
Роль -эндорфина в нейроэндокринной регуляции функций иммунной системы 14. 00. 36 Аллергология и иммунология
| Вид материала | Автореферат диссертации |
- Примерная программа дисциплины иммунология. Клиническая иммунология для студентов обучающихся, 327.77kb.
- Патофизиология эндокринной системы, 226.95kb.
- Образовательный стандарт послевузовской профессиональной подготовки специалистов Специальность:, 1035.98kb.
- Физиологическая роль сосудистых рефлексогенных зон в интегративной регуляции функций, 653.7kb.
- Программа курса "Иммунология", 85.71kb.
- Расписание занятий на элективе для студентов 6 курса «Клиническая аллергология», 41.99kb.
- Хроническая крапивница: этиопатогенетическая роль инфекции Helicobacter pylori и разработка, 739.8kb.
- «Оренбургская государственная медицинская академия Федерального агентства по здравоохранению, 433.12kb.
- Название работы, 475.33kb.
- Тематический план цикла по специальности "аллергология и иммунология", 39.23kb.
Примечание. Здесь и в табл. 6: * - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю по парному t-критерию Фишера наименьшей значимой разницы, a – р <0,05 – к спонтанной продукции.
стимулируют продукцию IL-1ra по сравнению с контролем как на 1-е, так и на 2-е сутки культивирования. Отмены стимулирующего влияния пептида на фоне блокады опиатных рецепторов не наблюдается. Анализ влияния пептида на продукцию IL-8 показал статистически достоверный угнетающий эффект как на 24, так и на 48 ч культивирования. В условиях блокады опиатных рецепторов налоксоном и налтриндолом угнетающее влияние -эндорфина нивелируется. Внесение налоксона и налтриндола на продукцию IL-8 не влияет.
Таблица 6. Влияние b-эндорфина на продукцию IL-1, TNF-, IL-6 в очищенной фракции моноцитов
| Цитокин, | Экспериментальное | Концентрация b-эндорфина, M | |||
| пг/мл | воздействие | контроль | 10-7 | 10-9 | 10-11 |
| | Без индуктора | 153,22 | 121,65 | 249,92 | 118,71 |
| IL-1, | | 33,48 | 19,83 | 84,88 | 28,97 |
| n=8 | ЛПС 0,1 мкг/мл | 230,26 | 202,84 | 223,97 | 235,21 |
| | | 42,25а | 36,47 | 71,16 | 53,29 |
| | Без индуктора | 137,52 | 153,81 | 148,31 | 133,20 |
| TNF-, | | 51,33 | 58,24 | 57,56 | 55,31 |
| n=8 | ЛПС 0,1 мкг/мл | 163,21 | 178,81 | 174,97 | 178,73 |
| | | 60,48а | 69,78 | 68,63 | 70,21 |
| | Без индуктора | 1204,61 | 1174,58 | 1281,19 | 1243,46 |
| IL-6, | | 354,15 | 356,34 | 357,40 | 332,03 |
| n=8 | ЛПС 0,1 мкг/мл | 1435,55 | 1442,89 | 1346,26 | 1414,19 |
| | | 379,02а | 340,04 | 342,73 | 367,26 |
| | Без индуктора | 1198,38 | 1230,63 | 1294,63 | 1148,68 |
| IL-8, | | 153,25 | 85,79 | 219,75 | 200,81 |
| n=4 | ЛПС 0,1 мкг/мл | 1307,22 | 1422,48 | 1159,64 | 1228,32 |
| | | 213,04 | 90,89 | 121,80 | 154,70 |
| | Без индуктора | 280,38 | 364,25 | 352,83 | 359,05 |
| IL-1ra, | | 90,82 | 128,06 | 133,52 | 146,50 |
| n=4 | ЛПС 0,1 мкг/мл | 302,13 | 343,40 | 377,35 | 348,80 |
| | | 88,60 | 115,95 | 148,54 | 141,58 |
При оценке влияния опиатов на уровень TNF- выявлено угнетение продукции TNF- по сравнению с контролем при совместном внесение в культуры b-эндорфина и антагонистов опиатных рецепторов на 24 и 48 ч культивирования (см. рис. 18). На продукцию IL-6 на 24 ч культивирования b-эндорфин не влияет. На 48 ч культивирования продукцию IL-6 стимулирует налтриндол как при изолированном внесении, так и в комбинации с b-эндорфином. Учитывая, что во фракции лейкоцитов помимо моноцитов, основных продуцентов провоспалительных цитокинов, присутствуют гранулоциты, преимущественно нейтрофилы (Abraham et al., 2003; Fujiharaa et al., 2003; Xing, Remick, 2004), так же способные продуцировать IL-1 и IL-8, мы
24 ч 48 ч
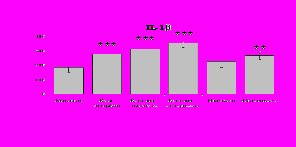
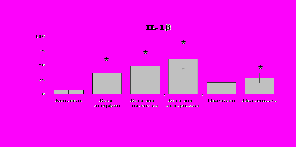
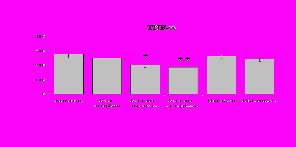
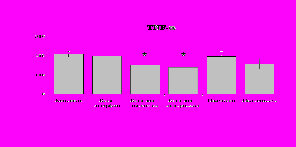
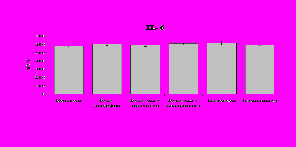
Рис. 18. Влияние -эндорфина 10-7М в условиях блокады опиатных рецепторов на продукцию IL-1, TNF-, IL-6 в нефракционированной клеточной суспензии в присутствии ЛПС.
* - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю по парному t-критерию Стъюдента..
исследовали влияние -эндорфина на продукцию IL-1 и IL-8 во фракциях мононуклеаров и нейтрофилов.
Результаты исследований влияния b-эндорфина на продукцию IL-1 и IL-8 во фракциях мононуклеаров и нейтрофилов представлены на рис. 20. b-эндорфин 10-7М стимулирует спонтанную и ЛПС-индуцированную продукцию IL-1b мононуклеарами. Аналогичная картина наблюдается во фракции нейтрофилов. На фоне LPS наблюдается статистически значимое усиление продукции IL-1b. b-эндорфин 10-7М стимулирует как спонтанную, так и LPS-индуцированную продукцию IL-1b нейтрофилами. ЛПС-индуцированная продукция IL-8 мононуклеарами и нейтрофилами под воздействием b-эндорфина снижается. В культурах без ЛПС пептид уровень IL-8 не изменяет.
При анализе влияния b-эндорфина на продукцию исследуемых цитокинов мононуклеарами в присутствии ФГА были получены следующие результаты (рис. 21). Во фракции мононуклеаров ФГА, как и ЛПС, усиливает выработку IL-1b и не влияет на уровень IL-8. b-эндорфин стимулирует как спонтанную, так и ФГА-индуцированную продукцию IL-1b мононуклеарами. Уровень IL-8 под воздействием пептида в нестимулированных ФГА пробах не изменяется, а в стимулированных - снижается. Как следует из полученных данных, продукцию цитокинов семейства IL-1 нефракционированными лейкоцитами, нейтрофилами и мононуклеарами (но не очищенными моноцитами) -эндорфин активирует, а продукцию IL-8 угнетает, таким образом, оказывая двоякое действие, с одной стороны, активируя процессы пролиферации и дифференцировки, а, с другой, угнетая хемотаксис. При этом в отличие от IL-1, депрессия IL-8
24 ч 48 ч
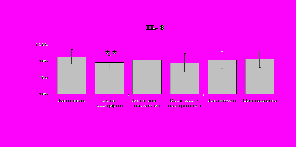
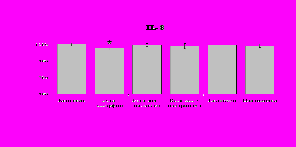
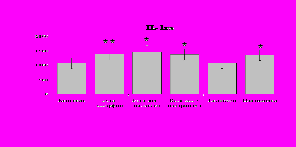

Рис. 19. Влияние -эндорфина 10-7М в условиях блокады опиатных рецепторов на продукцию IL-8 и IL-1ra в нефракционированной клеточной суспензии в присутствии ЛПС.
* - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю по парному t-критерию Стъюдента..
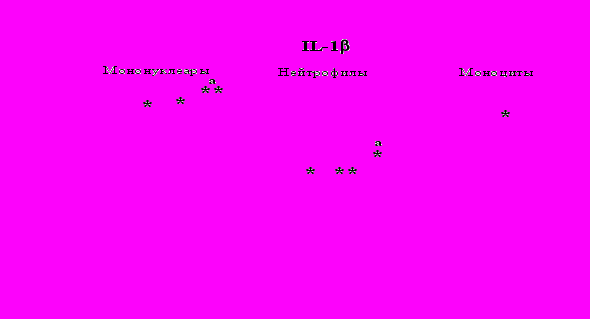
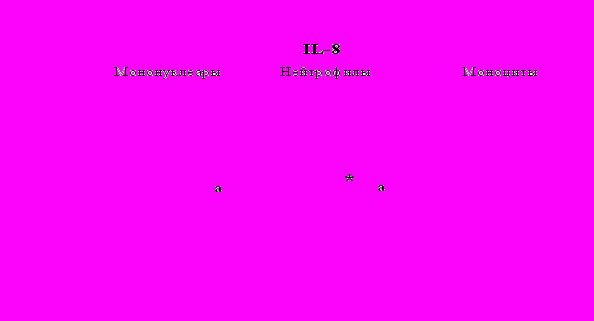
Рис. 20. Влияние -эндорфина 10-7М на продукцию IL-1 и IL-8 во фракциях мононуклеаров, нейтрофилов и моноцитов, стимулированных ЛПС.
К – контроль, 1 - -эндорфин, 2 – ЛПС, 3 – ЛПС+-эндорфин. * - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю, а - р < 0,05 к ЛПС по парному t-критерию Стъюдента..
проявляется только в ЛПС и ФГА-индуцированных культурах и нивелируется опиатными антагонистами, что указывает на вовлечённость различных рецепторных механизмов в регуляцию синтеза IL-1 и IL-8 -

Рис. 21. Влияние -эндорфина 10-7М на продукцию IL-1 и IL-8 мононуклеарами, стимулированными ФГА 2,5 мкг/мл в течении 24 ч культивирования.
К – контроль, 1 - -эндорфин, 2 – ФГА, 3 – ФГА+-эндорфин. * - р < 0,05; ** - р < 0,01; *** - р < 0,001 к контролю, а - р < 0,05 к ФГА по парному t-критерию Стъюдента.
эндорфином. Ранее (Nandhra, 2000) был зарегистрирован угнетающий блокируемый налоксоном, эффект b-эндорфина на продукцию IL-8 клетками хориодецидуальной оболочки. Н.Л. Елизарова с соавт. (2001) установили, что аминокислотная последовательность β-эндорфина в молекуле препарата тимуса – тактивина обусловливает активацию функциональной активности лимфоцитов, усиление фагоцитоза и снижение способности моноцитов к адгезии, последняя функция отменялась налоксоном, в то время как усиление фагоцитоза налоксоном не отменялось. По нашим данным, депрессия продукции IL-8 отменяется селективным -антагонистом налтриндолом, что свидетельствует о возможности реализации эффектов b-эндорфина через -рецептор. В то же время, как показано в работе B.M. Sharp (2006), в зависимости от экспериментальной модели и объекта исследования -рецептор может опосредовать проведение как стимулирующих, так и угнетающих сигналов.

Рис. 22. Схема возможного механизма влияния -эндорфина на процессы иммуногенеза.
Заключение
Результаты проведённых исследований представлены в виде схемы возможного механизма влияния -эндорфина на процессы иммуногенеза (рис. 22). Установлено, что -эндорфин является важным фактором в поддержании внутреннего гомеостаза. Приведённые в данные, свидетельствуют о том, что -эндорфин поддерживает гомеостаз иммунной системы через балансирование Th1 и Th2 ответа, смещая поляризацию Т-хелперов в сторону Th2-клеток, параллельно усиливая естественную резистентность. Следовательно, -эндорфин является кофактором, опосредующим переключение дифференцировки Т-хелперов с Th1 на Th2 тип и играющим решающую роль при целом ряде иммунопатологических состояний, а иммуномодулирующие функции опиоидных пептидов могут представлять значительный терапевтический интерес.
Оценивая роль эндогенных нейропептидов в стрессиндуцированных изменениях иммунной системы необходимо отметить, что полученные данные прямо указывают на непосредственное участие эндогенных опиатов в активации иммунной системы. Направленность ряда физиологических эффектов эндогенных опиоидных пептидов при стрессе противоположна эффектам глюкокортикоидов и катехоламинов. Ранее глюкокортикоиды и катехоламины рассматривались исключительно как стрессреализующие факторы, в то же время их основная биологическая роль может заключаться в ограничении опасной для организма стрессиндуцированной активации иммунных процессов.
ВЫВОДЫ
1. Подтверждена интегральная роль эндогенной опиоидной системы в нейроэндокринной регуляции иммуногенеза в норме, введении глюкокортикоидов и катехоламинов и в условиях стрессорного воздействия. Основные компоненты эндогенной опиоидной системы принимают непосредственное участие в активации иммунной системы при стрессе.
2. Установлено, что в условиях блокады , -опиатных рецепторов в период индукции иммунного ответа повышается степень выраженности иммуносупрессивного действия гормонов стресса (глюкокортикоидов и катехоламинов) на антителогенез и регистрируется снижение степени выраженности угнетающего действия глюкокортикоидов на количество ядросодержащих клеток в регионарном лимфатическом узле.
3. Впервые показано, что -эндорфин в высоких (100 мкг/мл) дозах угнетает, в низких (0,0005 мкг/кг) дозах стимулирует гуморальное звено иммунитета и при этом не влияет на степень выраженности реакции гиперчувствительности замедленного типа. В клеточных культурах -эндорфин стимулирует пролиферацию лимфоцитов и продукцию IL-4, не влияет на синтез IL-2, IFN- и при этом усиливает степень поляризации Т-хелперов в направлении Th2-клеток. Доминирующая роль в реализации стимулирующих эффектов -эндорфина на пролиферацию и продукцию IL-4 принадлежит -рецепторам.
4. Впервые обнаружена зависимость стимулирующего эффекта опиоидных пептидов на пролиферацию лимфоцитов и продукцию IL-4 от присутствия моноцитов в клеточной культуре.
5. Показано, что -эндорфин стимулирует фагоцитарную активность клеток естественного иммунитета и оказывает модулирующее действие на цитокинпродуцирующую функцию моноцитов и нейтрофилов. В условиях блокады , -опиатных рецепторов отмены стимулирующего влияния -эндорфина на продукцию IL-1 не наблюдается, при этом угнетение продукции IL-8 опиатными антагонистами отменяется.
Список работ, опубликованных по теме диссертации
Экспериментальные статьи
1. Шилов Ю.И., Гейн С.В. Адренергическая регуляция пролиферативного ответа лимфоцитов в культурах с Т-клеточными митогенами // Бюллетень экспериментальной биологии и медицины. - 1999. - Т. 128, № 8. - С. 207-209.
2. Шилов Ю.И., Гейн С.В, Черешнев В.А. Влияние блокады b-адренергических рецепторов при стрессе на антителообразование, гиперчувствительность замедленного типа, функции фагоцитирующих клеток при локальной форме иммунного ответа // Russian Journal of Immunology. - 2001. - V. 6, № 3. - P. 301-308.
3. Гейн С.В., Симоненко Т.А., Черешнев В.А. Влияние -эндорфина и селективного агониста -опиатных рецепторов DAGO на пролиферативную активность лимфоцитов // Доклады академии наук. - 2003. - Т. 391, № 1. - С. - 127-129.
4. Гейн С.В., Симоненко Т.А., Тендрякова С.П. Влияние агонистов опиатных рецепторов на процесс антителообразования и гиперчувствительность замедленного типа при локальном иммунном ответе // Вестн. Пермск. университета. Серия Биолгия. - 2004. - Вып. 2. - С. 166-168.
5. Гейн С.В., Тендрякова Т.А. Иммуномодулирующие эффекты гидрокортизона и адреналина в условиях блокады опиатных рецепторов // Докл. Академии наук. - 2004. - Т. 399, № 3. - С. 412-414.
6. Gein S.V., Simonenko T. A., Tendryakova S. P. The effects of rotation stress on measures of immunity. The role of opiate receptors // Neuroscience and Behavioral Physiology. - 2004. - V. 34, № 9. - P. 935-938.
7. Гейн С.В., Баева Т.А. Роль опиоидных пептидов в регуляции пролиферации лимфоцитов и изменении Тh1/Тh2 цитокинового профиля // Проблемы эндокринологии. - 2005. - № 5. - С. 49-51.
8. Гейн С.В., Шаравьёва И.Л, Баева Т.А., Тендрякова Т.А. Роль блокады различных типов опиатных рецепторов в регуляции иммуномодулирующих эффектов ротационного стресса // Вестник Уральской медицинской академической науки. - 2005. - № 3. - С. 70-73.
9. Гейн С.В., Баева Т.А., Гейн О.Н., Черешнев В.А. Роль моноцитов в реализации эффектов -эндорфина и селективных агонистов - и -опиатных рецепторов на пролиферативную активность лимфоцитов периферической крови // Физиология человека. - 2006. - Т. 32, № 3. – С. 111-116.
10. Гейн С.В., Шаравьёва И.Л, Тендрякова Т.А. Роль -опиатных рецепторов в регуляции гуморального и клеточноопосредованного иммунного ответа при ротационном стрессе // Доклады Академии наук. - 2006. - Т. 407, № 1. - С. 127-129.
11. Гейн С.В., Баева Т.А., Кичанова О.А. Влияние -эндорфина на антителогенез и продукцию ИЛ-4 в условиях блокады опиатных рецепторов // Бюллетень экспериментальной биологии и медицины. - 2006. - Т. 142, № 8. – С. 192-195.
12. С.В. Гейн, Е.Г. Чижова, С.П. Тендрякова Влияние гидрокортизона и адреналина в условиях блокады - и -опиатных рецепторов на развитие локального иммунного ответа у мышей // Российский физиологический журнал им. Сеченова. - 2006. - Т. 92, № 7. - С. 827-834.
13. Горшкова К.Г., Гейн С.В., Тендрякова С.П. Роль бета-эндорфина в регуляции продукции IL-1 и IL-8 моноцитами и нейтрофилами периферической крови // Вестник Уральской медицинской академической науки. - 2006. - № 3-1(14). - С. 46-47.
14. Гейн С.В., Баева Т.А., Чижова Е.Г., Кичанова О.А., Шаравьева И.Л. Эндогенная опиоидная система и гормоны стресса в сочетанной регуляции иммунных реакции при остром стрессе // Вестник Уральской медицинской академической науки. - 2006. - № 3-1(14). - С. 38-40.
15. Гейн С.В., Горшкова К.Г., Тендрякова С.П.. Роль b-эндорфина в регуляции продукции провоспалительных цитокинов моноцитами периферической крови in vitro // Бюллетень экспериментальной биологии и медицины. - 2007. - Т. 143, № 2. - С. 175-178.
Труды конференций
16. Shilov Ju.I., Gein S.V., Chereshnev V.A.. Adrenergic and cAMP-dependent regulation of mitogen-induced lymphocyte proliferative responses // Annual Meeting of AAAAI. J. Allergy and Clin. Immunol. - 1998. - V. 101, № 1. - S. 38-38.
17. Gein S.V., Shilov Ju.I. Influence of stress on expression of immune reactions // Межд. конф. "Проблемы загрязнения окружающей среды-98". Москва, 1998. - С. 238-238.
18. Шилов Ю.И., Гейн С.В. Влияние блокады бета-адренергических рецепторов и иммобилизационного стресса на реакции клеточноопосредованного и гуморального иммунитета // Научная сессия Пермской государственной медицинской академии. Пермь, 1998. - С. 68-68.
19. Гейн С.В., Шилов Ю.И. Адренергическая и цАМФ-зависимая регуляция пролиферативного ответа лимфоцитов в культурах с митогенами // Научная сессия Пермской государственной медицинской академии. Пермь, 1998. - С. 69-69.
20. Гейн С.В., Шилов Ю.И. Адренергическая и цАМФ-зависимая модуляция пролиферативного ответа лимфоцитов в культурах с митогенами // V Российский национальный конгресс "Человек и лекарство". Москва, 1998. - C. 330-330.
21. Шилов Ю.И., Гейн С.В. Модуляция пропранололом стресс-индуцированных изменений иммунных реакций // Там же. - C. 340-340.
22. Гейн С.В., Шилов Ю.И., Черешнев В.А. Влияние адренергических соединений на пролиферативный ответ лимфоцитов // II-й Национальный конгресс Российской Ассоциации аллергологов и клинических иммунологов «Современные проблемы аллергологии, клинической иммунологии и иммунофармакологии». Москва, 1998. - С. 345-345.
23. Шилов Ю.И., Гейн С.В. Влияние иммобилизационного стресса и блокады b-адренергических рецепторов на антителообразование и реакции гиперчувствительности замедленного типа в регионарных лимфатических узлах и селезенке при подкожной иммунизации // Иммунология. - 1998. - № 6. - С. 25-26.
24. Шилов Ю.И., Гейн С.В. Влияние адреналина и селективных антагонистов a1- и a2-адренорецепторов на пролиферативный ответ лимфоцитов периферической крови // Матер. III Всерос. научного Форума «Дни иммунологии в Санкт-Петербурге». Медицинская иммунология. - 1999. - Т. 1, № 3-4. - С. 28-29.
25. Гейн С.В., Шилов Ю.И. Роль адренорецепторов в регуляции пролиферативного ответа лимфоцитов в культурах с Т-клеточными митогенами // Всероссийской научно-практической конференция "Актуальные вопросы эндокринологии". Пермь, 1999. - С. 101-102.
26. Шилов Ю.И., Гейн С.В., Орлова Е.Г., Черешнев В.А. Влияние блокады b-адренергических рецепторов на иммунный ответ и функциональную активность фагоцитирующих клеток в условиях стресса // Аллергология и иммунология. - 2000. - Т. 1, № 2. - С. 116-117.
27. Шилов Ю.И., Гейн С.В. Иммуномодулирующие эффекты адренергических соединений в системах in vivo и in vitro // Труды российской научной конференции «Факторы клеточного и гуморального иммунитета при различных физиологических и патологических состояниях». Челябинск, 2000. - С. 134-135.
28. Кеворков Н.Н., Шилов Ю.И., Бахметьев Б.А., Ширшёв С.В., Куклина Е.М., Гейн С.В., Сидоров Д.В., Заморина С.А., Орлова Е.Г., Лихачева Н.С., Груздева Е.А., Харитонова А.В. Нейроэндокринная регуляция некоторых функций фагоцитирующих и иммунокомпетентных клеток // Тез. докл. научной конференции «Актуальные проблемы фундаментальных исследований в области биологии и медицины». Санкт-Петербург, 2000. - С. 67-68.
39. Гейн С.В. Роль b-адренергических механизмов в регуляции реакций клеточноопосредованного и гуморального иммунного ответа при стрессе // II конференция молодых ученых России с международным участием “Фундаментальные науки и прогресс клинической медицины”. Москва. - Т. 1. - С. 191-191.
30. Гейн С.В., Симоненко Т.А., Тендрякова С.П., Шилов Ю.И. Влияние блокады -,-опиатных рецепторов при стрессе на показатели клеточноопосредованного, гуморального иммунитета в условиях развития локальной формы иммуного ответа // I конференция иммунологов Урала. Екатеринбург, Иммунология урала. – 2001. - №1. – С. 2-3.
31. Гейн С.В., Симоненко Т.А., Тендрякова С.П. Участие -, , -опиатных рецепторов в развитии реакции гиперчувствительности замедленного типа и изменении числа антителообразующих клеток в модели принудительного головокружения // Труды международной научной конференции «Перспективы развития естественных наук в высшей школе». Пермь, 2001. - Т. 3. - С.120-124.
32. Гейн С.В., Симоненко Т.А., Тендрякова С.П. Роль -,-,-опиатных рецепторов в регуляции функций иммунокомпетентных и фагоцитирующих клеток при стрессе // II конференция иммунологов Урала. Пермь, Иммунология Урала. – 2002. – №1. – С. 4.
33. Гейн С.В., Симоненко Т.А. Участие -эндорфина и селективных лигандов , -опиатных рецепторов в регуляции пролифератив-ной активности лимфоцитов in vitro // III конференция иммунологов Урала. Челябинск, Иммунология урала. – 2003. - № 1. - С. 5.
34. Гейн С.В., Симоненко Т.А. Роль -эндорфина и опиодных пептидов в регуляции пролиферации лимфоцитов и изменении Тh1/Тh2 цитокинового профиля // Матер. VIII Всерос. научного Форума с международным участием «Дни иммунологии в Санкт-Петербурге». Медицинская иммунология. - 2004. - V. 6, № 3-5. - P. 226-227.
35. Гейн С.В., Симоненко Т.А. Опиоидная регуляция пролиферативного ответа лимфоцитов in vitro // Rus. J. Immunol. - 2004. - V. 9, № 1. - P. 38-38.
36. Гейн С.В., О.Н. Гейн, Т.В. Гаврилова Механизмы стрессорных нарушений функций иммунной системы и их коррекция // XIX Съезд физиологического общества им. И.П. Павлова 19-24 сентября 2004. Тез. докл. – Российский физиологический журнал им. Сеченова. - 2004. - Т. 90, №8. - С. 106-107.
37. Гейн С.В., Чижова Е.Г. Влияние гидрокортизона на фоне блокады опиатных рецепторов на развитие локального иммунного ответа // IX международная Пущинская школа-конференция молодых ученых «Биология-наука XXI века». Пущино, 2005. - С. 174.
38. Гейн С.В., Баева Т.А. Роль опиодных пептидов в регуляции пролиферации лимфоцитов и изменении Th1/Th2 цитокинового профиля // Там же. - С. 69.
39. Баева Т.А., Гейн С.В., Шаравьева И.Л., Тендрякова С.П. Роль блокады различных типов опиатных рецепторов в регуляции иммуномодулирующих эффектов ротационного стресса // Международная конференция "Проблемы загрязнения окружающей среды-2005". Пермь, 2005. - С. 56.
40. Гейн С.В., Баева Т.А., Черешнев В.А. Лиганды опиатных рецепторов в регуляции пролиферативной активности лимфоцитов in vitro // Международная конференция "Проблемы загрязнения окружающей среды-2005". Пермь, 2005. - С. 94.
41. Шилов Ю.И., Шилов С.Ю., Ланин Д.В., Гейн С.В., Орлова Е.Г., Черешнев В.А. Адренергические механизмы регуляции иммунного ответа и функций неспецифических эффекторных клеток при стрессе // Матер. IX Всерос. научного Форума с международным участием «Дни иммунологии в Санкт-Петербурге». Медицинская иммунология. - 2005. - Т. 7, № 2-3. - С. 127-127.
42. Гейн С.В., Баева Т.А., Черешнев В.А. Роль моноцитов в опиатэргической регуляции процессов пролиферации лимфоцитов и цитокинового синтеза // Там же. - С. 110-110.
43. Гейн С.В., Баева Т.А., Черешнев В.А. Роль -эндорфина в регуляции процессов пролиферации и продукции IL-4 лимфоцитами периферической крови in vitro // IV конференция иммунологов Урала. Уфа, Иммунология Урала. - 2005. - № 1 (4). - С. 4-5.
44. Гейн С.В., Чижова Е.Г., Тендрякова С.П. Влияние гидрокортизона на фоне блокады -, -опиатных рецепторов на иммунный ответ // Там же. - С. 5-6.
45. Гейн С.В., Горшкова К.Г. Влияние -эндорфина на продукцию IL-1, TNF-, IL-6 моноцитами периферической крови // V конференция иммунологов Урала. Оренбург, Иммунология урала. - 2005. - № 1. - с. 6-7.
46. Gein S.V., Gorshkova K.G., Baeva T.A. -endorphin-induced modulation of cytokine production by peripheral blood leukocytes // International symposium «Interaction of the nervous and immune systems in health and disease» Saint-Petersburg, 2007. - Р. 26-27.
Список основных сокращений
АКТГ - адренокортикотропный гормон
АОК - антителообразующая клетка
ГЗТ - гиперчувствительность замедленного типа
ЛПС - липополисахарид
ПОМК - проопиомеланокортин
РБТЛ - реакция бласттрансформации лимфоцитов
СНС - симпатическая нервная система
ФГА - фитогемагглютинин
ЦНС - центральная нервная система
ЯСК - ядросодержащая клетка
CD - маркеры дифференцировки лимфоцитов
DADLE - δ-агонист (d–Ala2,d-Leu5 - энкефалин
DAGO - -агонист (d-Ala2,N-Me-Phe4,Gly5-ol-энкефалин
IFN - интерферон
IL - интерлейкин
MPF - меланотропин потенцирующий фактор
TNF - фактор некроза опухоли