
И. Я. Яковлева Кафедра анатомии, физиологии и гигиены детей Д. А. Димитриев «Нейрон, его строение и функция» «Общие свойства сенсорных систем» Учебное пособие
| Вид материала | Учебное пособие |
| Для пространственного облегчения Латеральное торможение Реципрокное торможение |
- Физиология сенсорных систем Цель дисциплины, 13.63kb.
- План: Предмет и задачи возрастной анатомии, физиологии и гигиены. Значение возрастной, 69.36kb.
- Время и место проведения Конференция проводится в апреле 2012 года в Нижегородском, 41.09kb.
- Правила гигиены органов зрения, 62.45kb.
- Экзаменационная программа по физиологии высшей нервной деятельности и сенсорных систем., 48.31kb.
- Перечень вопросов для подготовки к итоговой государственной аттестации по специальности, 451.95kb.
- Особенности морфологического и гормонально-метаболического статуса спортсменок-борцов, 533.99kb.
- Учебное пособие раскрывает происхождение и значение более 1000 учебных терминов, 1634.01kb.
- Планирование темы Урок Биосоциальная природа человека и науки, изучающие его, 1716.33kb.
- Л. Н. Шиян Свойства и химия воды. Водоподготовка Учебное пособие, 1119.56kb.
Кроме активации протеинкиназ внутриклеточные эффекты цАМФ реализуются также через три других механизма: систему кальций-кальмодулин (о чем будет сказано ниже), трансметилазную систему и аденозин-5-монофосфат- аденозин. О роли системы кальций-кальмодулин будет сказано ниже. Трансметилазная система обеспечивает метилирование ДНК, всех типов РНК, белков хроматина и мембран, ряда гормонов на уровне тканей, фосфолипидов мембран.
2. Система гуанилатциклаза - цГМФ. Переход мембранной гуанилатциклазы в активную форму происходит не под непосредственным влиянием лиганд–рецепторного комплекса, а опосредованно через ионизированный кальций и оксидантные системы мембран. Типичная стимуляция активности гуанилатциклазы ацетилхолином также реализуется опосредованно через Са++. Под влиянием гуанилатциклазы из ГТФ синтезируется цГМФ, активирующий цГМФ-зависимые протеинкиназы, которые уменьшают скорость фосфорилироваия легких цепей иозина в гладких мышцах стенок сосудов, приводя к их расслаблению. В большинстве тканей биохимические и физиологические эффекты цАМФ и цГМФ противоположны. Так, цАМФ вызывает стимуляцию сокращений сердца, а цГМФ – их торможение. ЦГМФ играет роль в гиперполяризации рецепторов сетчатки глаза под влиянием фотонов света. Ферментативный гидролиз цГМФ осуществляется с помощью специфической фосфодиэстеразы.
3. Система фосфолипаза С – инозитол–3–фосфат.
Лиганд-рецепторный комплекс с участием регуляторного G – белка ведет к активации мембранного фермента фосфолипазы С, вызывающего гидролиз фосфолипидов мембраны с образованием двух вторичных посредников: инозитол–3–фосфата и диацилглицерола. Инозитол – 3 – фосфат вызывает выход Са++ из внутриклеточных депо, в основном из эндоплазматического ретикулума, ионизированный кальций связывается со специализированным белком кальмодулином, что обеспечивает активацию ряда протеинкиназ и фосфорилирование внутриклеточных структурных белков и ферментов. В свою очередь диацилглицерол способствует резкому повышению сродства протеинкинзы С ионизированному кальцию, который активирует ее без участия кальмодулина. Эти процессы также завершаются фосфорилированием других белков. Диацилглицерол одновременно может реализовывать и другой путь опосредования медиаторного эффекта, так как он активирует фофолипазу А-2. Под влиянием последней из мембранных фосфолипидов образуется арахидоновая кислота, являющаяся источником мощных по метаболическим и физиологическим эффектам веществ – простагландинов и лейкотриенов. Необходимо указать, что в разных клетках превалирует один или другой пути образования вторичных посредников, что в конечном счете и определяет физиологический эффект лиганда.
4. Система Са – кальмодулин. Ионизированный кальций поступает в клетку после образования лиганд-рецепторного комплекса либо из внеклеточной среды за счет активирования медленных кальциевых каналов мембраны (как это происходит, например, в миокарде) либо из внутриклеточных депо, под влиянием вышеописанных внутриклеточных процессов. В цитоплазме немышечных клеток кальций связывается со специальным белком-кальмодулином, а в мышечных клетках роль кальмодулина выполняет тропонин С. Связанный с кальцием кальмодулин изменяет свою конформацию, что приводит к активации многочисленные протеинкиназ, обеспечивающих фосфорилирование белков. Другим механизмом действия комплекса кальций - кальмодулин является активация фосфодиэстеразы цАМФ, что подавляет ее эффект как вторичного посредника. Кратковременное увеличение в клетке кальция и ее связывание с кальмодулином является пусковым стимулом для многочисленных физиологических процессов – сокращения мышц, секреции гормонов и выделения медиаторов, синтеза ДНК, изменения подвижности клеток, транспорта веществ через мембраны, изменения активности ферментов.
В значительной части клеток организма присутствуют или могут образовываться почти все из рассмотренных выше вторичных посредников (исключением является цГМФ). Это обуславливает различные взаимосвязи между вторичными посредниками: 1) равноправное участие, когда разные посредники необходимы для медиаторного эффекта; 2) один из посредников является основным, а другой лишь способствует реализации эффектов первого; 3) посредники действуют последовательно (например, инозитол – 3- фосфат обеспечивает освобождение кальция, диацилглиццерол облегчает взаимодействие кальция с протеинкиназой С); 4) посредники дублируют друг друга для обеспечения избыточности с целью надежности регуляции; 5) посредники являются антагонистами, т.е. один из них включает реакцию, а другой тормозит (например, в гладких мышцах сосудов инозитол–3–фосфат и кальций реализуют их сокращение, а цАМФ – расслабление).
Эффект от воздействия биологически активного вещества во многом зависит от того, с каким типом рецепторов он образовал лиганд-рецепторную связь. В группу рецепторов – ионных каналов входят, в частности, Н-холинорецепторы. Вторая группа рецепторов образована рецепторами, сопряженными с ферментами. Для этих рецепторов характерна каталитическая активность одной из образующих их молекул белка. Лиганд – рецепторное взаимодействие приводит к изменению каталитической активности данной субъединицы, а значит и синтеза внутриклеточных вторичных мессенджеров. По такому механизму функционируют рецепторы факторов роста, инсулиновые рецепторы и др. К третьей группе рецепторов относятся рецепторы, сопряженные с G-белками. G-белки – это особые ГТФ-связывающие белки, расположенные на внутриклеточной стороне цитоплазматической мембраны. Посредством этих белков рецепторы могут изменять активность внутриклеточных ферментативных систем и вызывать открытие ионных каналов. К G-сопряженным рецепторам относятся, например, β-адренорецепторы. G-белки представляют собой гетерогенную популяцию, различающиеся за счёт α-субъединицы. Имеется насколько видов G-белков:
-

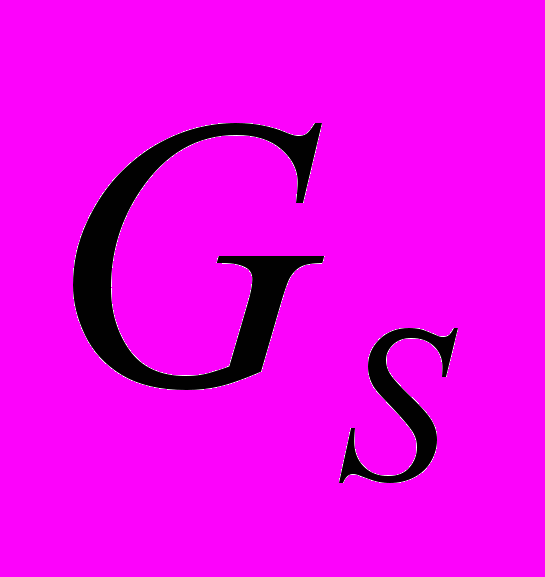 - белки. Оказывают стимулирующее влияние на аднилатциклазу, оказывает ингибирующее влияние на медленный тип кальциевых каналов.
- белки. Оказывают стимулирующее влияние на аднилатциклазу, оказывает ингибирующее влияние на медленный тип кальциевых каналов.
-
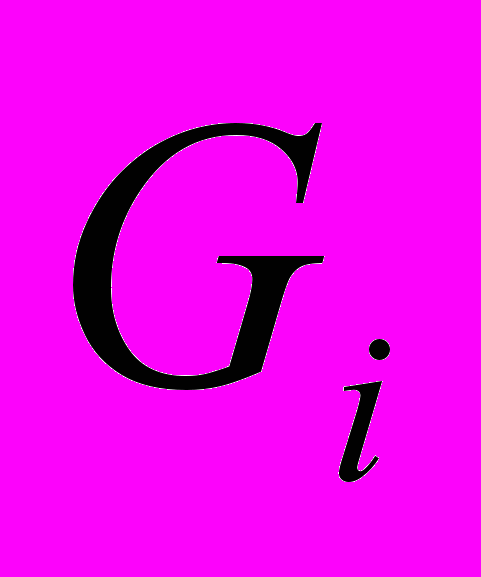 - белки. Оказывают ингибирующее воздействие на аденилатциклазу, активируют цАМФ-фосфодиэстеразу.
- белки. Оказывают ингибирующее воздействие на аденилатциклазу, активируют цАМФ-фосфодиэстеразу.
-
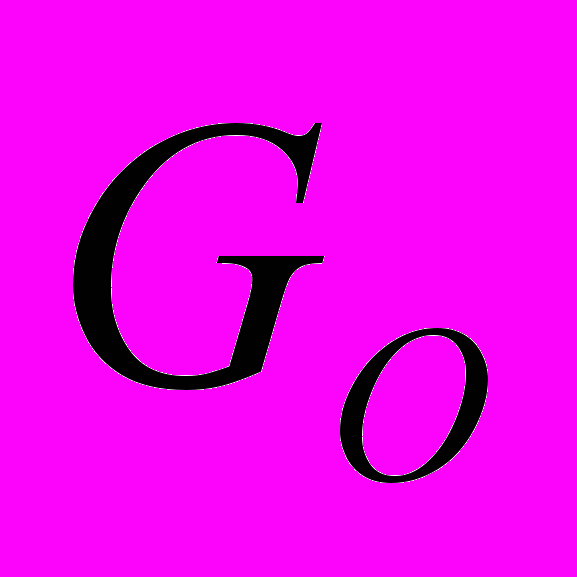 -белки. Их активация приводит к угнетению потенциал-зависимых кальциевых каналов и стимуляции калиевых каналов.
-белки. Их активация приводит к угнетению потенциал-зависимых кальциевых каналов и стимуляции калиевых каналов.
-
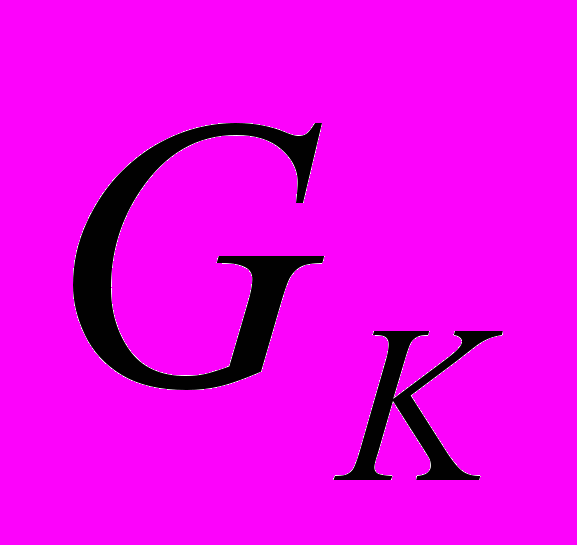 -белки. Вызывают активацию калиевых каналов и угнетение аденилатциклазы.
-белки. Вызывают активацию калиевых каналов и угнетение аденилатциклазы.
-
 -белки. Приводят к активации фосфолипазы С.
-белки. Приводят к активации фосфолипазы С.
Процессы, происходящие при воздействии тормозящих и возбуждающих медиаторов на рецепторы постсинаптической мембраны различны. Под воздействием возбуждающего медиатора на постсинаптической мембране возникает возбуждающий постсинаптический потенциал (ВПСП). Данные изучения ионных механизмов ВПСП указывают на то, что ВПСП представляет собой результат кратковременного повышения проводимости для мелких катионов. Другим вариантом развития событий может быть возникновение торможения. Известны два типа торможения – пресинаптической и постсинаптическое. При пресинаптическом торможении уменьшается или прекращается высвобождение медиатора из пресинаптических нервных окончаний, а при постсинаптическом торможении снижается возбудимость сомы и мембраны дендритов. Тормозящий постсинаптический потенциал (ТПСП) по своей сути представляет собой гиперполяризацию постсинаптической мембраны. Возникновение ТПСП связано с тем, что при действии тормозящего медиатора на постсинаптическую мембрану повышается проницаемость последней для ионов калия и хлора. Торможение, вызываемое ТПСП, обусловлено как гиперполяризацией мембранного потенциала, так и начальным повышением проводимости мембраны.
В качестве гистологического субстрата пресинаптического торможения удобнее всего рассматривать аксо-аксональные синапсы. В ходе проведенных различными учеными исследований было установлено, что снижение количества высвобождаемого медиатора в случае пресинаптического торможения связано с активацией именно этого вида синапсов. В механизме пресинаптического торможения большую роль играет деполяризация первичных афферентов (ДПА). Активация аксо-аксонного синапса сопровождается деполяризацией постсинаптической области. Такую деполяризацию можно зарегистрировать внутриклеточно в первичных афферентах спинного мозга, откуда она и получила свое название. ДПА сопровождается уменьшением амплитуды пресинаптического потенциала в результате инактивации натриевых каналов. В конечном счете, может происходить полная блокада проведения в пресинаптических окончаниях, сопровождающаяся высвобождением очень небольшого количество медиатора.
После того, как медиатор диффундировал через синаптическую щель, дальнейшая его диффузия из этого узкого пространства идет довольно медленно и не способна заметно снизить установившуюся концентрацию.
Важнейшим, с точки зрения физиологии, вопросом является взаимодействие между синапсами. И действительно, первый же шаг от этого элементарного уровня взаимодействия двух клеток возбудимых тканей вверх по шкале размерностей, первый же шаг к пониманию целостной функциональной организации нервной системы требует рассмотрения этого взаимодействия.
Объединяясь между собой посредством синапсов, нейроны формируют изумительные по сложности системы, чьё функционирование и определяет, в конечном счетё, высшие функции центральной нервной системы. Здесь мы рассмотрим лишь наиболее простые из этих нейронных сетей. Эти элементарные нейронные цепи служат, например, для усиления слабых сигналов, уменьшения слишком интенсивной активности, выделения контрастов, поддержания ритмов или сохранения рабочего состояния нейронов путём регулировки их входов. Сама собой напрашивается аналогия с интегральными электрическими цепями – стандартными элементами, которые выполняют наиболее часто повторяющиеся операции, и могут быть включены в схемы самых разнообразных электронных приборов. Эти простейшие нейронные системы расположены во всех отделах головного и спинного мозга и характеризуются некоторым набором структурно функциональных феноменов, к числу которых относится конвергенция, дивергенция, временная и пространственная суммация (облегчение), различные виды торможения и реверберации.
Афферентные волокна, несущие сигнал от периферических рецепторов, сначала входят в спинной мозг в составе дорсальных корешков, а затем ветвятся на много коллатералей, идущих к спинальным нервам. Таким образом один чувствительный нейрон оказывает влияние на несколько других нейронов. Это явление получило название дивергенция. Благодаря дивергенции афферентная информация поступает одновременно к разным участкам ЦНС. Тут следует отметить, что разветвление волокон дорсальных корешков является лишь частным случаем дивергенции. Это явление обнаруживается во всех отделах центральной нервной системы, что позволяет говорить о принципе дивергенции в нейронных сетях.
В качестве структурной и функциональной противоположности дивергенции мы с полным основанием можем рассматривать конвергенцию. При этом явлении несколько нейронов оказывают влияние на функцию одного нейрона. Число таких входов для одного нейрона очень велико (на мотонейроне оканчивается до 6000 коллатералей аксонов), а само явление столь широко представлено в ЦНС, что мы с полным основанием можем говорить о принципе конвергенции в нейронных связях. Благодаря конвергенции возможно интегрирование различным потоков информации на одном нейроне, что является одним из важнейших процессов, происходящих в ЦНС.
В случае, если возбуждающие постсинаптические потенциалы (ВПСП) быстро следуют друг за другом, то они суммируются благодаря своему относительно медленному временно́му ходу, достигая в конце концов порогового уровня. Это повышение возбудимости нейрона в результате последовательно возникающих ВПСП носит название временно́го облегчения. Временное облегчение ответа на активность аксона обусловлено тем, что ВПСП продолжается дольше, чем рефрактерный период аксона. Временное облегчение играет очень важную роль, потому что многие нейронные процессы имеют ритмический характер и, таким образом, могут суммироваться, давая начало надпороговому возбуждению в синапсах.
Для пространственного облегчения характерна суммация ВПСП, возникающих одновременно в разных синапсах. В результате этого возникает распространяющийся потенциал действия – процесс, который не может быть обеспечен одиночным ВПСП. Если рассматривать в целом оба вида облегчения, то можно сказать, что облегчение возникает в том случае, когда количество импульсов больше, чем сумма отдельных входов.
Выше мы уже отмечали, что кроме нейронных цепей, в которых происходит облегчение, существуют и простые тормозные цепи, для которых характерны различные типы торможения. Изучение их мы начнем с возвратного торможения. Эта форма торможения возникает в том случае, когда тормозные вставочные нейроны действуют на те же нервные клетки, которые их активируют. При этом торможение бывает тем глубже, чем сильнее было предшествующее возбуждение. Это позволяет использовать кибернетический принцип обратной связи для описания этих цепей, а само возвратное торможение рассматривать как торможение по принципу обратной связи. Частным случаем возвратного торможения является торможение Реншоу. Этот вид торможение встречается в мотонейронах: мотонейроны посылают коллатерали к вставочным нейронам (клеткам Реншоу), аксоны которых в свою очередь образуют тормозные синапсы на мотонейронах.
Латеральное торможение. При данном виде торможения тормозные вставочные нейроны соединены таким образом, что они влияют не только на возбужденную клетку, но и на соседние клетки с такими же функциями, в которых возбуждение отсутствует или является более слабым. В результате в этих соседних нейронах развивается очень глубокое торможение. Поскольку при этом возбужденный нейрон оказывается со всех сторон окруженным зоной торможения, это явление также называют окружающим торможением. Латеральное торможение играет особенно важную роль в афферентных системах.
Реципрокное торможение. Для изучения данного вида торможения необходимо ввести некоторую терминологию. Гомонимными называются все те нейроны, которые посылают аксоны к одной и той же мышце. Мышцы-агонисты, или синергисты, осуществляют движение в суставе в одном и том же направлении, а мышцы-антагонисты - в противоположных направлениях. Нервные волокна, идущие по направлению к гомонимным мотонейронам, образуют на этих нейронах возбуждающие синапсы, а посредством вставочного нейрона – тормозные синапсы на мотонейроны мышц-антагонистов. Если повышается тонус нервных волокон, идущих к мотонейронам мышц-сгибателей, то они оказывают тормозящие влияние на мотонейрон, иннервирующий мышцу-разгибатель, действующую на тот же сустав. Это торможение имеет большой физиологический смысл, потому что обеспечивает согласование тонуса мышц и облегчает движение в суставе автоматически, без дополнительного произвольного или непроизвольного контроля.