
Молекулярно-генетический анализ генофондов редких и исчезающих видов растений пермского края
| Вид материала | Автореферат диссертации |
- Красная книга, 61.32kb.
- Венции должны содействовать восстановлению исчезающих видов, в частности, путем разработки, 98.37kb.
- Молекулярно-генетический анализ тапеторетинальной абиотрофии в республике башкортостан, 292.82kb.
- «Путешествие по регионам Большого Алтая, 177.44kb.
- Международного Союза Охраны Природы Питеру Скотту. По его инициативе в мсоп в 1949, 1423.12kb.
- Темы Вашего учебного проекта, 61.28kb.
- Правительство Республики Казахстан постановляет : Утвердить прилагаемую сохранения, 754.5kb.
- Правительство Республики Казахстан постановляет: Утвердить прилагаемую программу сохранения, 712.05kb.
- Секция: Стратегия сохранения редких и исчезающих видов наземной фауны, 135.39kb.
- Об охране окружающей среды, 2522.13kb.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
3. Биологические особенности редких реликтовых видов растений и характеристика избранных для изучения популяций
3.1. Adonis vernalis L.
В Пермском крае отмечено 19 местонахождений A. vernalis. За последние 15 лет общая численность этого вида сократилась на 20-25% (Боронникова, 2008). Показано, что основной вклад в семенное размножение A. vernalis вносят средневозрастные генеративные особи (
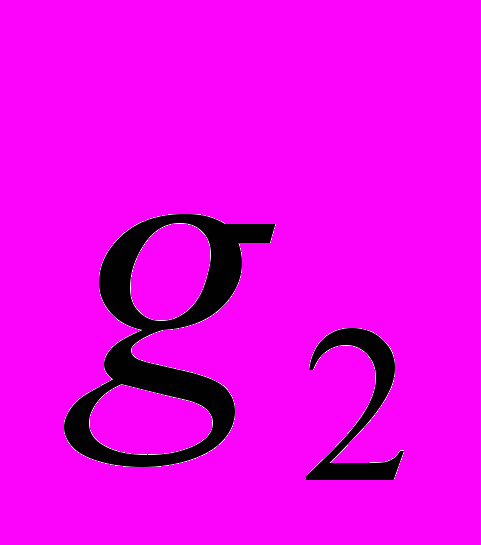 ), составляющие эффективную численность популяций. Установлено, что к популяциям с низкой общей численностью относятся Av3, Av6, Av7, Av8, Av10 (от 39 в Av7 до 184 особей в Av6), а к популяциям с высокой общей численностью – Av1,Av2, Av4, Av5, Av9 (от 248 в Av2 до 562 особей в Av1). Нами установлено, что в изученных популяциях A. vernalis с низкой общей численностью эффективная численность варьировала от 8 (Av7) до 64 особей (Av6), а в популяциях с высокой общей численностью эффективная численность – от 99 (Av4) до 194 (Av9) особей. Популяции Av3 , Av6, Av7 и Av10 находятся под антропогенным воздействием сильной степени, популяции Av, Av2, Av5 и Av8 – средней, а Av4 и Av9 – слабой степени.
), составляющие эффективную численность популяций. Установлено, что к популяциям с низкой общей численностью относятся Av3, Av6, Av7, Av8, Av10 (от 39 в Av7 до 184 особей в Av6), а к популяциям с высокой общей численностью – Av1,Av2, Av4, Av5, Av9 (от 248 в Av2 до 562 особей в Av1). Нами установлено, что в изученных популяциях A. vernalis с низкой общей численностью эффективная численность варьировала от 8 (Av7) до 64 особей (Av6), а в популяциях с высокой общей численностью эффективная численность – от 99 (Av4) до 194 (Av9) особей. Популяции Av3 , Av6, Av7 и Av10 находятся под антропогенным воздействием сильной степени, популяции Av, Av2, Av5 и Av8 – средней, а Av4 и Av9 – слабой степени.3.2. Adonis sibirica Patrin ex Ledeb.
В Пермском крае выявлено 16 местонахождений A. sibirica. Размножение этого вида в естественных условиях происходит только семенным путем. Установлено, что как и у A. vernalis, так и у A. sibirica принимают участие в семенном размножении преимущественно средневозрастные генеративные особи (
). Общая численность в As1 составила 22 особи, в As2 – 246, в As3 – 257 особей. В As1 к настоящему времени эффективная численность равна 3 особям, в As2 и As3 – 89 и 77 особям соответственно. C 1994 по 2008 гг. из-за строительства горнолыжной трассы общая численность As1 сократилась на 75.00 %, а эффективная численность – на 85.71% (рис. 1). Исследованные популяции находятся под антропогенным воздействием As1 сильной, As2 средней и As3 слабой степени.1.3. Paeonia anomala L.
В северной части Пермского края P. anomala представлен популяциями с большой стабильной численностью. В центральной части края из-за антропогенного воздействия численность популяций сокращается. Общая численность Pa2 в 1988 составляла 562 особи (Боронникова, 2002), а в 2008 году – 377 особей, то есть за 20 лет сократилась на 41.04%. Эффективная численность популяции за этот же период сократилась с 402 до 224 особей, то есть на 44.28% (рис. 1). Одной из причин сокращения показателей численности Pa2 является усиление антропогенного воздействия в связи со строительством новой автомобильной трассы в 1 км от Pa2. В Pa1 общая численность составила 957, а эффективная – 190 особей. В Pa3 общая численность составила 441 особь, а эффективная численность – 146 особей. По нашим данным в природе P. anomala размножается только семенным путем (Боронникова, 2002). Нами установлено, что в семенном размножении у P. anomala принимают участие особи g2 , редко g3. Изученные популяции находятся под антропогенным воздействием средней степени, кроме Pa2, которая испытывает антропогенное воздействие сильной степени.
| As1 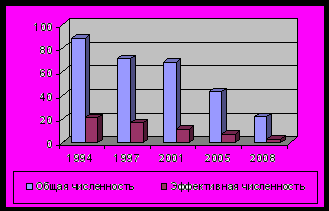 | Pa2 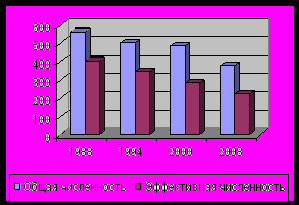 |
Рис. 1. Динамика общей и эффективной численности первой популяции (As1) A. sibirica и второй популяции (Pa2 ) P. anomala
1.4. Adenophora lilifolia (L.) A.DC.
Общая численность вида в крае сократилась за последние 10 лет в среднем на 15-20% (Боронникова, 2008). В изученных популяциях Ad. lilifolia этот показатель варьировал от 59 особей в Ad.lil.4 до 293 особей в Ad.lil.1, а эффективная численность – от 36 в в Ad.lil. 4 до 239 особей в Ad.lil.1. По нашим данным размножение вида в природе осуществляется и семенным и вегетативным, то есть комбинированным способом (Боронникова, 1999). Наибольший вклад в семенную продуктивность вносят особи g2 и g3, которые и определяют эффективную численность популяций. Сильную степень антропогенного воздействия испытывает популяция Ad.lil.4, среднюю – Ad.lil.1, а слабую – Ad.lil.2 и Ad.lil.3.
1.5. Digitalis grandiflora Mill.
D. grandiflora на территории Пермского края встречается, в основном, в островной Кунгурской лесостепи, где и отмечено 12 местонахождений этого вида. Общая численность изученных популяций D. grandiflora варьировала от 56 (Dg3) до 487 особей (Dg1), а эффективная – от 40 до 304 особей соответственно. Для вида характерен комбинированный (семенной и вегетативный) способ размножения. При анализе показателей семенной продуктивности особей D. grandiflora разных возрастных групп генеративного периода было отмечено, что у средневозрастных особей показатели выше, чем у особей остальных групп (Боронникова, 2009б). Эффективная численность у данного вида представлена особями g2 и g3. Изученные популяции D. grandiflora находятся под антропогенным воздействием сильной (Dg3), средней (Dg1, Dg2) и слабой степени (Dg4, Dg5).
Таким образом, избранные для изучения 5 редких реликтовых видов представлены в Пермском крае, кроме P. anomala, небольшим числом сравнительно малых по численности популяций. Три из изученных видов (A. vernalis, A. sibirica, P. anomala) размножаются только семенным путем. Для Ad. lilifolia и D. grandiflora характерен комбинированный (и семенной и вегетативный) способ размножения. Эффективная численность популяций выявлена благодаря изучению участия в семенном размножении особей разных возрастных групп генеративного периода. Самые низкие значения данного показателя характерны для видов рода Adonis (3 особи у A. sibirica и 8 особей у A. vernalis), а самые высокие – для P. anomala (от 146 до 224 особей).
4. Анализ генофондов редких видов растений на основании оценок полиморфизма фрагментов ДНК, фланкированных инвертированными микросателлитными повторами (ISSR-PCR маркеры)
4.1. Анализ генетического разнообразия популяций двух видов рода Adonis
В изученных популяциях A. vernalis выявлено 109 амплифицированных ISSR-фрагментов ДНК, из которых 100 были полиморфными. Число амплифицированных фрагментов ДНК в общей выборке растений варьировало в зависимости от праймера от 19 (М9) до 29 (М12). В среднем при ISSR-анализе у A. vernalis один праймер инициировал синтез 22 фрагментов ДНК. Число полиморфных ISSR-фрагментов A. vernalis варьировало от 16 до 29, а их размеры – от 210 до 1900 пн (рис. 2).
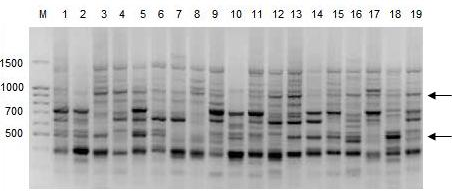
Рис. 2. ISSR-спектр популяции A.vernalis (Аv1) c праймером М1: цифрами обозначены номера проб, М - молекулярный маркер, стрелками указаны некоторые полиморфные фрагменты ДНК
Доля полиморфных локусов в общей выборке в среднем (при
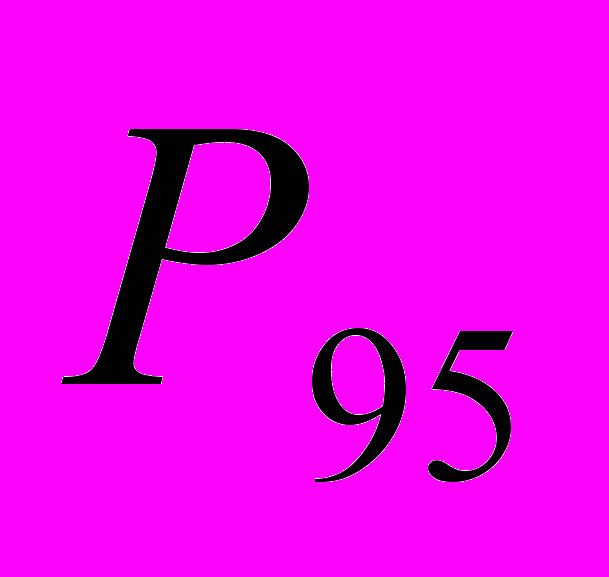 ) составила 91.74%. В популяциях данный показатель варьировал от 34.86% в Av2 до 62.39% в Av6. Ожидаемая гетерозиготность A. vernalis по локусам равна 0.313. Этот показатель самый высокий в Av4 (0.242), а низкий – в Av2 (0.147) (Боронникова и др., 2007; Боронникова и др., 2008). Абсолютное число аллелей на локус (в нашем случае на фрагмент ДНК) на общую выборку составило 1.973, а эффективное число аллелей на локус (
) составила 91.74%. В популяциях данный показатель варьировал от 34.86% в Av2 до 62.39% в Av6. Ожидаемая гетерозиготность A. vernalis по локусам равна 0.313. Этот показатель самый высокий в Av4 (0.242), а низкий – в Av2 (0.147) (Боронникова и др., 2007; Боронникова и др., 2008). Абсолютное число аллелей на локус (в нашем случае на фрагмент ДНК) на общую выборку составило 1.973, а эффективное число аллелей на локус (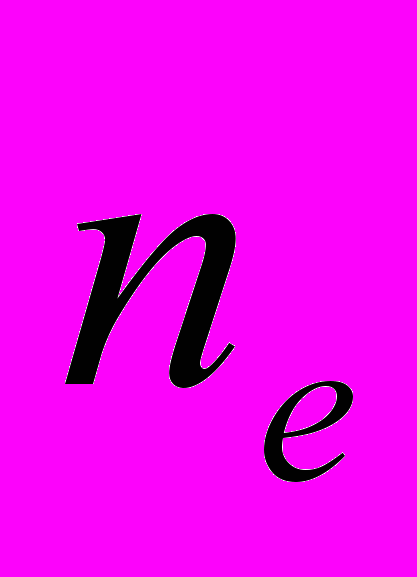 ) – 1.521. Число редких аллелей (фрагментов ДНК с частотой <=0,05) варьировало по популяциям от 0 (Av4) до 8 (Av6).
) – 1.521. Число редких аллелей (фрагментов ДНК с частотой <=0,05) варьировало по популяциям от 0 (Av4) до 8 (Av6). Генетическая структура изученных популяций A. vernalis: ожидаемая доля гетерозиготных генотипов в общей популяции выше (
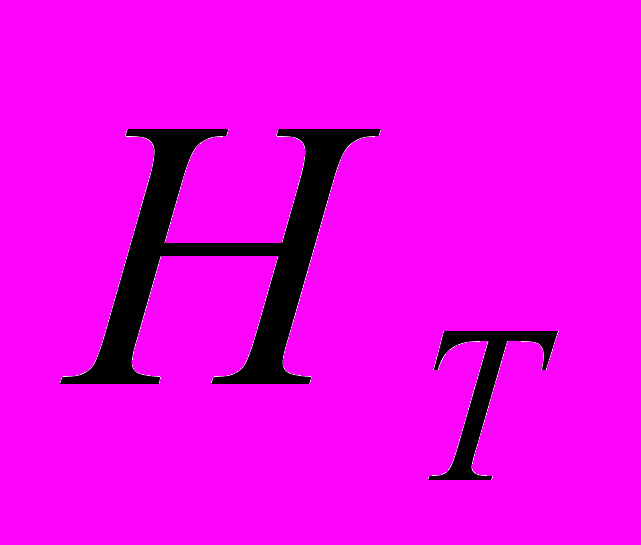 =0.313), чем в субпопуляциях (
=0.313), чем в субпопуляциях (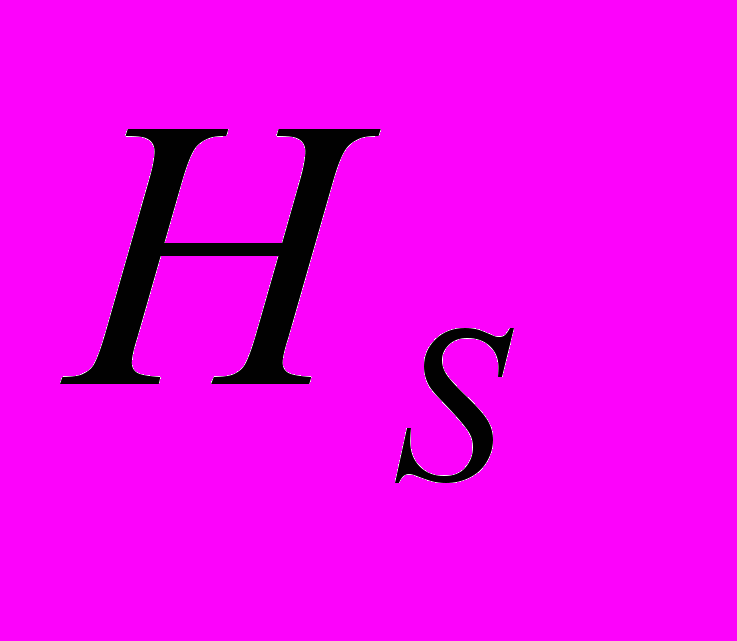 =0.184). Коэффициент подразделенности популяций (
=0.184). Коэффициент подразделенности популяций (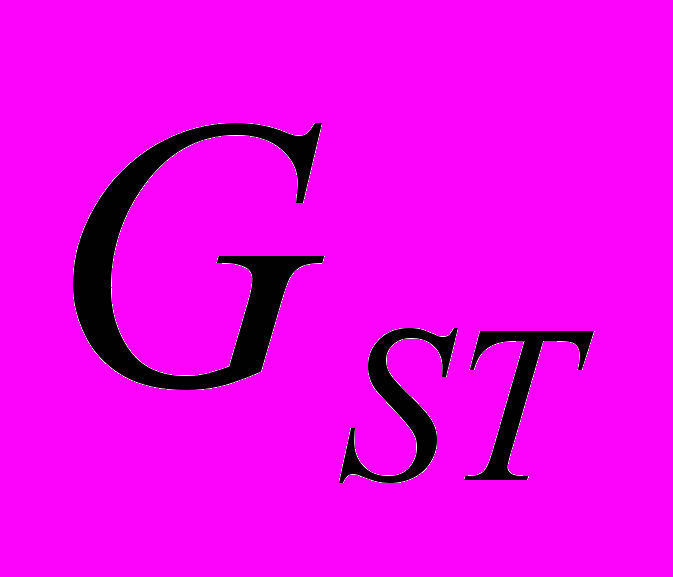 ) показывает, что на межпопуляционную компоненту генетического разнообразия A. vernalis приходится 41.22% разнообразия, то есть изученные популяции сильно дифференцированы (Боронникова и др., 2009г). Наименьшее генетическое расстояние между исследуемыми популяциями A. vernalis отмечено между Av2 и Av6 (0.109), а наиболее генетически удаленными являются популяции Av1 и Av4 (0.305). На дендрограмме (рис. 3) популяции A. vernalis сформировали 3 кластера по 2 выборки, соответствующие исследованным популяциям. Узлы ветвления имеют высокую поддержку (индекс бутстрепа >50%).
) показывает, что на межпопуляционную компоненту генетического разнообразия A. vernalis приходится 41.22% разнообразия, то есть изученные популяции сильно дифференцированы (Боронникова и др., 2009г). Наименьшее генетическое расстояние между исследуемыми популяциями A. vernalis отмечено между Av2 и Av6 (0.109), а наиболее генетически удаленными являются популяции Av1 и Av4 (0.305). На дендрограмме (рис. 3) популяции A. vernalis сформировали 3 кластера по 2 выборки, соответствующие исследованным популяциям. Узлы ветвления имеют высокую поддержку (индекс бутстрепа >50%).
Рис. 3. UPGMA дендрограмма генетического сходства популяций, построенная на основании полиморфизма ISSR-маркеров A.vernalis. Шкала сверху – генетические дистанции. На дендрограмме указаны значения бутстрепа (в %)
Среднее значение индексов разнообразия Шеннона (
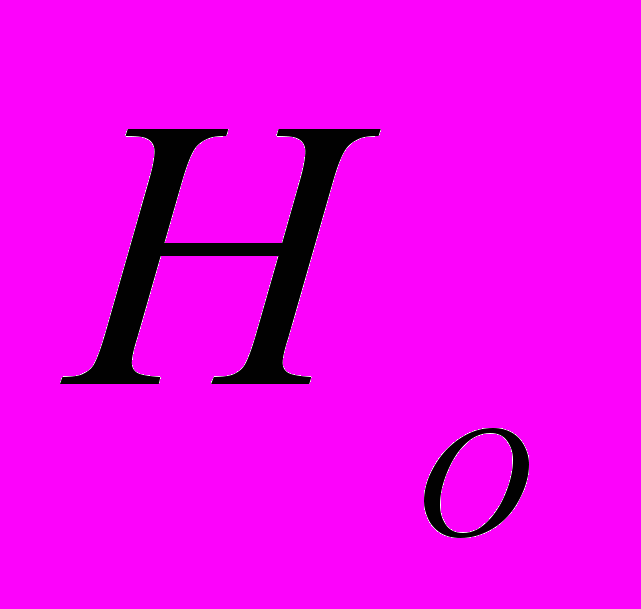 ) в изученных популяциях A. vernalis, рассчитанные по ISSR-праймерам, составило 0.278. Выше этот показатель в Av4. Индекс разнообразия Шеннона, рассчитанный на общую популяцию A. vernalis, равен 0.477. На долю внутрипопуляционного генетического разнообразия A. vernalis приходится 58.40%, а на долю межпопуляционного – 42.60%. Итак, оба подхода к определению генетического разнообразия, то есть определение показателя подразделенности популяций () и коэффициента разнообразия Шеннона () у A. vernalis дали близкие результаты.
) в изученных популяциях A. vernalis, рассчитанные по ISSR-праймерам, составило 0.278. Выше этот показатель в Av4. Индекс разнообразия Шеннона, рассчитанный на общую популяцию A. vernalis, равен 0.477. На долю внутрипопуляционного генетического разнообразия A. vernalis приходится 58.40%, а на долю межпопуляционного – 42.60%. Итак, оба подхода к определению генетического разнообразия, то есть определение показателя подразделенности популяций () и коэффициента разнообразия Шеннона () у A. vernalis дали близкие результаты.Таким образом, самые высокие показатели генетического разнообразия отмечены в популяциях Октябрского района (Av4, Av5, Av6), а самые низкие показатели – в самой северной популяции на Спасской горе (Av2). Высокое число редких аллелей в Av2 (
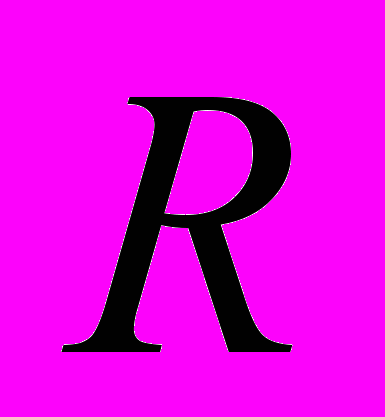 =7) указывает на необходимость сохранения данной популяции в качестве генетически уникальной.
=7) указывает на необходимость сохранения данной популяции в качестве генетически уникальной. Полиморфизм структурных элементов генома A. sibirica был выявлен с использованием тех же ISSR-праймеров, что и у A. vernalis. В трех изученных популяциях A. sibirica ISSR-праймеры инициировали синтез меньшего, чем у A. vernalis, числа ампликонов, равного 74, из которых 66 были полиморфными (
=89.19%). Размеры ампликонов в A. sibirica варьировали от 270 до 1620 пн. В среднем при ISSR-анализе у A. sibirica один праймер инициировал синтез 14.8 фрагментов ДНК, что значительно меньше, чем у A. vernalis (22 ампликона). Высокий процент полиморфизма ISSR-маркеров отмечен в As3 (=81.08%), а самый низкий – в As1 (=35.13%). Низкие значения ожидаемой гетерозиготности (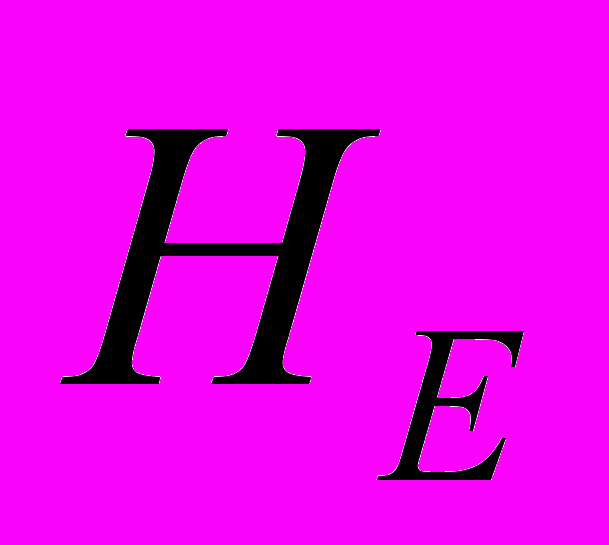 ) отмечены в As1 (0.142), а высокие – в As2 (0.243). В общей популяции A. sibirica ожидаемая гетерозиготность выше и составила 0.356. Абсолютное число аллелей на локус на общую выборку A. sibirica составило 1.914. Этот параметр выше в As2 и ниже в As1. Эффективное число аллелей на локус () на общую популяцию составило 1.608. Наибольшее значение данного показателя отмечено в As3 (1.409), а его резкое снижение – в As1 (1.257). Наибольшее число редких аллелей отмечено в As3 (=11). Редкие аллели отсутствуют в As1, что свидетельствует об обеднение генофонда этой популяции. Генетическая структура популяций A. sibirica такова: ожидаемая доля гетерозиготных генотипов выше в общей популяции (
) отмечены в As1 (0.142), а высокие – в As2 (0.243). В общей популяции A. sibirica ожидаемая гетерозиготность выше и составила 0.356. Абсолютное число аллелей на локус на общую выборку A. sibirica составило 1.914. Этот параметр выше в As2 и ниже в As1. Эффективное число аллелей на локус () на общую популяцию составило 1.608. Наибольшее значение данного показателя отмечено в As3 (1.409), а его резкое снижение – в As1 (1.257). Наибольшее число редких аллелей отмечено в As3 (=11). Редкие аллели отсутствуют в As1, что свидетельствует об обеднение генофонда этой популяции. Генетическая структура популяций A. sibirica такова: ожидаемая доля гетерозиготных генотипов выше в общей популяции (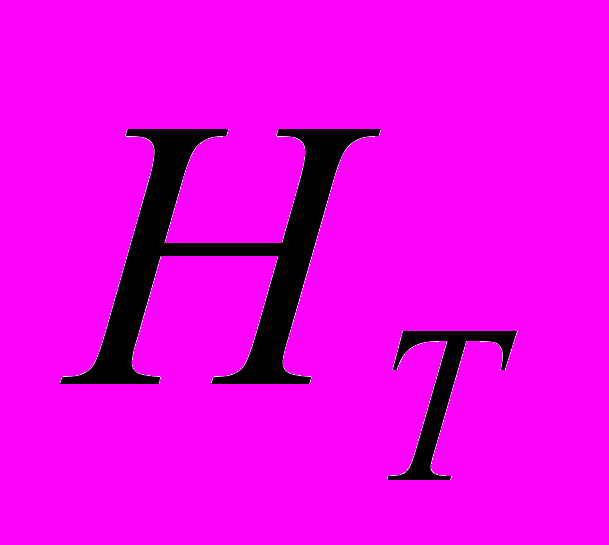 =0.334), чем в субпопуляциях (
=0.334), чем в субпопуляциях (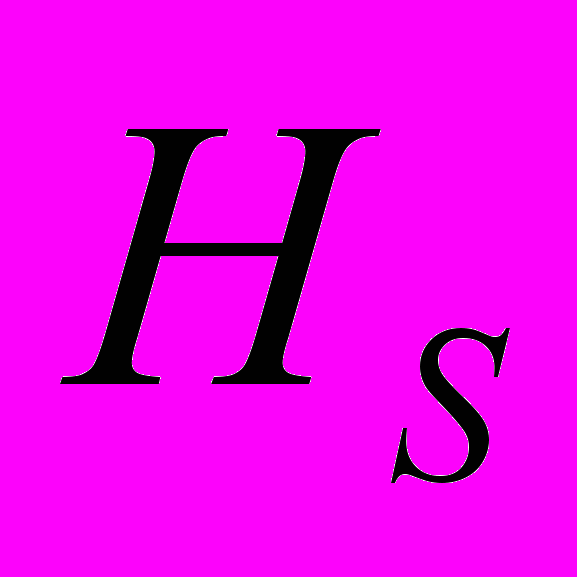 =0.214). Показатель генетической подразделенности популяций () A. sibirica равен 0.358. Генетически более гетерогенна As3 (=81.08%; =0.235; =1.409), а наименьшие основные показатели генетической изменчивости отмечены в As1 (=35.13%; =0.145; =1.257). Таким образом, на межпопуляционную изменчивость у A. sibirica приходится 35.77% генетического разнообразия. Наибольшее генетическое расстояние отмечено между As2 и As3 (0.290), а наименьшее между As1 и As2 (0.166). Кластерный анализ показал, что As1 генетически ближе к As2. Третья популяция (As3) находится на большем генетическом расстоянии от первых двух популяций и характеризуется наименьшей степенью родства с ними. Среднее значение индексов разнообразия Шеннона () в изученных популяциях A. sibirica, рассчитанные по ISSR-праймерам, составило 0.292. Выше этот показатель в As3. Индекс Шеннона, рассчитанный на общую популяцию A. sibirica, равен 0.504. На долю внутрипопуляционного генетического разнообразия A. sibirica приходится 59.55%, а на долю межпопуляционного – 40.45% разнообразия.
=0.214). Показатель генетической подразделенности популяций () A. sibirica равен 0.358. Генетически более гетерогенна As3 (=81.08%; =0.235; =1.409), а наименьшие основные показатели генетической изменчивости отмечены в As1 (=35.13%; =0.145; =1.257). Таким образом, на межпопуляционную изменчивость у A. sibirica приходится 35.77% генетического разнообразия. Наибольшее генетическое расстояние отмечено между As2 и As3 (0.290), а наименьшее между As1 и As2 (0.166). Кластерный анализ показал, что As1 генетически ближе к As2. Третья популяция (As3) находится на большем генетическом расстоянии от первых двух популяций и характеризуется наименьшей степенью родства с ними. Среднее значение индексов разнообразия Шеннона () в изученных популяциях A. sibirica, рассчитанные по ISSR-праймерам, составило 0.292. Выше этот показатель в As3. Индекс Шеннона, рассчитанный на общую популяцию A. sibirica, равен 0.504. На долю внутрипопуляционного генетического разнообразия A. sibirica приходится 59.55%, а на долю межпопуляционного – 40.45% разнообразия. Таким образом, молекулярно-генетический анализ двух видов рода Adonis показал, что полиморфизм ISSR-маркеров высок у обоих видов (91.74% у A.vermalis и 89.19% у A. sibirica). У A. vernalis большая часть генетической изменчивости (по данным G-статистики) является внутрипопуляционной (58.78%), на межпопуляционную изменчивость приходится 41.22 % генетического разнообразия. У A. sibirica сохраняется та же тенденция: внутрипопуляционная изменчивость составляет большую часть (64.20%), на межпопуляционную изменчивость данного вида приходится 35.77% генетического разнообразия. Среди изученных популяций A. vernalis тенденция к обеднению генофонда отмечена в Av2 (
=34.86%; =0.122; =1.199; =7) и Av3 (=40.37%; =0.139; =1.125). Среди изученных популяций A. sibirica деградация генофонда выявлена у As1 (=35.13%; =0.145; =1.257). Данная популяция находится в критическом состоянии и требует экстренных мер охраны. 4.2. Анализ генетической изменчивости Adenophora lilifolia (L.) A.DC.
У Ad. lilifolia, выявлено 56 амплифицированных ISSR-фрагментов ДНК, из которых 46 были полиморфными. Число амплифицированных фрагментов ДНК в общей выборке растений варьировало в зависимости от праймера от 9 (праймеры М8, Х10) до 15 (праймер М1), а их размеры – от 279 до 1370 пн. В среднем один праймер инициировал синтез 11 фрагментов ДНК. Доля полиморфных локусов в общей выборке в зависимости от ISSR-праймера колебалась от 75.00% до 90.91% и, в среднем, составила
=82.14 %. Данный показатель выше в Ad.lil.1 и ниже в Ad.lil.4. Ожидаемая гетерозиготность по локусам Ad. lilifolia составила 0.228. Абсолютное число аллелей на локус равно 1.930, а эффективное число аллелей на локус () – 1.412. Ожидаемая гетерозиготность выше в Ad.lil.3 и значительно ниже в Ad.lil.4. Абсолютное число аллелей (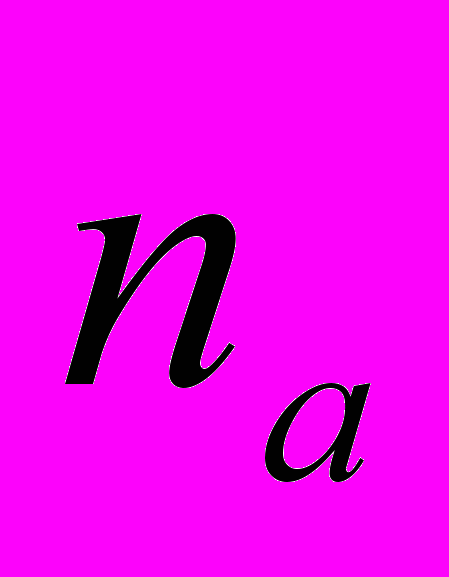 ) на локус выше в Ad.lil.1, а эффективное число аллелей на локус () и ожидаемая гетерозиготность () выше в Ad.lil.3. Наименьшие значения эти показатели имеют в Ad.lil.4. Ожидаемая доля гетерозиготных генотипов на общую популяцию Ad. lilifolia () составила 0.270; а ожидаемая доля гетерозиготных генотипов в субпопуляциях () равна 0.228. Итак, среднее выборочное генетическое разнообразие по популяциям ниже, чем общее генетическое разнообразие в общей популяции. Коэффициент подразделенности популяций () показал, что на межпопуляционную компоненту Ad. lilifolia приходится 15.51% разнообразия (Боронникова, 2009г).
) на локус выше в Ad.lil.1, а эффективное число аллелей на локус () и ожидаемая гетерозиготность () выше в Ad.lil.3. Наименьшие значения эти показатели имеют в Ad.lil.4. Ожидаемая доля гетерозиготных генотипов на общую популяцию Ad. lilifolia () составила 0.270; а ожидаемая доля гетерозиготных генотипов в субпопуляциях () равна 0.228. Итак, среднее выборочное генетическое разнообразие по популяциям ниже, чем общее генетическое разнообразие в общей популяции. Коэффициент подразделенности популяций () показал, что на межпопуляционную компоненту Ad. lilifolia приходится 15.51% разнообразия (Боронникова, 2009г). Таким образом, при молекулярно-генетическом анализе Ad. lilifolia установлено, что доля полиморфных локусов и абсолютное число аллелей выше в Ad.lil.1, а эффективное число аллелей и ожидаемая гетерозиготность – в Ad.lil.3. Все выше перечисленные показатели генетического разнообразия достоверно ниже в Ad.lil.4.
4.3. Анализ генетической изменчивости популяций
Digitalis grandiflora Mill.
При ISSR-анализе структурных элементов генома D. grandiflora выявлено 111 амплифицированных фрагмента ДНК, из которых 90 были полиморфными (81.08%). Число амплифицированных фрагментов ДНК в общей выборке растений варьировало в зависимости от праймера от 15 (X10) до 24 (M9), а их размеры – от 200 до 2200 пн. В среднем, при ISSR-анализе D.grandiflora один праймер инициировал синтез 22.2 фрагментов ДНК. Число полиморфных фрагментов в общей выборке варьировало от 14 до 22. Доля полиморфных локусов и в Dg1 и в Dg2 составила 66.36%, а в Dg3 – 50.45%. Ожидаемая гетерозиготность (
) D. grandiflora равна 0.237. Данный показатель выше в Dg1 (0.207). Абсолютное число аллелей на локус () у D. grandiflora равно 1.991, а эффективное число аллелей на локус () – 1.394. Абсолютное число аллелей на локус ниже в Dg1, а эффективное число аллелей на локус ниже в Dg2 и Dg3. Число редких аллелей у D. grandiflora выше, чем у других изученных видов и равно 18. Наименьшее их число отмечено в Dg1 (=3), а наибольшее – в Dg3 (=17). Анализ генетической структуры популяций показал, что ожидаемая доля гетерозиготных генотипов на всю подразделенную популяцию D. grandiflora (
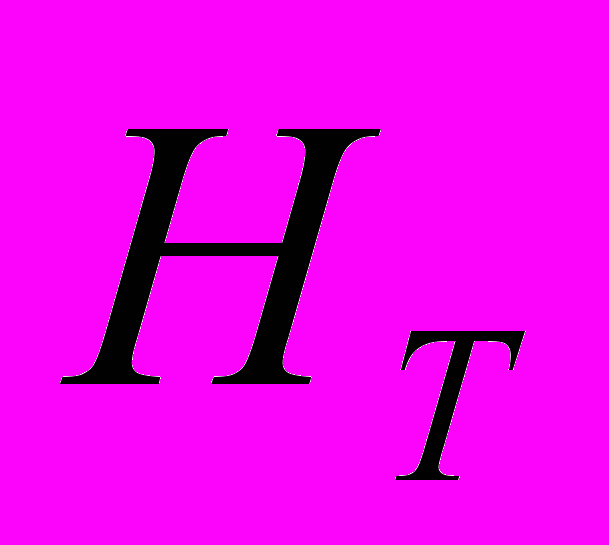 ) равна 0.261, а в субпопуляциях (
) равна 0.261, а в субпопуляциях ( ) – 0.190. Показатель генетической подразделенности популяций () D. grandiflora равен 0.272. Таким образом, на межпопуляционную изменчивость у данного вида приходится 27.22% генетического разнообразия. Среднее значение индексов разнообразия Шеннона () в изученных популяциях D. grandiflora, рассчитанные по ISSR-праймерам, составило 0.292. Кластерный анализ показал, что Dg2 генетически ближе к Dg3 (
) – 0.190. Показатель генетической подразделенности популяций () D. grandiflora равен 0.272. Таким образом, на межпопуляционную изменчивость у данного вида приходится 27.22% генетического разнообразия. Среднее значение индексов разнообразия Шеннона () в изученных популяциях D. grandiflora, рассчитанные по ISSR-праймерам, составило 0.292. Кластерный анализ показал, что Dg2 генетически ближе к Dg3 (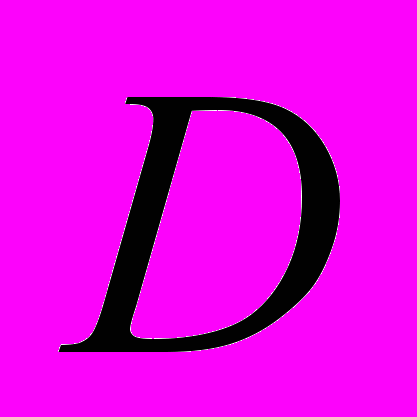 =0.076). Dg1 находится на большем генетическом расстоянии от Dg2 (=0.269) и Dg3 (=0.329). Среди изученных популяций D. grandiflora тенденция к обеднению генофонда отмечена в Dg3 (=50.45%; =0.181; =1.281).
=0.076). Dg1 находится на большем генетическом расстоянии от Dg2 (=0.269) и Dg3 (=0.329). Среди изученных популяций D. grandiflora тенденция к обеднению генофонда отмечена в Dg3 (=50.45%; =0.181; =1.281). Таким образом, ISSR-маркеры позволяют получить ценные сведения о генетическом полиморфизме участков генома, фланкированных микросателлитными повторами, определить основные параметры генетического разнообразия и дифференциации популяций, на основании которых установить основные характеристики генофондов редких и исчезающих видов растений.