисиц (vulpes vulpes L.) при клеточном содержании в условиях забайкалья 03. 03. 01 физиология
| Вид материала | Автореферат |
- Понятие адаптации, 125.13kb.
- Физиологическая характеристика оздоровительных и экстремальных видов спортивной деятельности, 417.86kb.
- Доклад на тему: Топография мышц плечевого пояса методом обычного тонкого препарирования, 53.76kb.
- Вопросы адаптации детей к школе, 133.74kb.
- Коц Я. М. Спортивная физиология. Учебник для институтов физической культуры, 4839.54kb.
- Н. П. Пахомов 1971г. Предисловие Н. П. Пахомов старейший знаток гончих собак и охоты, 917.03kb.
- Рассказ о северском донце, 201.76kb.
- Физиологическая роль сосудистых рефлексогенных зон в интегративной регуляции функций, 653.7kb.
- Физиология желез внутренней секреции, 15.13kb.
- Курсовая работа Адаптация иммигрантов-мусульман в условиях современной России: проблемы, 166.14kb.
Влияние различных рационов кормления
на биохимические показатели крови
Определение уровня биохимических показателей крови и активности ферментов особенно перспективно при изучении влияния состава кормовых рационов на организм американских норок и серебристо-черных лисиц, разводимых в клеточных условиях. Питание – это наиболее древний фактор, к которому в процессе эволюции произошла адаптация обмена.
При содержании животных на высокоуглеводном рационе отмечено повышение уровня глюкозы (у норок на 14,3%, у лисиц – 19,2%) и ферментов углеводного обмена. У норок уровень ЛДГ и α-амилазы в данном случае выше на 21,5% и 15,5% соответственно по сравнению с содержанием на сбалансированном рационе, у лисиц на 32,6% и 11,6%.
П
24
ри высокоуглеводной диете наблюдается снижение уровня щелочной фосфатазы, что связано с уменьшением интенсивности белкового синтеза. У норок отмечены более высокие сдвиги в снижении щелочной фосфатазы по сравнению с лисицами, что составило 15,4%, а у лисиц – 7,5%. Также при содержании зверей на данном рационе отмечено снижение общего билирубина на 19,3% у норок и 16,9% у лисиц. Произошло значительное снижение уровня триглицеридов у норок на 38,5%, у лисиц на 28,6%.
У норок и лисиц, содержащихся на рационе с повышенным уровнем белка, отмечено повышение активности аминотрансфераз при одновременном увеличении содержания общего белка в крови. Увеличение АсТ у норок произошло на 11,4%, у лисиц на 8,1%; АлТ на 9,2% и 5,6% соответственно.
При анализе картины изменения ферментной активности крови лисиц и норок при увеличении жиров в рационе кормления отмечено снижение уровня ЛДГ, очевидно, за счет ингибирования его синтеза. В данном случае уровень ЛДГ у норок составил 969,4 ед/л (18,1%), у лисиц – 900,1 ед/л (15,8%) по сравнению с нормой.
Изменение количества жира в рационе, особенно его избыток, сопровождается изменением активности ряда ферментов крови. В частности увеличение содержания жира в кормовых рационах до 60 % вызывает увеличение трансаминаз, АсТ и АлТ крови при этом возрастают в 1,5 – 2 раза. Более выраженные изменения отмечены у лисиц: уровень АсТ увеличился на 81,2%, АлТ на 93,8%. У норок уровень АсТ составил 190,9 ед/л, что выше нормы на 52,0%; уровень АлТ составил 139,5 ед/л, что на 74,2% выше нормы.
В наших исследованиях при содержании норок и лисиц на диете, содержащей большое количество жиров, отмечено увеличение активности щелочной фосфатазы на 18,5% у норок и 29,7% у лисиц.
При определении активности α-амилазы в случае содержания норок и лисиц на рационе с увеличенной долей жира отмечено снижение активности фермента на 36,2 и 49,7% соответственно (таблица 7).
У
25
величение доли жирных кормов в рационах питания пушных зверей неоправданно, т. к. происходит повышение концентрации общих липидов в крови. Ограничение жизненного пространства (содержание в клетках) способствует нарушению утилизации жиров, что также приводит к гиперлипемии.
Таблица 7 – Биохимические показатели крови норок и лисиц при различных рационах кормления
| Показатели | Рацион | Рацион | ||||||
| Сбалан- сированный | Высоко- углеводный | Высоко- белковый | Высоко- жировой | Сбалан- сированный | Высоко- углеводный | Высоко- белковый | Высоко- жировой | |
| Норки, n=8 | Лисицы, n=10 | |||||||
| Общий белок, г/л | 77,2±6,85 | 70,3±5,30 | 86,8±4,2 | 71,0±5,46 | 65,2±1,19 | 60,4±5,18 | 67,2±4,91 | 65,0±4,76 |
| Общий билирубин, мкмоль/л | 15,0 ±0,32 | 12,1±0,85** | 16,1±1,02 | 17,8±1,04* | 13,7±0,06 | 11,5±0,98* | 12,7±1,03 | 14,9±1,04 |
| Глюкоза, ммоль/л | 9,1±0,51 | 10,4±0,90 | 8,2±0,54 | 8,0±0,5 | 7,3±0,04 | 8,7±0,63* | 7,0±0,40 | 7,0±0,65 |
| Мочевина, ммоль/л | 2,3±0,13 | 2,1±0,32 | 3,4±0,12*** | 2,5±0,13 | 2,3±0,02 | 2,0±0,06 | 2,9±0,09 | 2,6±0,08 |
| Креатинин, ммоль/л | 69,2±2,41 | 68,0±3,41 | 68,3±3,78 | 69,0±4,60 | 70,0±3,40 | 72,0±6,31 | 70,5±6,44 | 70,9±6,74 |
| О  бщие липиды, ммоль/л бщие липиды, ммоль/л | 6,7±0,06 | 5,2±0,36*** | 6,0±0,51 | 7,2±0,65 | 5,4±0,42 | 4,9±0,27 | 5,3±0,30 | 6,1±0,42 |
| Холестерин, ммоль/л | 6,5±0,58 | 6,0±0,41 | 6,6±0,47 | 7,8±0,59 | 4,2±0,18 | 3,9±0,24 | 4,2±0,28 | 5,6±0,38** |
| Триглицериды, ммоль/л | 1,3±0,07 | 0,8±0,4 | 1,3±0,01 | 2,1±0,14*** | 0,7±0,01 | 0,5±0,01 | 0,8±0,03** | 1,24±0,06*** |
| Мочевая кислота, ммоль/л | 48,1±2,54 | 48,0±2,1 | 52,1±2,64 | 48,0±2,17 | 47,0±1,56 | 45,0±3,28 | 52,0±3,85 | 49,0±3,12 |
| АсТ, ед/л | 125,6±8,79 | 130,3±9,36 | 139,9±10,31 | 190,9±12,91*** | 112,0±6,72 | 116,6±7,31 | 121,1±6,39 | 203,2±11,20*** |
| АлТ, ед/л | 80,1±9,35 | 81,7±6,44 | 87,5±5,32 | 139,5±8,46*** | 65,8±8,21 | 67,5±4,48 | 69,5±5,41 | 127,5±7,49*** |
| ЩФ, ед/л | 32,4±1,44 | 27,4±2,08 | 34,7±2,44 | 38,4±3,07 | 24,0±2,35 | 22,2±1,55 | 25,6±1,98 | 29,7±2,31 |
| ЛДГ, ед/л | 1183,7±43,41 | 1438,2±59,61** | 1102,0±48,37 | 969,4±58,41* | 1069,0±10,15 | 1417,5±22,37*** | 1048,0±37,46 | 900,1±41,54*** |
| КК, ед/л | 964,8±21,42 | 958,7±46,32 | 960,0±39,14 | 983,2±57,13 | 909,0±8,71 | 900,2±36,71 | 923,7±19,31 | 944,1±38,70 |
| α-амилаза, ед/л | 468,1±15,67 | 540,5±32,16* | 438,1±31,15 | 298,6±18,49*** | 556,8±32,78 | 621,6±34,13 | 507,2±27,02 | 280,1±14,17*** |
| -ГТФ, ед/л | 436,1±18.92 | 402,0±21,47 | 507,8±23,47* | 491,3±32,11 | 428,7±21,15 | 408,0±28,19 | 496,0±30,19 | 480,7±31,25 |
Примечание - * P≤0,05; ** P≤0,01; *** P≤0,001 в сравнении с показателями при сбалансированном рационе.
Несомненно, приспособление животных к необычным для них видам пищи приводит к изменению химического состава крови, что не во всех случаях является признаком патологии. Следует отметить, что все изменения, происходящие с биохимическими показателями крови, имеют кратковременный характер. При возврате животных на содержание со сбалансированным рационом происходит изменение биохимического профиля крови к референтным величинам. В звероводческих хозяйствах, выращивающих одновременно и американских норок и серебристо-черных лисиц, подход к кормлению не учитывает видовые особенности ферментной системы крови животных. В связи этим, установление референтных величин по всем биохимическим показателям крови, их адаптация к различным условиям, в том числе к различным кормовым добавкам – очень важная задача, требующая дальнейшего изучения.
Оценка уровня стрессированности американских норок
и серебристо-черных лисиц в зависимости от сезонов года
В условиях резкоконтинентального климата Забайкалья, в условиях гиподинамии в силу ограниченности пространства, невозможности укрыться от неблагоприятных условий в зимний период и в моменты высокой солнечной активности летом у животных возникают стрессовые ситуации, вызывающие изменения физиологического статуса.
Выбранные объекты для исследований - американская норка и серебристо-черная лисица - подвергаются одомашниванию в течение сравнительно недолгого времени. Условия клеточного содержания остаются для них ещё непривычными и они сохранили злобность, повышенную возбудимость (Олейник, 1980; Берестов, 2002).
Важную роль в адаптивных реакциях организма играют гормоны коры надпочечников – кортикостероиды, регулирующие процессы метаболизма.
В данной работе представлены результаты по определению уровня кортикостероидных гормонов в крови экспериментальных животных.
С
27
огласно результатам исследований норки отличаются от лисиц более высоким уровнем кортикостероидов. Концентрация кортикостероидов в крови норок, разводимых в клеточных условиях, составила в среднем 28,6 нг/мл, доверительные границы в пределах от 20 до 37,5 нг/мл.
Высокая индивидуальная изменчивость концентрации кортикостероидов отмечена и у лисиц. У лисиц содержание кортикостероидов в крови составило в среднем 20,3 нг/мл. Концентрация гормонов находилась в пределах от 14,8 до 25,8 нг/мл (таблица 8).
В исследованиях была определена зависимость уровня кортикостероидов от климатических условий в различные сезоны года. В частности, мы определяли концентрацию кортикостероидов в январе - самый холодный период года. Температура месяца в среднем в условиях Забайкалья составляет минус 35 – 40°С. В период высокой аридности и активной солнечной инсоляции в июле плюс 33-41°С.
Таблица 8 - Уровень кортикостероидных гормонов в крови норок и лисиц, нг/мл (M±m)
| Виды животных | Норма | Зима (январь) | Лето (июль) |
| Норки, n=8 | 28,6±1,31 | 39,7±2,96 (138,8%)** | 30,7±1,89 (107,3%) |
| Лисицы, n=10 | 20,3±0,87 | 25,1±1,57 (123,6%)* | 25,9±2,01 (127,6%)* |
Примечания
1 В % указаны отклонения показателей от нормы;
2 * P≤0,05; ** P≤0,01; *** P≤0,001 в сравнении с нормой.
В зимний период уровень кортикостероидных гормонов норок значительно повышается по сравнению с таковым у лисиц. Низкая температура окружающей среды являются стресс-фактором для норок, хотя предки клеточных норок – это выходцы из Северной Америки, климатические условия которой достаточно суровы. Но постоянное нахождение норок в клетках без возможности укрыться от неблагоприятных температур, очевидно, способствует увеличению кортикостероидов в крови животных.
П
28
о-видимому, для обеспечения более высокого уровня метаболизма у норок при понижении температуры окружающей среды усиливается функциональная активность надпочечников. Согласно исследованиям И.Л. Туманова (2003) у норок наблюдается более высокий уровень газообмена зимой по сравнению с полярными лисицами и песцами, что, очевидно, требует и более высокого уровня гормональной активности в зимний период.
При определении концентрации кортикостероидов у норок и лисиц в июле выявилась несколько иная картина по сравнению с зимой. Большее повышение концентрации кортикостероидов наблюдалось у лисиц, чем у норок. Уровень кортикостероидов у норок в крови в летний период составил 30,7 нг/мл. У лисиц отмечена концентрация гормонов в крови на уровне 25,9 нг/мл.
Таким образом, на основании полученных данных можно судить о видовой специфике активности надпочечников у норок и лисиц, которая проявляется в более высоком уровне кортикостероидных гормонов у норок. При понижении температуры окружающей среды кортикостероидная активность усиливается в большей степени у норок по сравнению с лисицами. Лисицы – это обитатели высоких широт, тогда как норки относятся к животным умеренных широт, в связи с этим при значительном понижении температуры воздуха, которая является стрессовым фактором, происходит повышение концентрации кортикостероидных гормонов в крови, более выраженное у норок. Разное соотношение кортикостероидной активности норок и лисиц отражает различный уровень их адаптации к сезонам года.
Влияние различных рационов кормления
на уровень кортикостероидных гормонов
у американских норок и серебристо-черных лисиц
В исследованиях была предпринята попытка выяснить зависимость концентрации кортикостероидов в крови у норок и лисиц от различных рационов питания, которые были предложены в экспериментах по изучению внешнесекреторной деятельности поджелудочной железы у экспериментальных животных.
П
29
ри содержании на рационах с увеличенной долей углеводов у норок отмечено повышение уровня кортикостероидных гормонов на 31,2%. У лисиц при содержании на указанном рационе наблюдалось не столь значительное повышение и уровень кортикостероидных гормонов в среднем составил 23,1 нг/мл, что выше уровня нормы на 13,7% (рисунок 6).
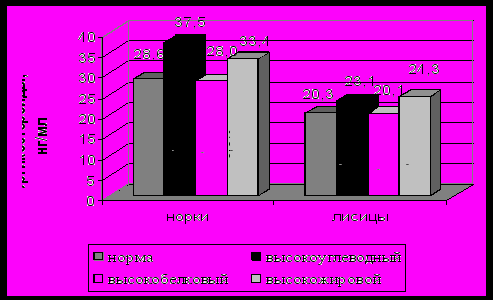
Рисунок 6 - Зависимость уровня кортикостероидных гормонов от рационов кормления норок и лисиц.
Увеличение доли белковых кормов в рационе экспериментальных животных не выявило достоверных изменений в уровне кортикостероидных гормонов.
Избыточное количество жира в кормовом рационе способствует изменению концентрации кортикостероидов в крови. Эти изменения в большей степени проявляются у лисиц. Уровень кортикостероидов в крови лисиц при содержании на высокожировом рационе составил в среднем 24,3 нг/мл. У норок отмечена концентрация кортикостероидов в крови на уровне 33,4 нг/мл.
Таким образом, изменение качественного состава рационов кормления способствует повышению уровня кортикостероидных гормонов в крови. На основании результатов исследований можно сделать вывод о том, что изменение кормовой базы является достаточно сильной стрессовой ситуацией для животных и длительное содержание на несбалансированных кормах приводит к снижению продуктивности животных, что у норок и лисиц проявляется в качестве меховой продукции.
30
Влияние рационов кормления на меховую продукцию экспериментальных животных
В исследованиях проведен анализ кормления норок и лисиц в период формирования зимнего волосяного покрова (июль-октябрь) с учетом изменения рациона. Товарные качества шкурки определяются в первую очередь развитием волосяного покрова в целом, а также их количественным соотношением. Вариации в качестве волосяного покрова могут зависеть от многих причин, знание которых необходимо для получения продукции высокого качества. Волосяной покров норок и лисиц имеет видовые отличия, что обусловлено в первую очередь образом жизни их диких предков в процессе эволюции.
Согласно результатам исследований длина шкурки после убоя животных, содержащихся на углеводной диете, была меньше, чем у животных на сбалансированном рационе: у норок на 11,8%, у серебристо-черных лисиц – 10,6%.
При содержании норок и лисиц на кормах с большим содержанием углеводов наблюдается разреженность волоса, кроющие волосы не достигают нормальной длины. У норок и лисиц при высокоуглеводном рационе длина направляющих волос на 17,9% и 11,5% меньше длины волос, чем при содержании на сбалансированном рационе. Длина остевых волос короче на 23,2% и 17,3%. На данном рационе также уменьшилась длина пуховых волос на 26,8% и 8,7% соответственно (таблица 9).
Количество направляющих волос сократилось у норок на 8,8%, у лисиц на 20,0%, остевых - у норок 8,0%, лисиц - 48,4%, пуховых – у норок на 1,8%, у лисиц на 5,3%. За счет сильного укорочения кроющих волос и изменения количественного соотношения направляющих, остевых и пуховых волос у лисиц и норок при содержании на углеводных кормах был получен дефект просвечивающей подпуши, что приводит к снижению ценовой категории меховой продукции.
31
Таблица 9 - Качественные показатели меха норок и лисиц
| Показатели меха | Рацион кормления | ||||
| Сбаланси-рованный | Высоко-углеводный | Высоко-белковый | Высоко-жировой | ||
| Норки (n=8) | |||||
| Длина шкурки, см | 58,5±1,23 | 51,6±1,65 (88,2%)** | 66,7±2,35 (114,0%)** | 62,3±1,89 (106,5%) | |
| Длина волос, мм | направ. | 29,6±1,19 | 24,3±1,95 (82,1%)* | 33,2±2,01 (112,2%) | 30,1±2,25 (101,7%) |
| остевые | 26,7±0,86 | 20,5±1,81 (76,8%)** | 30,7±2,12 (115,0%) | 28,3±1,68 (106,0%) | |
| пуховые | 21,3±1,23 | 15,6±1,24 (73,2%)** | 21,5±0,96 (101,0%) | 18,7±1,37 (87,8%) | |
| Коэф. мягкости | 1,3∙10-3 | 1,3∙10-3 | 1,8∙10-3 | 1,4∙10-3 | |
| Лисицы (n=10) | |||||
| Длина шкурки, см | 95,8±6,37 | 85,6±5,49 (89,4%) | 102,7±7,29 (107,1%) | 97,3±7,1 (101,6%) | |
| Длина волос, мм | направ. | 77,1±0,74 | 68,2±3,21 (88,5%) | 81,5±4,61 (105,7%) | 80,2±3,31 (104,0%) |
| остевые | 62,5±0,86 | 51,7±2,44 (82,7%)*** | 63,9±1,56 (102,2%) | 61,4±3,12 (98,2%) | |
| пуховые | 46,3±1,21 | 42,3±1,83 (91,3%) | 51,2±0,91 (110,6%)** | 53,5±2,54 (115,6%)* | |
| Коэф. мягкости | 1,9∙10-3 | 1,8∙10-3 | 2,0∙10-3 | 1,3∙10-3 | |
Примечания
1 В % указаны отклонения от показателей при сбалансированном рационе;
2 * P≤0,05; ** P≤0,01; *** P≤0,001 в сравнении со сбалансированным рационом.
При содержании животных на кормах, богатых протеинами, выявлена следующая картина. Средняя длина направляющих волос превышает показатели при обычном рационе на 12,2% у норок и 5,7% у лисиц. Длина ости у норок при высокобелковом рационе кормления превышает показатели при сбалансированном рационе у норок на 15,0%, у лисиц – 2,2%. По длине пуховых волос изменения у норок незначительные и составляют 1,0%. Средняя длина пуховых волос у лисиц при повышенном протеиновом питании превосходит показатели при обычном рационе на 10,6%.
В
32
количественном соотношении волос на данном рационе отмечены незначительные изменения. У норок плотность волос по всем категориям на единицу площади увеличилась: направляющих на 3,3%, остевых на 9,5%, пуховых – 5,2%. У лисиц – направляющих на 3,6%, пуховых на 2,8%. По количеству остевых волос у лисиц также отмечены незначительные изменения.
По длине шкурковой продукции при содержании на повышенном протеиновом питании у норок отмечено увеличение показателей по сравнению со сбалансированным рационом на 14,0%. У лисиц эти изменения составили 7,1% в сторону увеличения.
Несмотря на увеличение длины волос всех категорий у исследованных животных при белковом рационе отмечено раннее созревание кроющих волос (конец августа – начало сентября), ость тусклая, местами бурого цвета (у лисиц) и к началу зимы отмечены секущиеся концы. Кроме того, коэффициент мягкости меха у исследованных животных изменился в сторону жесткости и составил у норок 1,8∙10-3 , у лисиц 2,0∙10-3.
При увеличении доли жира в кормовом рационе в период активного созревания меха у норок выявлено незначительное увеличение длины шкурки на 6,5%, удлинение направляющих и остевых волос на 1,7% и 6,0% соответственно. Но при этом происходит значительное уменьшение длины пухового волоса на 12,2%, что, несомненно, сказывается на тепловых свойствах меха. По количеству основных категорий волос отмечено их незначительное сокращение (от 0,6 до 1,1%). Кроме того, при содержании норок на жировом рационе наблюдали «неуравненность» меха.
У лисиц при содержании на высокожировом рационе отмечены снижение уровня серебристости и желтизна волос. Выявлены незначительные изменения по количеству волос на единицу площади шкурки: число направляющих волос сократилось на 2,4%, остевых – 6,8%, пуховых – 1,7%. Наблюдалось значительное и достоверное увеличение длины пуховых волос на 15,6% (P≤0,05).
Т
33
аким образом, увеличение протеинового и жирового кормления приводит не только к удорожанию рациона, но и к снижению качества меховой продукции. Для снижения потерь на качестве пушнины необходимо придерживаться полноценного сбалансированного рациона питания пушных зверей, так как кардинальное изменение традиционной кормовой базы приводит к дефектам шкурковой продукции.