
Шестопалов александр михайлович экологический полиморфизм и территориальная значимость различных вирусных патогенов
| Вид материала | Автореферат диссертации |
- Волков Александр Михайлович учебно-методический комплекс, 385.93kb.
- Волков Александр Михайлович учебно-методический комплекс, 441.18kb.
- Нормативные документы по биотерроризму, 1760.66kb.
- Берлянт Александр Михайлович. Картография [Текст]: учебник, 552.83kb.
- Махнев Дмитрий Анатольевич, Нечепоренко Юрий Алексеевич, Сабиров Роман Равильевич,, 173.84kb.
- Т н. Никулин Олег Алексеевич, Новиков Юрий Михайлович, Пивник Александр Васильевич,, 158.75kb.
- Русская Православная Церковь Заграницей»: 19 ноября 1920 года, на борту парохода «Великий, 362.42kb.
- Астаны Александр Васильевич убаськин. Александр Васильевич специалист хорошо известный, 45.11kb.
- Петрухин Станислав Михайлович, Жеребцов Алексей Юрьевич, Комогорцев Игорь Борисович,, 37.1kb.
- Экономическая оценка финансового состояния, 81.64kb.
2.2.2. Эколого-эпизоотологические и молекулярно-биологические характеристики актуальных вирусных патогенов с эволюционных позиций их экологического
полиморфизма и территориальной значимости (в условиях юга Западной Сибири)
2.2.2.1. Вирус бешенства
Эколого-эпизоотологические характеристики
Согласно официальной статистике, бешенство среди животных разных видов в период с 1997 по 2009 год было зарегистрировано более чем в 2/3 субъектов РФ.
Эпизоотическая ситуация бешенства на юге Западной Сибири всегда определялась функционированием Западно-Сибирско-Казахстанского природного очага с циркуляцией в нем возбудителя среди диких животных и активным вовлечением в эпизоотический процесс домашних и сельскохозяйственных животных.
В Западной Сибири в восьмидесятые годы в динамике проявления эпизоотического процесса бешенства отмечали достаточно резкие колебания. Наивысший подъем эпизоотии наблюдали в 1987-1988 годах. Период до 1997 года характеризовался депрессией очага.
Эпизоотическая ситуация бешенства резко осложнилась в конце 1997 года. В течение 1998 года в Новосибирской области бешенство среди диких плотоядных было зарегистрировано в 90 населенных пунктах. В 1999 году – среди диких плотоядных было зарегистрировано в 22 пунктах, среди сельскохозяйственных животных – в 12 и среди домашних животных – в 8 пунктах.
В 1999 году, по сравнению с 1998 годом, произошло снижение числа неблагополучных пунктов по бешенству животных, в том числе и среди диких плотоядных. Если в 1998 году в целом по области было зарегистрировано 162 неблагополучных пункта по бешенству, то в 1999 году – только 45 пунктов. Следует отметить, что в Новосибирской области в 1998-99 годах практически во всех неблагополучных районах была проведена пероральная иммунизация против бешенства диких плотоядных (вакцины "Лисвульпен" и "Синраб").
По итогам 2000 года эпизоотическая ситуация по бешенству в Новосибирской области продолжала улучшаться: было зарегистрировано всего 10 неблагополучных пунктов в 4 районах.
Однако с 2001 года эпизоотическая ситуация обострилась. В 2001 году было зарегистрировано уже 34 неблагополучных пункта в 7 районах, в 2002 году – 163 неблагополучных пункта в 25 районах. В 2003 году в области было зарегистрировано 87 неблагополучных по бешенству пунктов (в 23 районах), в 2004 – 130 (в 27 районах), в 2005 – 49 (в 15 районах), в 2006 – 70 (в 17 районах) и в 2007 – 119 (в 20 районах). В 2008 и 2009 годах эпизоотическая ситуация бешенства оставалась напряженной. Было зарегистрировано 48 неблагополучных по бешенству пунктов.
Таким образом, в период 1997–2009 гг. в Новосибирской области наблюдалось волнообразное изменение уровня заболеваемости с преобладанием бешенства диких животных, но, в целом, сложившуюся ситуацию можно оценить как закономерное проявление эпизоотии, характеризующееся 3–5 летней цикличностью.
Периодичность проявления бешенства, в первую очередь, связана с экологией животных – носителей вируса. Биологический цикл их активности и особенности мест обитания обусловливают сезонные колебания заболеваемости бешенством среди животных.
Контакт с больными дикими животными является причиной заражения домашних и сельскохозяйственных животных. Вспышкам бешенства в животноводческих хозяйствах часто предшествовали случаи появления в этих пунктах диких лисиц. В Новосибирской области в период 1997 – 2009 годов в эпизоотический процесс, наряду с дикими, были вовлечены домашние и сельскохозяйственных животные (рис. 1).
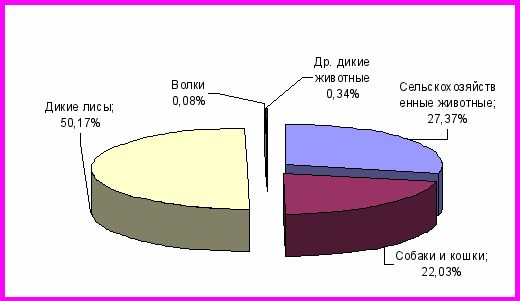
Рисунок 1 – Распределение заболевших бешенством сельскохозяйственных и диких животных, собак и кошек за 1997-2009 годы по Новосибирской области.
Анализ эпизоотических данных показал, что на долю городского бешенства приходится 18,5% от общего количества случаев болезни, зарегистрированных в Новосибирской области за изучаемый период. В городе бешенство было диагностировано у кошек и собак. Особо следует отметить значительный удельный вес бешенства кошек в 1998 – 1999 годы. На долю животных этого вида пришлось 11,15% от общего количества случаев бешенства животных за изучаемый период. В настоящее время все домашние животные подлежат обязательной вакцинации против бешенства. Возможно, благодаря таким мероприятиям (увеличение объемов вакцинации в три раза), удалось добиться заметного снижения удельного веса бешенства среди домашних животных.
Спонтанная зараженность вирусом бешенства лисиц без видимых признаков
болезни на стационарно неблагополучной территории
Показатель спонтанной зараженности лисиц в Западной Сибири в разные периоды колебался от 0,0 до 5,6%. В связи с этим особый интерес для нас представляло исследование популяции диких лисиц, которые являются основным резервуаром вируса бешенства в природе, на наличие возбудителя данной болезни. Было проведено исследование 302 образцов головного мозга лисиц, добытых на территории Алтайского края. Спонтанная зараженность диких лисиц, добытых на стационарно неблагополучной по бешенству территории, была подтверждена различными диагностическими методами более чем у 2,3% особей из числа обследованных.
Изучение носительства лиссавирусов у летучих мышей,
обитающих на юге Западной Сибири
Исследованию на носительство лиссавирусов были подвергнуты рукокрылые, обитающие как в горной, так и в степной части юго-востока Западной Сибири. Всего было собрано и обследовано 288 проб от летучих мышей 9 видов.
При исследовании материала методом ОТ-ПЦР в 34 образцах (18,27%) удалось выявить фрагменты РНК, специфичные для лиссавирусов. У трех полученных ПЦР–продуктов были определены нуклеотидные и выведены аминокислотные последовательности. Филогенетический анализ показал, что все выделенные фрагменты относятся к генотипу 1 лиссовирусов, что характерно для летучих мышей Американского континента. В Евразии от летучих мышей в основном выделяют European bat lyssavirus 1 (EBLV1) и 2 (EBLV2). Что касается Сибири, ранее здесь регистрировались лишь единичные случаи изоляции лиссавирусов от этих животных без изучения генотипа.
Используя материал, положительный в ПЦР-анализе, было проведено 3 пассажа на беспородных белых мышах. На 5-21 день после заражения у 1–2 мышей из группы развивалось заболевание с паралитическими проявлениями. Однако все эти проявления с каждым пассажем были выражены слабее и исчезали к третьему пассажу. Выделить изоляты вируса бешенства от летучих мышей нам не удалось.
Таким образом, нами было показано наличие РНК лиссавирусов генотипа 1 у летучих мышей, обитающих в Новосибирской области и Алтайском крае.
Молекулярно-биологические характеристики вируса бешенства
Изучение молекулярно-генетических характеристик лиссавирусов, циркулирующих
у различных видов животных на территории юга Западной Сибири,
с позиций их эпизоотологической значимости
Для молекулярно-генетических исследований были выбраны образцы материала от различных видов диких и домашних животных, полученные в период 2002-2008 гг., а также от человека, умершего от бешенства. Всего с помощью ПЦР было исследовано 166 проб.
После подтверждения диагноза методом ОТ-ПЦР, для 31 пробы были определены нуклеотидные последовательности гена, кодирующего NP белок, и выведены аминокислотные последовательности. При филогенетическом анализе было выявлено, что все вирусы бешенства, выделенные нами на юге Западной Сибири, группируются в одну филогенетическую группу (рис. 2), но полученные изоляты можно разделить на два основных кластера. Первый из них формируют лиссавирусы от летучих мышей, барсука, лисицы и человека (BAT-YN1, BAT-BN1, BAT-BА, BAT-PА, BADG-4983, MAN-44, FOX-3728) и фиксированные штаммы вирусов бешенства CVS и HEP-Flury. Второй кластер представлен изолятами вирусов бешенства, выделенными в 2004 году от трех собак, лошади и быка (DOG-33650, DOG-35093, DOG-49142, HOR-19549, COW-38442). Эти два кластера имеют достоверный индекс поддержки. Формально, можно сгруппировать и третий кластер, в который можно отнести вирусы, выделенные от животных, заболевание которых произошло в 2003 и 2008 годах. Но третий кластер не имеет достоверного индекса поддержки. Сравнительный анализ нуклеотидных последовательностей фрагмента гена нуклеопротеина со штаммами, представленными в базе данных
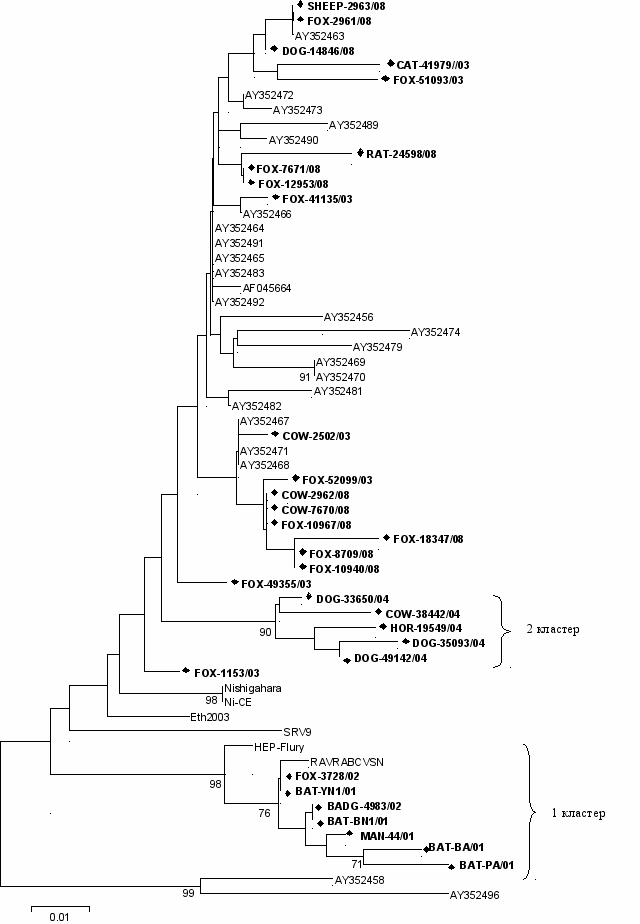 Рисунок 2 – Филогенетическое дерево, построенное на основании фрагмента гена нуклеопротеина (572-776 п.н.) изолятов лиссавирусов, выделенных нами на ЮЗС (выделены жирным шрифтом) со штаммами, представленными в базе данных GenBank
Рисунок 2 – Филогенетическое дерево, построенное на основании фрагмента гена нуклеопротеина (572-776 п.н.) изолятов лиссавирусов, выделенных нами на ЮЗС (выделены жирным шрифтом) со штаммами, представленными в базе данных GenBank GenBank, показал, что на юге Западной Сибири циркулируют близкородственные генетические варианты вируса бешенства, относящиеся к генотипу 1.
Итогом работы по изучению вируса бешенства явилось подтверждение главной роли диких плотоядных (53,0%) в резервации и распространении рабической инфекции в природных очагах на юге Западной Сибири. Кроме диких плотоядных установлено наличие вируса бешенства у барсуков, косуль, зайцев, сурков, бобров и мышевидных грызунов. Установлено, что периодичность эпизоотий бешенства на юге Западной Сибири в течение 1997–2008 годов составила 3–5 лет. Была установлена зависимость эпизоотологической роли основных резервуаров вируса бешенства от направленности и интенсивности антирабических мероприятий. Увеличение в 3 раза объемов иммунизации домашних животных обеспечило снижение на 46,0% случаев бешенства у этой категории животных. Ежегодная пероральная иммунизация диких лисиц в природных эпизоотических очагах эффективно снижала проявление эпизоотии бешенства на 36,0%. Выявлены фрагменты гена нуклеопротеина лиссавирусов генотипа 1 у насекомоядных летучих мышей, обитающих на территории юга Западной Сибири. Показано близкое генетическое родство лиссавирусов, циркулирующих на территории юга Западной Сибири в популяциях животных разных видов, которые формируют две основные филогенетические группы в пределах генотипа 1.
2.2.2.2. Вирус Западного Нила
Эколого-эпизоотологические характеристики
В настоящей главе диссертационной работы экосистемы юга Западной Сибири были рассмотрены с точки зрения существования в них условий, благоприятных для поддержания циркуляции ВЗН, не связанной с сезонным заносом вируса. При рассмотрении биотических сообществ основное внимание уделялось животным, способным вовлекаться в трансмиссивный цикл ВЗН (птицы, мелкие млекопитающие, кровососущие комары и клещи). Из абиотической компоненты рассматривались физические факторы, влияющие на видовой состав, численность и активность членистоногих переносчиков – температура, влажность и скорость ветра, а также температурный диапазон, определяющий эффективность репликации вируса в их организме. Наличие вирусных маркеров у позвоночных животных и кровососущих членистоногих, участвующих в трансмиссивном цикле ВЗН, а также существование благоприятных условий для репликации вируса свидетельствуют о циркуляции ВЗН в цепочке птица↔комар (клещ)→другое позвоночное животное в пределах определенных биотопов.
Вовлечение в трансмиссивный цикл ВЗН случайных хозяев вируса, не являющихся компонентами природных экосистем – сельскохозяйственных животных, людей – приводящее к развитию заболевания и (или) формированию иммунитета, является косвенным доказательством циркуляции вируса в местных популяциях кровососущих членистоногих и формирования сезонных очагов циркуляции. При составлении характеристики биотопов, выбранных для сбора полевого материала, учитывалось наличие перелетных птиц, мелких млекопитающих и членистоногих кровососущих, способных вовлекаться в трансмиссивный цикл ВЗН, а также антропогенная нагрузка, влияющая на численность и видовой состав животных, участвующих в циркуляции ВЗН. Для большинства выбранных биотопов было характерно присутствие диких перелетных птиц и кровососущих членистоногих, способных вовлекаться в циркуляцию ВЗН. В то же время, биотопы различались по экологическим условиям, влияющим на обитание и активность участников трансмиссивного цикла, а это, в свою очередь, может влиять на циркуляцию вируса в конкретном биотопе.
Подавляющее большинство добытых во всех ландшафтных зонах птиц относились к воробьеобразным, т.е. к ВЗН-восприимчивым. Сравнение распределения вирусных маркеров по сезонам и местам сбора материала выявило определенные различия. При сопоставлении сезонов 2003-2004 гг. в Северной Кулунде было отмечено увеличение степени инфицирования птиц в сезоне 2004 г. Степень инфицирования птиц, добытых в южной лесостепи в разные периоды сезона 2004 г. оказалась различной как при сравнении между периодами сборов, так и при сравнении с тем же сезоном в Северной Кулунде. Среди птиц, добытых летом, доля особей, позитивных по наличию одного из главных маркеров ВЗН белка Е, была выше, чем у птиц, добытых весной. В целом, по выявлению гликопротеина и вирусной РНК процент инфицированных птиц в 2004 г. в южной лесостепи был меньше, чем у птиц, добытых в Северной Кулунде. РНК ВЗН не была выявлена ни в одном образце внутренних органов, вирусный белок Е не был обнаружен ни в одном образце головного мозга птиц, добытых в сезоны 2006 и 2007 гг. С большой частотой гликопротеин Е обнаруживался в печени птиц, а в сезоне 2006 г. – в селезенке у одной птицы. Суммарная частота выявления этого маркера составила в сезоне 2006 г. 43,2%, в сезоне 2007 г. – 42,4%.
Отдельно можно отметить особенности распределения гликопротеина Е и РНК ВЗН в разных группах птиц. В группе грачей, добытых в начале мая на юге Барабинской лесостепи, выявление белка Е совпадало с выявлением вирусной РНК, в то время как в головном мозге большей части птиц, добытых там же в летние месяцы, выявлялся гликопротеин Е, а процент птиц с двумя маркерами был в несколько раз меньше. Высокий процент совпадения по обоим маркерам отмечался и у птиц 2004 г. в Северной Кулунде. Большая частота выявления гликопротеина и вирусной РНК у этих птиц свидетельствует о том, что в организме птиц имела место продуктивная ВЗН-инфекция с попаданием вируса в ЦНС и последующей репликацией в головном мозге. В пользу этого говорит тот факт, что часть грачей с положительными находками гликопротеина и РНК ВЗН были найдены мертвыми, хотя нами за время сборов не было отмечено больших эпизоотий среди этих птиц.
Таким образом, по результатам анализа маркеров ВЗН, большинство обследованных птиц как мигрирующих, так и оседлых видов ранее имели контакт с вирусом Западного Нила. Результатом этого у части птиц стало развитие продуктивной ВЗН-инфекции с попаданием вируса в ЦНС, которая могла стать причиной их гибели; у другой части птиц имела место абортивная инфекция, ограниченная внутренними органами. В таком случае, при различном исходе инфекции у птиц, эффективность передачи вируса от них другим участникам трансмиссивного цикла (как позвоночным, так и членистоногим) также должна различаться и коррелировать с распределением в организме этих животных вирусных маркеров. Для проверки этого предположения был проведен анализ внутренних органов млекопитающих на наличие и особенности распределения маркеров ВЗН.
Млекопитающие
Из отловленных мелких млекопитающих в степной зоне большую часть составили мышевидные грызуны. Распределение маркеров ВЗН у зверьков степных и лесостепных биотопов отличалось от такового для подзоны осиново-березовых лесов. Наиболее часто гликопротеин Е и вирусная РНК встречались в головном мозге животных лесостепных биотопов, причем вирусный антиген был обнаружен во всех образцах головного мозга. Почти полное совпадение в выявлении гликопротеина Е и РНК наблюдалось у грызунов, отловленных во влажном колке с действующей колонией грачей, далее по частоте встречаемости маркеров следовали животные, отловленные на остепненном лугу и прибрежной полосе р. Чулым. Этот факт свидетельствует о протекании в организме всех обследованных животных продуктивной ВЗН-инфекции с попаданием вируса в ЦНС.
В Северной Кулунде чаще всего вирусные маркеры встречались у зверьков, отловленных на заросшем поле у оз. Кротово. На втором месте по частоте встречаемости маркеров следовали животные, отловленные на участке полынно-злаковой степи, несмотря на отсутствие в этих биотопах благоприятных условий для активности комаров. Этот факт можно объяснить как перемещением животных в биотопы, где активны комары, так и возможным участием в передаче вируса преимагинальных стадий клещей рода Dermacentor. В целом, частота встречаемости обоих вирусных маркеров (белок Е/РНК) у мелких млекопитающих Северной Кулунды составила 40,0%/12,7%. Таким образом, среди мелких млекопитающих Северной Кулунды доля животных с протеканием продуктивной ВЗН-инфекции заметно меньше по сравнению с животными южной лесостепи.
Кровососущие членистоногие
Комары
Хотя маркеры ВЗН у комаров нами выявлены не были, в Северной Кулунде и на юге Барабинской лесостепи имеются виды комаров, способные участвовать в трансмиссивном цикле ВЗН и для которых характерна высокая численность в некоторых биотопах. Это создает предпосылки для поддержания циркуляции вируса в период их активности.
Клещи
Результат анализа на распределение маркеров ВЗН у иксодовых клещей из разных биотопов в сезоне 2006 г. показали, что наиболее часто белок Е и РНК ВЗН встречались у имаго Ix. рersulcatus и Ix. pavlovskyi с участием всех возрастных стадий.
Молекулярно-биологические характеристики вариантов ВЗН
Из части образцов, в которых были выявлены вирусные маркеры, с помощью ОТ-ПЦР были получены фрагменты кДНК, соответствующие участку гена белка Е ВЗН, которые были секвенированы. Секвенированные образцы имели следующее происхождение: два образца от серых ворон из Северной Кулунды 2003 г. (ChCorvus10-03, ChCorvus31-03), три образца от грачей из южной лесостепи 2004 г. (ChRook7-04, ChRook14-04, ChRook27-04), образец от красной полёвки из южной лесостепи 2004 г. (ChRodent-04), два образца от заболевших людей (Пациенты №1 и №3, ChiWS-04 и HumWS-04 соответственно) и два пула имаго клещей Ix. persulcatus и Ix. pavlovskiy из подзоны осиново-березовых лесов 2006 г. (Tomsk-1, Tomsk-2).
Генотипирование исследуемых нуклеотидных последовательностей позволило идентифицировать эти последовательности как относящиеся к генотипу Ia вируса Западного Нила. Западносибирские варианты ВЗН 2003-2004 гг. и 2006 г. оказались высоко гомологичными (не более 1-3% расхождения) с волгоградским штаммом VLG-4 ВЗН. При анализе филогенетического дерева (рис.3) можно видеть, что исследованные варианты характеризуются невысоким уровнем генетических отличий. Можно также видеть, что исследованные западносибирские варианты ВЗН не имеют гостальной специфичности. Филогенетический анализ показал, что выявленные в 2003-2004 гг. и 2006 г. последовательности ВЗН кластеризуются вместе с другими современными российскими изолятами ВЗН. Это говорит о том, что эти варианты вируса генетически связаны со штаммами ВЗН, изолированными на юге Европейской части России. В отношении вариантов, выделенных от птиц, полученные результаты согласуются с орнитологическими данными, и это может служить доказательством заноса ВЗН перелётными птицами на юг Западной Сибири с эндемичных территорий на юге РФ. Поскольку для клещей Ix. persulcatus и особенно Ix. pavlovskiy, птицы являются прокормителями, выявление вариантов ВЗН в клещах свидетельствует об участии птиц в заносе вируса и передаче членистоногим переносчикам. Генетическая близость исследуемых вариантов с патогенными для людей штаммами ВЗН юга России позволяет предполагать, что западносибирские варианты ВЗН обладают эпидемическим потенциалом и могут быть причиной заболевания ВЗН людей на юге Западной Сибири.
Из филогенетического дерева можно видеть, что варианты ВЗН от серых ворон и грача (ChRook27-04) образуют одну группу. По сравнению с прототипным штаммом VLG-4, они характеризуются общей консервативной заменой Arg94Lys, которая также свойственна вариантам ВЗН, выделенным в различных регионах мира и относящимся к генотипам Ib, II, III и IV, но не выявлена у чирков и грачей юга Западной Сибири в 2002 году (генотип Ia). Каждый вариант от серых ворон имеет индивидуальные консервативные уникальные замены – у одного обнаружена Ala73Thr, у другого – Pro89Ala, также не встречавшиеся ранее у других вариантов вируса. В целом эта группа характеризуется консервативной уникальной заменой Pro76Ala и значимой заменой Leu69Hys, выявленной у грача и вороны, также являющейся уникальной. Эти замены выявлены в вариантах ВЗН от птиц, принадлежащих к видам с разным миграционным статусом, добытых в разное время и в разных ландшафтных зонах, что может указывать на общий источник инфицирования птиц ВЗН, а также на возможное участие организма хозяина в воздействии на изменчивость вируса.
Таким образом, генотипирование позволило характеризовать западносибирские варианты ВЗН как генетически близкие между собой и патогенным для людей штаммам ВЗН, изолированным на юге России. Сравнительный аминокислотный анализ выявил замены только в вариантах от восприимчивых к ВЗН врановых птиц, несмотря на значительную генетическую близость этих вариантов ВЗН с вариантами от других хозяев. Полученные результаты свидетельствуют о способности вируса адаптироваться к разным хозяевам в различных ландшафтно-экологических условиях, а также об эпидемическом потенциале ВЗН, циркулирующего на юге Западной Сибири.
Итогом по изучению циркуляции вируса западного Нила на территории юга Западной Сибири явилось выявление циркуляции ВЗН среди воробьеобразных птиц (Passeriformes), мигрирующих и оседлых видов, в степной, лесостепной зонах и подзоне осиново-березовых лесов юга Западной Сибири. Также было установлено, что в циркуляцию ВЗН в степной, лесостепной зонах и подзоне осиново-берёзовых лесов юга Западной Сибири вовлекаются мелкие млекопитающие – грызуны, насекомоядные и мелкие хищники. Среди комаров-эудоминантов выявлены эпидемиологически значимые виды, способные принимать участие в трансмиссивном цикле ВЗН. Показано вовлечение в циркуляцию ВЗН имагинальных и преимагинальных стадий иксодовых клещей Ix. persulcatus и Ix. pavlovskiy в подзоне осиново-берёзовых лесов юга Западной Сибири. В степной, лесостепной зонах и подзоне осиново-берёзовых лесов юга Западной Сибири показано формирование природных очагов ВЗН, не связанных с сезонным заносом. Было установлено вовлечение в трансмиссивный цикл ВЗН на юге Западной Сибири случайных участников – сельскохозяйственных животных и людей.
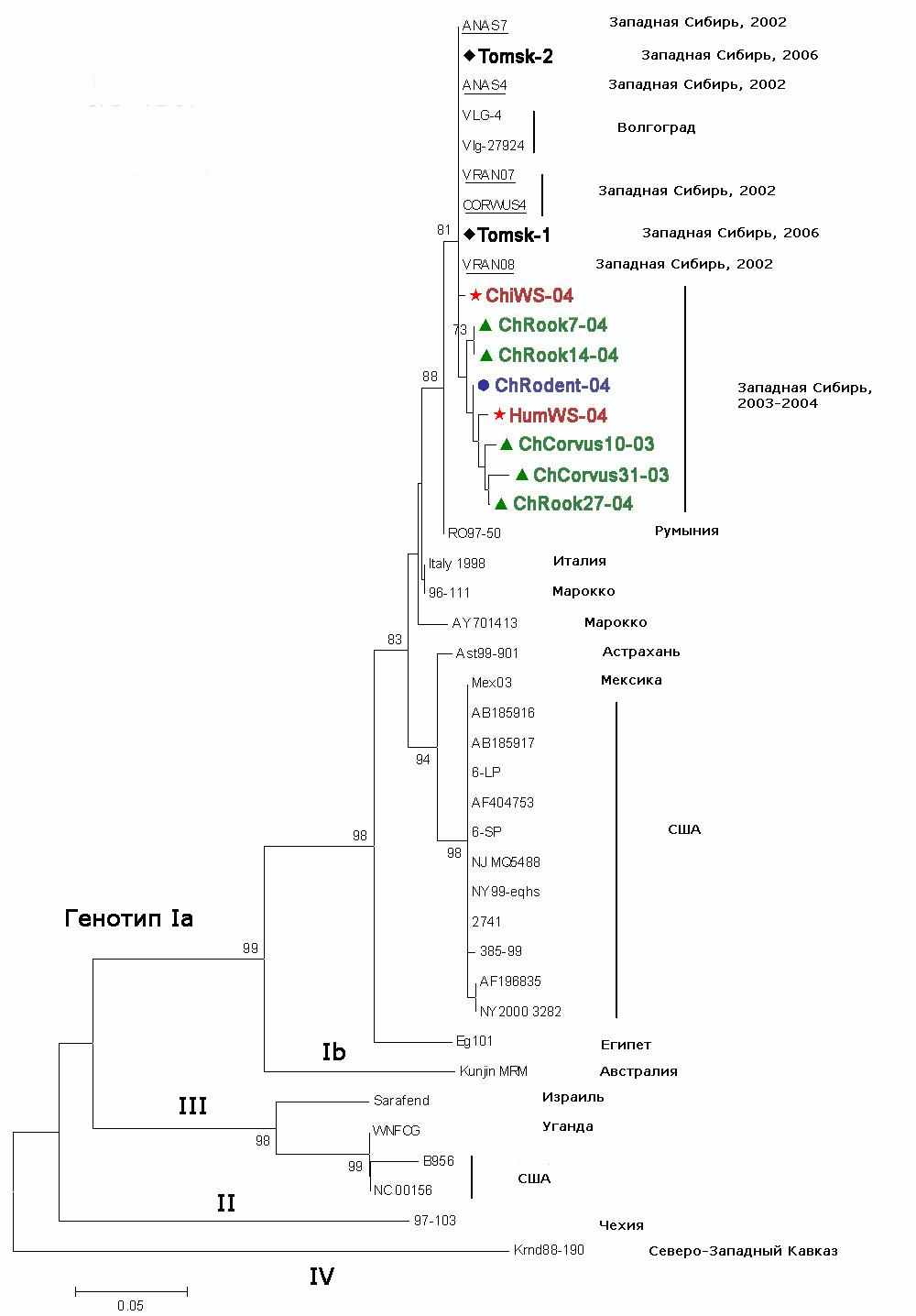
Рисунок 3 – Филогенетическое дерево полевых изолятов и штаммов ВЗН, построенное на основании сравнения нуклеотидных последовательностей фрагмента гена белка Е (322 нуклеотида) со штаммами, представленными в базе данных GenBank .
Примечания:
 – птицы (серые вороны и грачи);
– птицы (серые вороны и грачи);  – красная полёвка;
– красная полёвка;  - - люди;
- - люди;  – клещи Ix. persulcatus и Ix. Pavlovskiy (фрагменты РНК выявленные нами у различных хозяев на ЮЗС).
– клещи Ix. persulcatus и Ix. Pavlovskiy (фрагменты РНК выявленные нами у различных хозяев на ЮЗС).Секвенирование и последующий анализ фрагментов гена белка Е ВЗН, выделенного от разных хозяев, показал, что западносибирские варианты ВЗН принадлежат генотипу Iа ВЗН, являются генетически близкими между собой и близки с патогенными для людей штаммами ВЗН, изолированными на юге России.