
Физиологическая адаптация американских норок (Mustela vizon schr.) и серебристо-черных лисиц (vulpes vulpes L.) при клеточном содержании в условиях забайкалья 03. 03. 01 физиология
| Вид материала | Автореферат |
- Понятие адаптации, 125.13kb.
- Физиологическая характеристика оздоровительных и экстремальных видов спортивной деятельности, 417.86kb.
- Доклад на тему: Топография мышц плечевого пояса методом обычного тонкого препарирования, 53.76kb.
- Вопросы адаптации детей к школе, 133.74kb.
- Коц Я. М. Спортивная физиология. Учебник для институтов физической культуры, 4839.54kb.
- Н. П. Пахомов 1971г. Предисловие Н. П. Пахомов старейший знаток гончих собак и охоты, 917.03kb.
- Рассказ о северском донце, 201.76kb.
- Физиологическая роль сосудистых рефлексогенных зон в интегративной регуляции функций, 653.7kb.
- Физиология желез внутренней секреции, 15.13kb.
- Курсовая работа Адаптация иммигрантов-мусульман в условиях современной России: проблемы, 166.14kb.
Сравнительный анализ функциональной деятельности поджелудочной железы у американских норок и серебристо-черных лисиц
Для получения данных в сравнительном аспекте проведены экспериментальные исследования по изучению активности ферментов гомогената ткани поджелудочной железы норок и лисиц (таблица 2).
Таблица 2 - Активность ферментов в гомогенате ткани поджелудочной железы пушных зверей, M±m
| Вид животных | n | Амилаза мг/г.мин | Протеазы мг/г.мин | Липаза мкмоль/г.мин | Протеазно- амилазное соотношение |
| Норки | 22 | 851±7,6 | 405±3,9 | 14±0,2 | 1 : 2,1 |
| Лисицы | 24 | 676±6,2 | 234±1,9 | 11±0,1 | 1 : 2,8 |
А
12
ктивность амилазы в гомогенате поджелудочной железы у лисиц составила 676±6,2 мг/г.мин, у норок - 851±7,6 мг/г.мин. Общая протеолитическая активность гомогената хищных пушных зверей имела параллельные изменения по сравнению с амилолитической активностью. Концентрация протеаз у норок составила 405±3,9; у лисиц 234±1,9 мг/г.мин. Липаза имеет такое же распределение как амилаза и протеазы. Активность липазы в среднем у норок составляет 14,0±0,2; у лисиц - 11±0,1 мкмоль/г.мин.
Сравнивая результаты исследований активности ферментов в панкреатическом соке и гомогенате ткани поджелудочной железы, следует отметить незначительные различия в показателях. Отличия активности амилазы в панкреатическом соке и в гомогенате ткани поджелудочной железы лисиц составили 2,7%, протеаз – 3,1%, липазы – 5,8%.
Эволюционная адаптация поджелудочной железы к качеству питания исследованных животных проявляется при сопоставлении соотношения протеаз к амилолитической активности. По результатам исследований протеазно-амилазное соотношение у норок составляет 1:2,1; у лисиц 1:2,8. При изучении экзокринной функции поджелудочной железы лисиц протеазно-амилазное соотношение в панкреатическом соке составило 1:2,9.
Анализ данных показал, что метод определения активности ферментов в гомогенате ткани поджелудочной железы является достоверным и может использоваться при изучении данной функции у животных, применение фистул у которых затруднено. По данным P. Kohishevski (1973), проводившего исследования активности ферментов поджелудочной железы европейской норки дикой популяции, соотношение протеолитической активности к амилолитической составляет 1:2. Тогда как у норок клеточного содержания, в рационе которых в большей степени присутствуют растительные корма, это соотношение составляет 1:2,1. Несмотря на различные методы, использованные в исследованиях P. Kohishevski и в наших опытах, протеазно-амилазное соотношение у европейской норки дикой популяции и американской норки клеточного разведения приблизительно одинаковы.
Д
13
ля растительноядных и всеядных видов характерна адаптивная диссоциация пищеварительных ферментов, их организм генетически запрограммирован на потребление самой разнообразной пищи (Уголев, 1972, 1980). По данным В.М. Олейник (1984) у кроликов, как представителей грызунов, протеазно-амилазное соотношение в поджелудочной железе составляет 1:21,2; у всеядных крыс 1:11,5. Согласно нашим ранним исследованиям (Санжиева, 1993) у белок данное соотношение составляет 1:20, ондатр 1:34. По результатам исследований С.Н. Сафаровой (1977) у нутрий отношение амилолитической активности к общему количеству протеаз составляет 1:290.
На рисунке 1 в сравнительном аспекте представлены результаты наших экспериментов и других исследователей, выполненные под руководством Ц.Ж. Батоева.
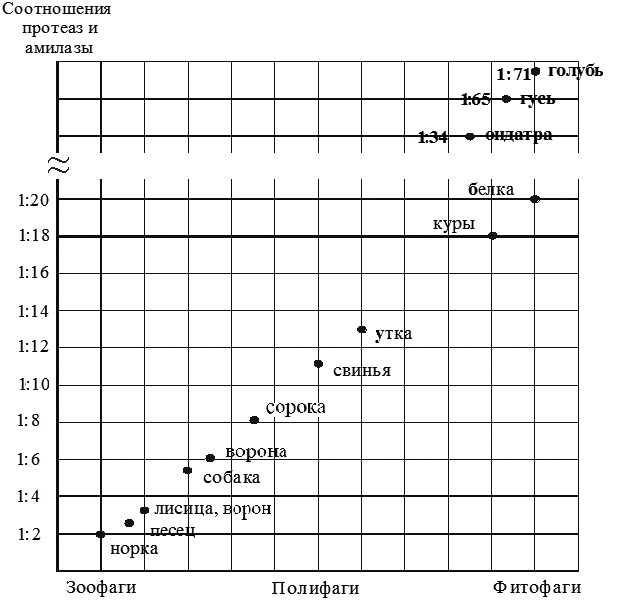
Рисунок 1 - Соотношения активности протеаз и амилазы поджелудочной железы в связи с типами питания разных видов животных и птиц.
Примечание - Куры, утки, гуси (исследования Батоева ); собаки (Цыбекмитовой); свиньи (Алиева); норки, песцы, лисицы, белки, ондатры (Санжиевой.); голуби (Аюрзанаевой); черная ворона, ворон (Содбоевой); сорока (Баниевой).
Н
14
орка – типичный представитель плотоядного питания, из-за самого низкого соотношения протеаз и амилазы в ткани поджелудочной железы занимает крайнюю позицию на кривой. Верхняя граница у лисиц стыкуется и перекрывается с показателями у собак.
Таким образом, учитывая принадлежность исследованных животных к разным семействам куньих и собачьих отряда хищные, а также их различную экологическую специализацию в прошлом, можно считать, что отличия в активности ферментов являются видовыми особенностями данных зверей. То, что при разведении в клетках в одинаковых климатогеографических условиях, чрезвычайно отличающихся от естественной среды обитания, различия в соотношении ферментов поджелудочной железы не стерлись, свидетельствует о том, что процесс доместикации пушных зверей еще не закончился.
Статус пищеварительных ферментов поджелудочной железы у американских норок и серебристо-черных лисиц
Для исследования влияния диеты опытных норок и лисиц содержали на рационах с повышенным уровнем углеводов (50%), жира (60%) и белка (80%) в течение определенного срока (июль-октябрь). В контрольных опытах животные получали сбалансированный рацион с содержанием переваримых веществ: белок – 50-60%, жир -20-35%, углеводы -15-20% от общей энергии кормосмеси.
Результаты опытов свидетельствуют, что изменение активности панкреатических ферментов под влиянием диеты в большинстве случаев не слишком велико и главное не является однонаправленным.
Заметных сдвигов в активности ферментов поджелудочной железы под влиянием белкового, углеводного и жирового рационов у норок и лисиц не наблюдалось. Только в отношении липазы выявлена тенденция к повышению активности фермента у животных на высокожировой диете, у норок на 8,6%, у лисиц на 10,7%.
15
Известно, что у всеядных крыс при смене диеты уже через 7 дней наблюдается отчетливая адаптивная диссоциация панкреатических ферментов (Уголев, Груздков, Егорова, 1981; Corring, 1980). Однако у американских норок и серебристо-черных лисиц подобные изменения не отмечены (таблица 3).
Таблица 3 - Влияние состава кормов на активность пищеварительных ферментов поджелудочной железы норок и лисиц, M±m
| Фермент | Рационы | |||
| | Сбалан- сированный | Высоко-углеводный | Высоко-белковый | Высоко-жировой |
| Норки (n=10) | ||||
| Амилаза, мг/ мл.мин. | 851±7,6 | 872±8,1 (102,5%) | 857±7,2 (100,7%) | 881±8,5 (103,5%)* |
| Протеазы, мг/ мл.мин. | 405±3,9 | 413±3,2 (101,9%) | 417±3,7 (103,0%)* | 418±2,8 (103,2%)* |
| Липаза, мкмоль/ мл.мин. | 14,0±0,71 | 14,3±1,25 (102,1%) | 14,1±1,13 (100,7%) | 15,2±1,16 (108,6%) |
| Лисицы (n=10) | ||||
| Амилаза, мг/ мл.мин. | 676±6,2 | 700±4,7 (103,6%)** | 673±3,0 (99,7%) | 715±4,3 (105,8%)*** |
| Протеазы, мг/ мл.мин. | 234±1,9 | 238±4,0 (101,6%) | 232±2,0 (99,4%) | 244±2,2 (104,4%)** |
| Липаза, мкмоль/ мл.мин. | 11,0±0,13 | 11,4±0,16 (103,5%) | 10,6±0,14 (96,4%) | 12,2±0,15 (110,7%)*** |
Примечания
1 В % указаны отклонения концентраций ферментов от их концентрации при сбалансированном рационе;
2 *P≤0,05; **P≤0,01; ***P≤0,001 в сравнении с группой животных на сбалансированном рационе.
Результаты опытов подтверждают наше предположение об отсутствии у хищников типичной для растительноядных и всеядных животных адаптивной диссоциации панкреатических ферментов в ответ на качественное изменение рациона. Эти особенности отражают эволюционную специализацию пищеварительной системы хищников к лучшему использованию белка и слабой ее способности к утилизации углеводов. Отсутствие адаптивной диссоциации панкреатических ферментов на качественное изменение рациона, по-видимому, генетически детерминированный признак.
Особенности биохимического профиля крови
американских норок и серебристо-черных лисиц
И
16
спользование биохимических методов для постоянного наблюдения за физиологическим состоянием американских норок и серебристо-черных лисиц в сравнительном аспекте является весьма перспективным.
В ходе экспериментальных исследований впервые определены нормы биохимических показателей крови норок и лисиц в условиях Забайкалья, представленные в таблице 4.
Таблица 4 - Биохимические показатели крови норок и лисиц в норме, М±m
| Показатели | Норки, n=8 | Лисицы, n=10 |
| Общий белок, г/л | 77,2±6,85 | 65,2±1,19 |
| Общий билирубин, мкмоль/л | 15,0 ±0,32 | 13,7±0,06 |
| Глюкоза, ммоль/л | 9,1±0,51 | 7,3±0,04 |
| Мочевина, ммоль/л | 2,3±0,13 | 2,3±0,02 |
| Креатинин, ммоль/л | 69,2±2,41 | 70,0±3,40 |
| Общие липиды, ммоль/л | 6,7±0,06 | 5,4±0,42 |
| Холестерин, ммоль/л | 6,5±0,58 | 4,2±0,18 |
| Триглицериды, ммоль/л | 1,3±0,07 | 0,7±0,01 |
| Мочевая кислота, ммоль/л | 48,1±2,54 | 47,0±1,56 |
В биохимической картине крови исследованных животных отмечаются межвидовые различия, которые заключаются в более высоком содержании у норок общего белка на 15,5%, билирубина – 8,7%, глюкозы – 19,8%, общих липидов – 19,4%, холестерина – 35,4% и триглицеридов – 46,2%.
Анализ экспериментальных данных свидетельствует о заметном влиянии сезонного фактора на биохимические показатели и ферментную активность крови у норок и лисиц, что выражается в определенной направленности и различной степени изменений физиологических показателей в течение года (таблица 5).
Анализируя таблицу, следует отметить, что значительное и достоверное снижение (P≤0,05; P≤0,01) биохимических показателей крови норок в летний период происходит в уровне общего белка, глюкозы, мочевины, холестерина и триглицеридов. В этот же период отмечено достоверное повышение (Р≤0,01) общих липидов в крови норок на 36,2%.
У
17
лисиц в летний период выявлено достоверное уменьшение содержания глюкозы на 29,3% (Р≤0,001) и холестерина на 32,0% (Р≤0,01). В летний период у лисиц также отмечено достоверное повышение уровня общих липидов на 24,5% (Р≤0,05).
У норок уровень белка в крови выше, чем у лисиц, как в среднем по показателям в норме, так и по сезонам года. Очевидно, это видовая особенность интенсивности обмена белка у норок.
Таблица 5 - Биохимические показатели крови норок и лисиц в зимне-летний период, M±m
| Показатели | Норки, n=8 | Лето в % к зиме | Лисицы, n=10 | Лето в % к зиме | ||
| зима | лето | зима | лето | |||
| Общий белок, г/л | 89,8±5.31 | 76,2±1,19 | 84,9* | 72,5±5,81 | 63,8±4,31 | 88,0 |
| Общ.билирубин, мкмоль/л | 16,0 ±0,95 | 14,8±0,87 | 92,5 | 14,0 ±0,67 | 13,8±0,08 | 98,6 |
| Глюкоза, ммоль/л | 10.8±0,91 | 6,9±0,51 | 63,9** | 8,2±3,46 | 5,8±0,28 | 70,7*** |
| Мочевина, ммоль/л | 2,9±0,19 | 2,3±0,02 | 79,6** | 2,5±0,17 | 2,2±0,01 | 88,0 |
| Креатинин, ммоль/л | 68,7±4,25 | 69,0±5,41 | 100,4 | 69,0±2,41 | 68,5±4,45 | 99,3 |
| Общие липиды, ммоль/л | 5,8±0,47 | 7,9±0,42 | 136,2** | 4,9±0,32 | 6,1±0,48 | 124,5* |
| Холестерин, ммоль/л | 7,9±0,68 | 5,1±0,32 | 64,6** | 5,0±0,37 | 3,4±0,19 | 68,0** |
| Триглицериды, ммоль/л | 1,5±0,06 | 1,2±0,08 | 79,5** | 0,9±0,02 | 0,7±0,01 | 77,8 |
| Мочевая кислота, ммоль/л | 48,0±3,52 | 49,0±2,46 | 102,1 | 47,5±2,48 | 48,0±2,01 | 101,1 |
Примечание - * P≤0,05; ** P≤0,01; *** P≤0,001.
Из числа показателей углеводного и жирового обмена у пушных зверей наиболее важными являются уровень гликемии, содержание триглицеридов и холестерина. В норме уровень гликемии у различных видов животных колеблется. Сведения о содержании сахара в крови пушных зверей в научной литературе фрагментарны. В работах В.А. Берестова (1971, 1981) приводятся данные содержания глюкозы у стандартных норок (177 мг%) и серебристо-черных лисиц (125 мг%) в условиях Карелии.
В
18
наших исследованиях данный показатель определяли у норок и лисиц в условиях Забайкалья по сезонам года и в зависимости от рациона питания.
Согласно результатам исследований уровень гликемии у норок составил в среднем 9,1 ммоль/л, у лисиц – 7,3 ммоль/л. При использовании коэффициента пересчета ммоль/л в мг%, равный 0,0555, нами получены данные по содержанию глюкозы у норок - 165,5 мг%, у лисиц - 131,8 мг%. Сезонный диморфизм уровня гликемии у норок и лисиц имеет параллельные изменения, которые характеризуются постепенным уменьшением концентрации глюкозы в крови летом и увеличением к зимнему периоду с 9,1 до 10,8 ммоль/л у норок и с 7,3 до 8,2 ммоль/л у лисиц. Очевидно, это связано с сезонными изменениями характера обмена веществ у животных, эволюционно сложившимися к условиям внешней среды, среди которых ведущее и решающее значение имеет питание, поскольку в естественных условиях звери зимой вынуждены переносить большие кормовые лишения, в силу чего они приобрели способность снижать осенью, при обилии пищи в природе, интенсивность обмена веществ и тем самым создавать в организме запасы питательных веществ для использования их в менее благоприятное время года (Берестов, 2002; Туманов, 2003).
Согласно результатам исследований в уровне липемии также отмечаются межвидовые различия, которые заключаются в более высоком содержании общих липидов в плазме крови у норок по сравнению с лисицами. Уровень общих липидов у норок находится в пределах от 6,3 до 7,1 ммоль/л, у лисиц данный показатель составил 4,9 - 5,9 ммоль/л.
При определении уровня общих липидов в крови экспериментальных животных по сезонам года выявлена следующая картина. Содержание липидов повышается в летний период по сравнению с нормой в среднем у норок на 17,9%, у лисиц на 12,9%. Это объясняется процессами активного запасания липидов в летний период. Клеточные звери сохранили динамический стереотип диких предков и запасание липидов в организме в период достаточного количества корма летом, в отличие от зимних месяцев - генетически детерминировано.
С
19
нижение уровня липидов (у норок на 13,4%; у лисиц на 9,3%) в крови зимой по сравнению с нормой у экспериментальных животных свидетельствует об активном расщеплении жиров для энергетического обеспечения организма при повышенной теплопродукции в зимний период.
Сравнивая полученные результаты по уровню холестерина у норок и лисиц, следует отметить более высокие показатели у первых как в среднем, так и по сезонам года. У норок в норме уровень холестерина составил 6,5 ммоль/л, а в январе - 7,9 ммоль/л; в июле - 5,1 ммоль/л. У лисиц уровень холестерина составил в среднем 4,2 ммоль/л. В зимний период у лисиц также отмечается некоторое повышение холестерина в сыворотке крови до 5,0 ммоль/л и снижение летом до уровня 3,4 ммоль/л. Анализ экспериментальных данных свидетельствует о заметном влиянии сезонного фактора на биохимические показатели и ферментную активность крови у норок и лисиц, что выражается в определенной направленности и различной степени изменений физиологических показателей в течение года.
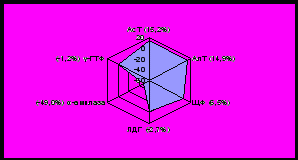
У норок установлено значительное повышение общего белка, триглицеридов, холестерина, мочевины, глюкозы; снижение общих липидов в зимний период по сравнению с нормой. Летом происходит снижение общего белка, триглицеридов, мочевины практически до исходного уровня - нормы, за исключением холестерина и глюкозы, отличия которых ниже нормы на 21,5% и 24,1% соответственно, при этом наблюдается значительное повышение уровня общих липидов (рисунок 2).
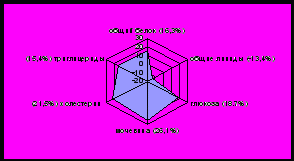 |  |
| январь | июль |
Рисунок 2 - Отклонения от нормы биохимических показателей крови норок, %
У
20
лисиц изменения в уровне общего белка, общих липидов, холестерина, мочевины и глюкозы менее выражены по сравнению с таковыми показателями у норок. В летний период у лисиц также наблюдается значительное снижение уровня глюкозы и холестерина (рисунок 3).
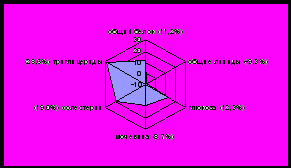 |  |
| январь | июль |
Рисунок 3 - Отклонения от нормы биохимических показателей крови лисиц, %
По данным гистограмм следует, что более существенные изменения с биохимическими показателями крови у норок происходят в зимний период, очевидно суровые климатические условия Забайкалья предъявляют повышенные требования к организму животных. А в летний период изменения незначительны и практически соответствуют норме, кроме показателей холестерина, глюкозы и общих липидов. Уровень сдвигов биохимических показателей крови в зимний период у норок более выражен по сравнению с лисицами. В летний период изменения биохимических показателей крови норок и лисиц имеют однонаправленный характер.
В связи с этим особый интерес представляет изучение активности ферментов крови, характеризующих различные пути метаболизма. К ним в первую очередь относятся трансаминазы, играющие большую роль в регуляции обмена аминокислот. Данные по функциональному состоянию ферментной системы крови норок и лисиц в норме и по сезонам года представлены в таблице 6.
В
21
активности ферментов крови также отмечены видовые различия, которые проявляются в большем содержании АсТ, АлТ, щелочной фосфатазы, ЛДГ, креатинкиназы у норок. У лисиц отмечено более высокое содержание α-амилазы.
Таблица 6 - Активность ферментов крови норок и лисиц в норме и по сезонам года, М±m
| Показатели | Норма | Зима | Лето | Лето к зиме в % |
| Норки, n=8 | ||||
| АсТ, ед/л | 125,6±8,79 | 114,3±7,46 | 144,7±9,32 | 126,6* |
| АлТ, ед/л | 80,1±5,35 | 70,4±4,32 | 92,1±6,41 | 130,8* |
| ЩФ, ед/л | 32,4±1,44 | 24,3±2,01** | 34,5±2,44 | 142,0 |
| ЛДГ, ед/л | 1183,7±43,41 | 1392,1±68,97* | 1152,0±84,47 | 82,8* |
| КК, ед/л | 964,8±21,42 | 960,0±45,26 | 956,7±66,33 | 99,6 |
| α-амилаза, ед/л | 468,1±15,67 | 530,6±12,17** | 238,7±19,00*** | 45,0*** |
| -ГТФ, ед/л | 436,1±18.92 | 497,2±15,69* | 430,8±10,56 | 86,6** |
| Лисицы, n=10 | ||||
| АсТ, ед/л | 112,0±6,72 | 105,1±8,22 | 129,3±7,14 | 123,0* |
| АлТ, ед/л | 65,8±4,21 | 60,6±4,61 | 74,5±6,20 | 122,9 |
| ЩФ, ед/л | 24,0±2,35 | 27,0±1,78 | 25,0±1,18 | 92,6 |
| ЛДГ, ед/л | 1069,0±90,15 | 1187,2±88,5 | 898,3±70,56 | 75,7 |
| КК, ед/л | 909,0±8,71 | 900,5±64,33 | 908,0±78,23 | 100,8 |
| α-амилаза, ед/л | 556,8±32,78 | 602,7±15,46 | 542,3±11,24 | 90,0** |
| -ГТФ, ед/л | 428,7±21,15 | 506,8±13,01** | 430,1±25,63 | 84,9* |
Примечание – * P≤0,05; ** P≤0,01; *** P≤0,001 в сравнении с нормой.
Аминотрансферазы играют ключевую роль в промежуточном обмене, т.к. обеспечивают синтез и разрушение отдельных аминокислот в организме. АсТ и АлТ у норок и лисиц в зимний период, согласно нашим исследованиям, обладают самой низкой активностью по сравнению другими периодами года. К лету их активность нарастает, преимущественно у АсТ, и в июле увеличивается по сравнению с январем у норок на 26,6%, у серебристо-черных лисиц на 23,0%. Высокая активность трансаминаз в летний период свидетельствует о большой способности тканей к синтезу собственных специфических белков за счет активного окислительного расщепления экзогенных аминокислот, интенсивно поступающих с пищей.
С
22
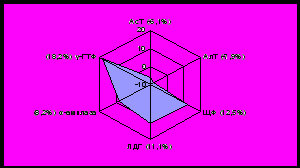
одержание АсТ и АлТ у норок зимой снижается на 9,0% и 12,1%, у лисиц на 6,1% и 7,9% соответственно. Наблюдаемые изменения хорошо согласуются с данными об особенностях белкового обмена, который у норок и лисиц достигает максимальной величины летом, а зимой снижается (рисунок 4,5).
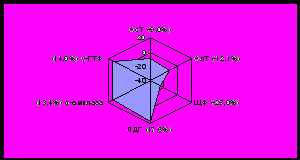 |  |
| январь | июль |
Рисунок 4 - Отклонения активности ферментов крови норок от нормы, %
 |  |
| январь | июль |
Рисунок 5 - Отклонения активности ферментов крови лисиц от нормы, %
Активность щелочной фосфатазы в зимний период у норок снижается на 25,0%. С наступлением летнего периода её уровень незначительно повышается и достигает максимальных значений, что объясняется запасанием белка в данный период.
При снижении ферментов белкового обмена в зимний период у экспериментальных животных наблюдается увеличение уровня ферментов углеводного обмена ЛДГ и α-амилазы, в большей степени выраженное у норок. Повышение уровня данных ферментов свидетельствует о возрастающей роли гликолиза в энергетическом обеспечении организма. В летний период отмечено значительное снижение активности α-амилазы у норок на 49,0% и ЛДГ у лисиц на 16,0%.
Изменение активности АcТ, АлТ, ЩФ, ЛДГ, α-амилазы является отражением особенностей обмена веществ в разные сезоны года.
А
23
нализируя полученные результаты, следует отметить, что пушные звери отличаются от многих домашних животных более высоким уровнем биохимических показателей крови. Им присущи также сезонные изменения биохимического состава крови, заключающиеся в их увеличении в зимний период и снижении летом. Выявленное нами более высокое, по сравнению с лисицами, содержание в крови норок общего белка, сахара, холестерина, липидов и ферментов, является убедительным доказательством высокой интенсивности обменных процессов в организме животных данного вида.
Установление доверительных границ биохимического статуса крови пушных зверей позволяет выявить адаптационные возможности организма. Сравнительно высокие результаты изученных показателей могут быть обусловлены высоким мышечным напряжением представителей данных видов. Процесс метаболизма и его ферментные реакции подвержены сезонным перестройкам, что способствует адаптации клеточных норок и лисиц к крайне суровым климатическим условиям Забайкалья.