
Итак, у нас остался тип взаимоотношений «-», или конкуренция. Рассмотрим этот тип взаимоотношений между популяциями и особями подробнее
| Вид материала | Лекция |
СодержаниеK1. Вторая линия (описывающая условие стабильности численности «вида-конкурента») пересечет ось ординат в точке со значением K Феноменология динамики численности |
- «Особенности правовых взаимоотношений организации и добровольцев», 122.47kb.
- Ортопедической стоматологии, 200.57kb.
- Iv специфика взаимоотношений в группе умственно одаренных детей, 281.91kb.
- Аналитический доклад, 278.28kb.
- Урока Тема урока Тип, форма урока, 78.15kb.
- «Основы экологии» (8часов), 286.22kb.
- Курс: 4 Тип: контрольная Теория структурно-функционального анализа Введение, 249.9kb.
- Взаимоотношений, 8391.69kb.
- Взаимоотношений, 8391.83kb.
- Личность как социальный тип Содержание, 215.27kb.
Лекция 17
Конкуренция
Итак, у нас остался тип взаимоотношений «– –», или конкуренция. Рассмотрим этот тип взаимоотношений между популяциями и особями подробнее.
Как я уже говорил, взаимодействия между организмами принято подразделять на гомотипические (то есть внутривидовые) и гетеротипические (то есть межвидовые). До сих пор мы говорили преимущественно о гетеротипических реакциях, и это естественно – ведь нас интересовали в большей мере отношения между популяциями, нежели между особями. Впрочем, если в рассматривавшихся нами ранее взаимодействиях гомотипические реакции и встречались (например, каннибализм или паразитирование самцов на самках своего вида), это были достаточно экзотические случаи. В случае же конкуренции ситуация иная: и межвидовая, и внутривидовая, даже внутрипопуляционная, конкуренция – явления, чрезвычайно важные и с точки зрения их распространенности в природе, и с точки зрения их роли в функционировании экосистем, динамики популяций и даже эволюции.
Суть конкуренции состоит в том, что популяции или особи должны распределить между собой некий ограниченный ресурс (или ресурсы), при этом каждая из этих популяций или особей получает этот ресурс в меньшем объеме, нежели если бы она имела доступ к нему в одиночестве. Таким образом и возникает эта формула «– –», причем в случае дефицита жизненно важного ресурса отрицательное взаимовлияние двух конкурентов может оказаться фатальным по меньшей мере для одного из них.
Ресурсы, за которые возможна конкуренция, могут быть различными. Первое, что приходит в голову при поиске этих примеров, - это пища. Действительно, конкуренция за пищу в природе чрезвычайно распространена. Однако предметом конкуренции могут оказываться и пространство, и убежища, и вода, и свет, и даже информационная среда. Так, за свет и пространство нередко конкурируют растения, например, деревья в лесу. Примером конкуренции за информационную среду может послужить взаимодействие поющих видов насекомых в одном биотопе: в случае сходства сигналов разных видов видоспецифические сигналы одних из них могут оказываться помехами для других.
По типу взаимодействия особей между собой различают два типа конкуренции: интерференцию и эксплуатацию. В случае интерференции конкурирующие особи взаимодействуют друг с другом непосредственно, подавляя друг друга и тем самым высвобождая для себя ту часть ресурса, которая использовалась конкурентом. Взаимоотношения особей или популяций принимают при этом характер прямого антагонизма. Например, широко известен факт, что львы при любом удобном случае уничтожают детенышей леопарда и, наоборот, так же леопарды при случае поступают с львятами. При этом такие действия нельзя объяснить просто хищничеством, поскольку львы могут нападать на детенышей леопарда в присутствии его матери, то есть подвергаясь риску вступить с ней в схватку, никак не оправданному при наличии поблизости менее опасных жертв. Кроме того, нередко такие убитые детеныши остаются несъеденными. В пустыне Мохаве (Калифорния и Невада) местный снежный баран и одичавший осел конкурируют за воду и корм. При прямых столкновениях ослы доминируют над баранами: когда ослы приближаются к источникам воды, занятым баранами, последние уступают им место, а иногда и вовсе уходят из данной местности. Другие случаи подобных взаимодействий мы обсуждали, говоря об аллелопатии; очевидно, что при этом взаимодействия двух разных видов грибов или растений, подавляющих рост друг друга с помощью химических веществ, вполне соответствует определению интерференционной конкуренции.
Интерференционная конкуренция может быть и вариантом конкуренции внутривидовой. Например, у самых разных животных, начиная от собак и кончая навозными жуками-скарабеями, можно наблюдать драки из-за пищи. Иногда в этой ситуации бывают столь же беспощадные схватки, как и между особями разных видов. Например, для некоторых наездников, личинки которых паразитируют по одной в особи хозяина, известно такое поведение: отродившаяся из яйца личинка активно разыскивает внутри хозяина других особей своего вида и стремится уничтожить их при помощи челюстей. Следует, однако, заметить, что нередко внутривидовая интерференционная конкуренция отличается от межвидовой по меньшей мере двумя особенностями. Во-первых, предметом конкуренции могут оказываться и партнеры для размножения, то есть особи противоположного пола. Во-вторых, у животных, имеющих более-менее сложные формы поведения, агрессия обычно оказывается в той или иной мере ритуализированной, в этом случае ее целью является не уничтожение конкурента, а его изгнание или перемещение на более низкую ступень в иерархии стаи. Примером обеих своеобразных черт этого варианта взаимодействия между особями могут послужить ритуальные поединки самцов в период размножения, свойственные многим млекопитающим и птицам.
В случае эксплуатационной конкуренции особи взаимодействуют друг с другом не прямо, а косвенно, через общее потребление какого-либо ресурса. В целом, в этом случае успех одного из видов определяется более эффективным использованием ограниченного ресурса, проявляющимся в более быстром наращивании численности, а иногда и просто в более быстром использовании ресурса, которого просто не остается в количестве, достаточном для конкурента. Классическим примером такой конкуренции является продемонстрированное в лабораторных опытах Г.Ф. Гаузе взаимодействие двух разных видов инфузорий-туфелек – Paramecium caudatum и Paramecium aurelia – в среде с ограниченным запасом пищи. Эти опыты мы рассмотрим сегодня подробнее, говоря о динамике численности популяций в системе из двух конкурентов. Случаи эксплуатационной конкуренции нередки и в природных условиях, хотя то, что она не сопровождается при этом и элементами конкуренции интерференционной, доказать можно далеко не всегда. Тем не менее, именно к эксплуатационной конкуренции, по-видимому, относятся многочисленные примеры вытеснения из исконных областей обитания видов более поздними вселенцами, не проявляющими прямой агрессии по отношению к первым. Например, на территории России за XX век узкопалый речной рак сильно расширил на север границу своего ареала и во многих местах вытеснил другой вид – широкопалого речного рака. Прямая агрессия узкопалого рака в отношении широкопалого и по частоте, и по успешности сопоставима с внутривидовой агрессией у обоих видов. Тем не менее, при появлении в водоеме, заселенном широкопалым раком, узкопалый вытесняет его за несколько лет. Предполагается, что в основе этого явления лежит меньшая требовательность узкопалого речного рака к чистоте воды и большая устойчивость к некоторым инфекциям. Другим примером может послужить вытеснение в нашем городе черного таракана рыжим (прусаком): за последние несколько десятилетий черный таракан, в начале XX века широко распространенный в петербургских домах, стал большой редкостью. Прямой агрессии по отношению друг к другу эти виды, в целом, не проявляют, однако черный таракан медленнее развивается и проявляет меньшую заботу о потомстве, а именно: самки прусака носят свои оотеки (капсулы с яйцами) практически до выхода личинок, в то время как черные тараканы быстро бросают их на произвол судьбы.
Приведенные примеры тем, что в описанных случаях один вид неизменно вытеснял другой, демонстрируют одну важную черту конкурентных взаимодействий – их асимметрию. Это означает, что степень отрицательного воздействия одной популяции или особи на другую нередко отличается в ту или иную сторону от величины ответного воздействия. Считается, что асимметричная конкуренция – явление, значительно более распространенное, чем симметричная. Как уже говорилось, крайним случаем этой асимметрии является аллелопатия, когда один конкурент настолько сильно и эффективно подавляет другого, что не встречает ответного сопротивления.
Рассмотрим, какие закономерности должна проявлять численность особей в конкурирующих популяциях. Одной из наиболее распространенных моделей является модель Лотки-Вольтерра, видоизмененная Гаузе. В отличие от уравнения Лотки-Вольтерра, разработанного для системы хищник – жертва, данная модель исходит из предположения о логистическом росте популяций и включает в себя больше параметров. Как в случаях аналитических моделей обычно и бывает, мы рассматриваем замкнутую систему, то есть численность популяций ограничивается только взаимодействием особей, входящих в состав этих двух популяций, а никакие прочие факторы в расчет не принимаются. Уравнения, описывающие численность обеих популяций, практически идентичны:
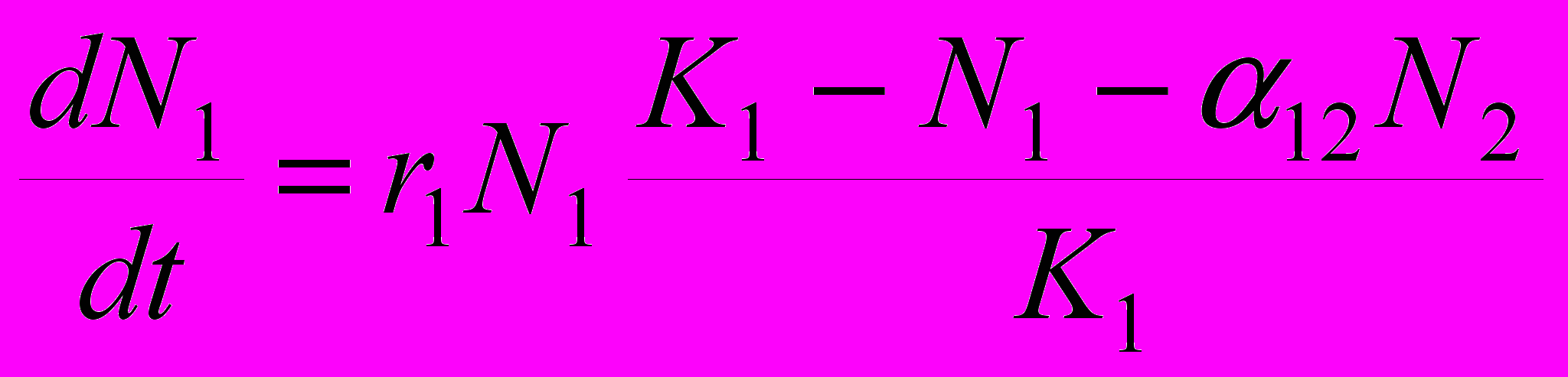
и
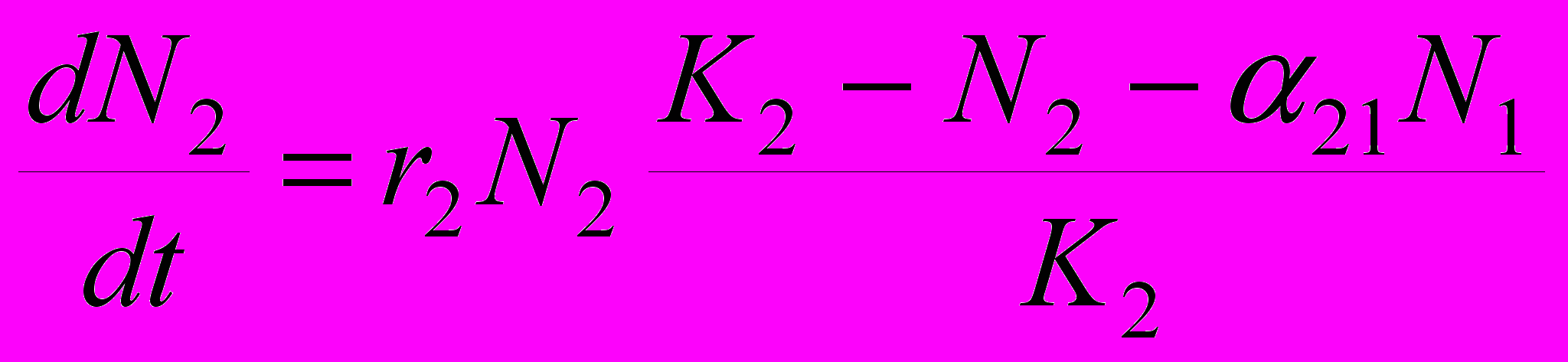 ,
,где N1 и N2 – численность соответственно 1-й и 2-й популяции,
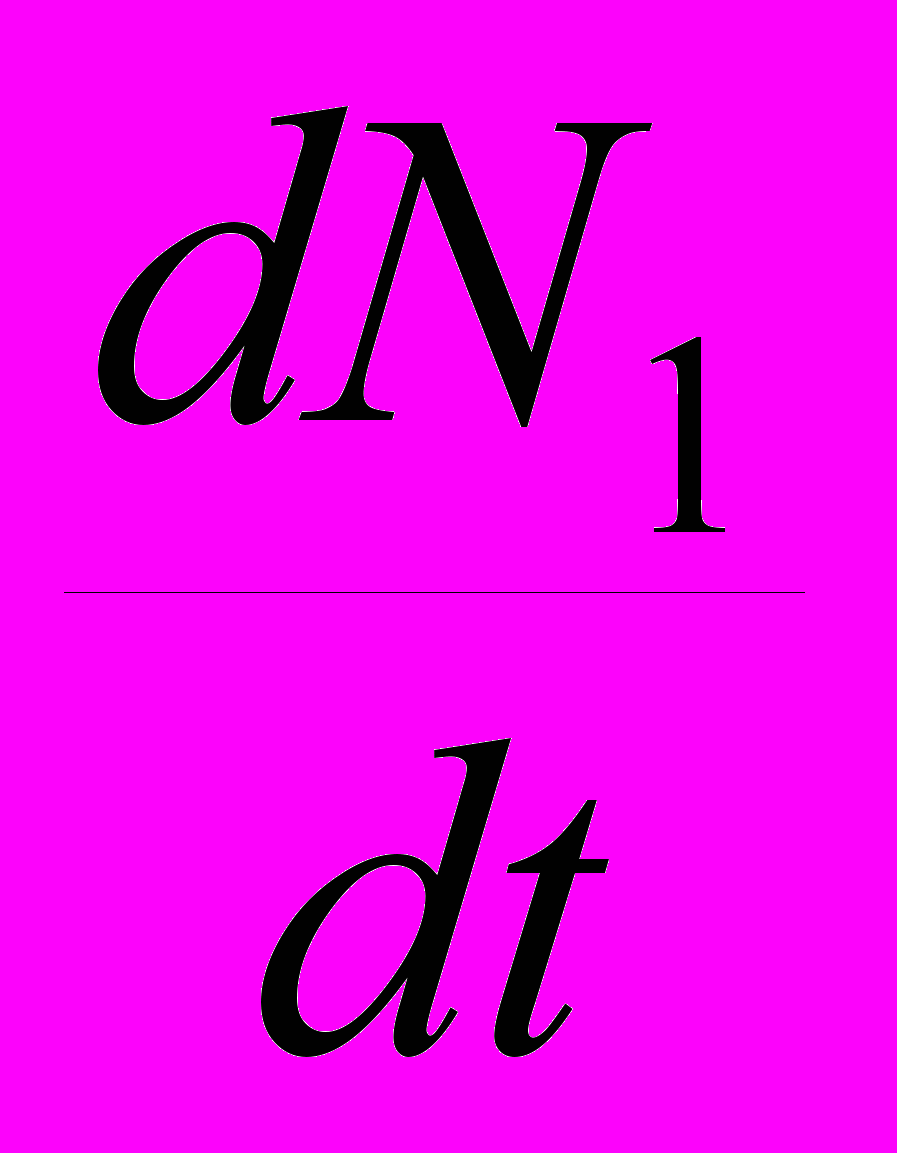 и
и 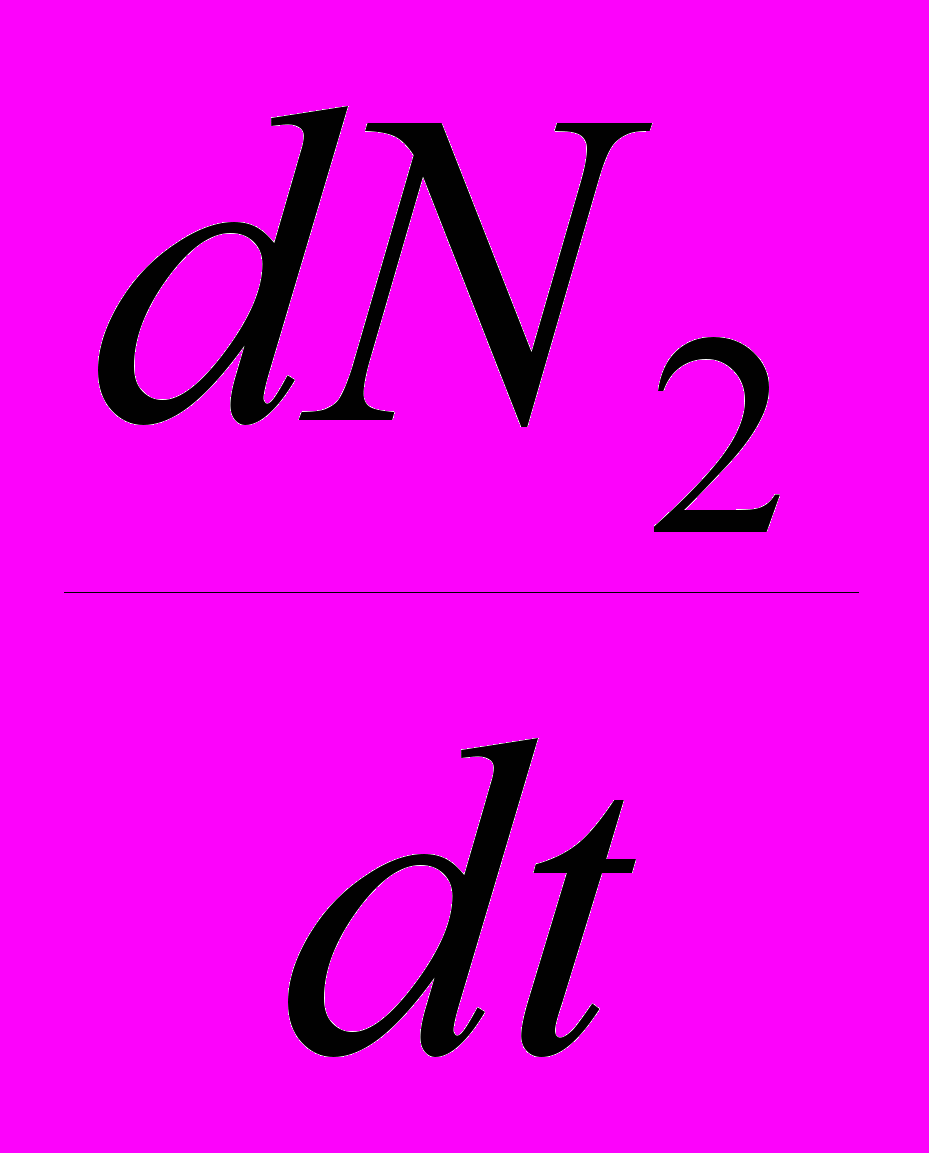 – их скорости приращения численности, r1 и r2 – их потенциальные удельные скорости роста (то есть константы экспонент для данных популяций в случае их неограниченного роста), K1 и K2 – их верхние пределы численности. Наконец, имеются два коэффициента , отражающие влияние одного вида на другой: 12 – степень влияния 2-го вида на 1-й, а 21 – степень влияния 1-го вида на 2-й. Поясню смысл такого коэффициента . Степень влияния – это снижение численности вида, подверженного этому влиянию. Теперь о том, что означает величина коэффициента . Будем называть тот вид, угнетающее влияние на который конкурента нас интересует, «нашим видом». Тогда величина показывает, сколько особей вида-конкурента угнетающе влияет на популяцию нашего вида в той же мере, как одна особь нашего вида. Например, если 12 = 0.5, то это означает: для того, чтобы угнетающее влияние вида 2 на популяцию вида 1 было таким же, как действие на нее же одной особи вида 1, любую особь вида 1 можно заменить половиной особи вида 2. Иными словами, одна особь вида 2 действует на одну особь вида 1 так же, как на особь вида 1 действует 2 особи вида 1.
– их скорости приращения численности, r1 и r2 – их потенциальные удельные скорости роста (то есть константы экспонент для данных популяций в случае их неограниченного роста), K1 и K2 – их верхние пределы численности. Наконец, имеются два коэффициента , отражающие влияние одного вида на другой: 12 – степень влияния 2-го вида на 1-й, а 21 – степень влияния 1-го вида на 2-й. Поясню смысл такого коэффициента . Степень влияния – это снижение численности вида, подверженного этому влиянию. Теперь о том, что означает величина коэффициента . Будем называть тот вид, угнетающее влияние на который конкурента нас интересует, «нашим видом». Тогда величина показывает, сколько особей вида-конкурента угнетающе влияет на популяцию нашего вида в той же мере, как одна особь нашего вида. Например, если 12 = 0.5, то это означает: для того, чтобы угнетающее влияние вида 2 на популяцию вида 1 было таким же, как действие на нее же одной особи вида 1, любую особь вида 1 можно заменить половиной особи вида 2. Иными словами, одна особь вида 2 действует на одну особь вида 1 так же, как на особь вида 1 действует 2 особи вида 1.Если одна особь вида-конкурента по угнетающему влиянию соответствует меньше, чем одной особи нашего вида, это означает, что межвидовая конкуренция слабее, чем внутривидовая у нашего вида. В численном выражении это будет выглядеть так: < 1. Наоборот, если одна особь вида-конкурента по угнетающему влиянию соответствует нескольким особям нашего вида, то это означает, что межвидовая конкуренция сильнее, чем внутривидовая. В численном выражении это будет выглядеть так: > 1.
Как всегда, попробуем выяснить, при каких условиях численность популяции при данных условиях будет постоянной, то есть ее прирост – нулевым.
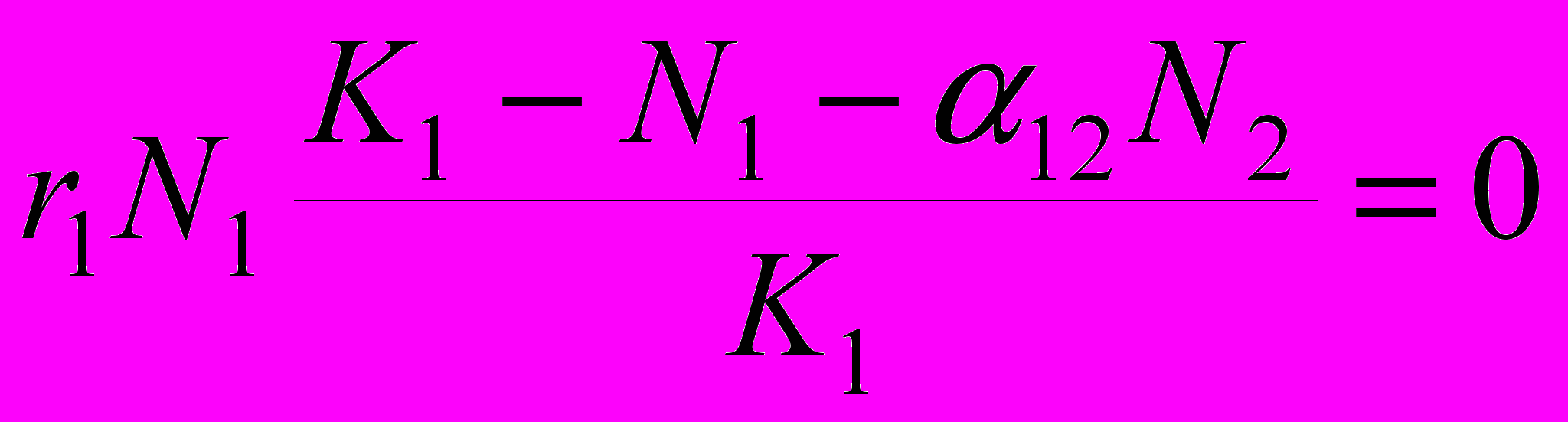
Поскольку величины r, N и K по смыслу модели не могут равны нулю, равенство нулю должно означать, что K1 – N1 – 12N2 = 0, или N1 = K1 – 12N2. Нетрудно заметить, что последнее выражение – ни что иное, как уравнение прямой линии. Смысл его состоит в том, что при постоянстве максимально возможной численности «нашей» популяции (K1) и влияния на численность «нашей» популяции вида-конкурента выполняется следующая закономерность: чем ниже численность нашего вида, тем выше нужна численность конкурента для того, чтобы поддерживать данный постоянный уровень численности этой нашей популяции.
Это же выражение можно представить и как зависимость численности вида-конкурента от численности нашего вида: 12N2 = K1 – N1, или
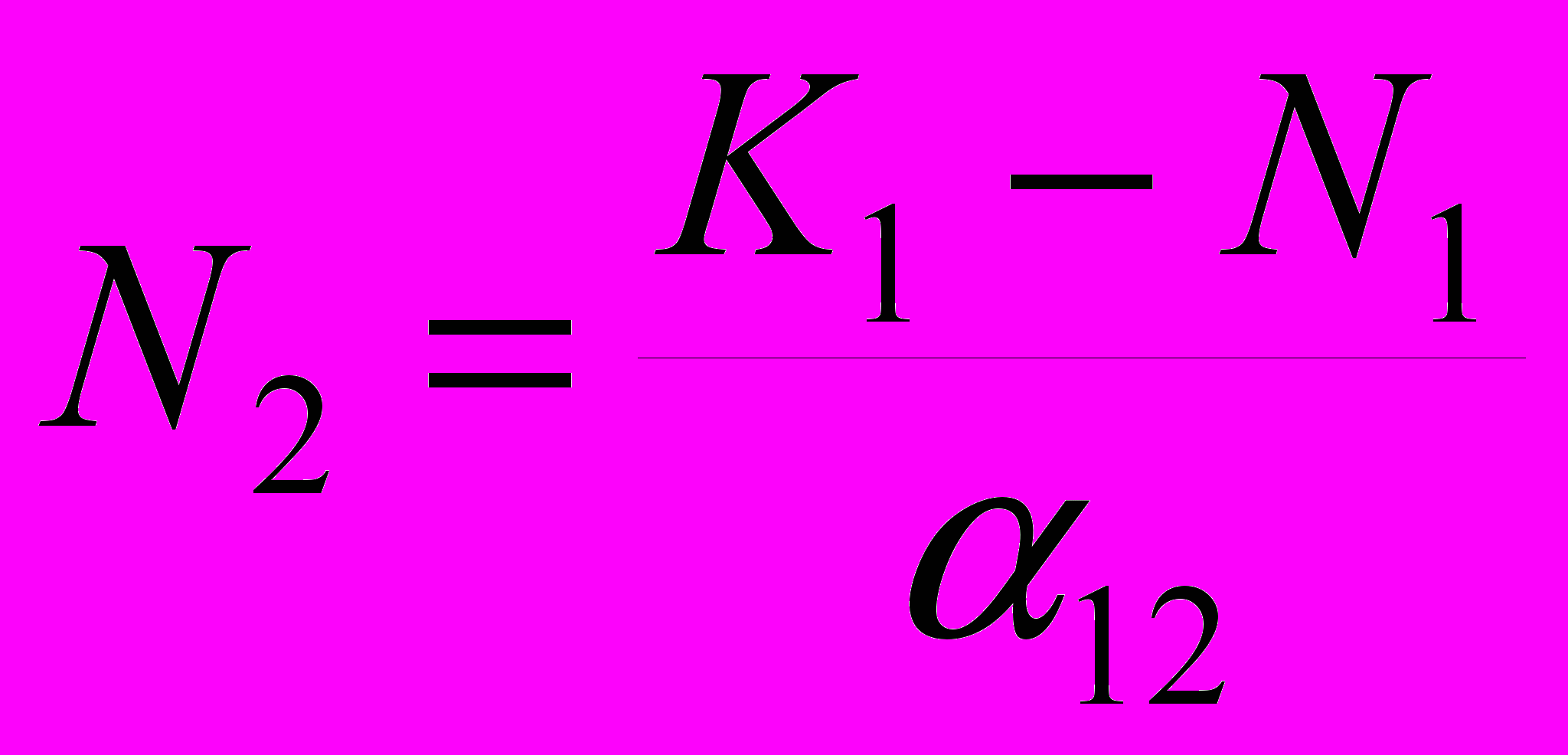 , или
, или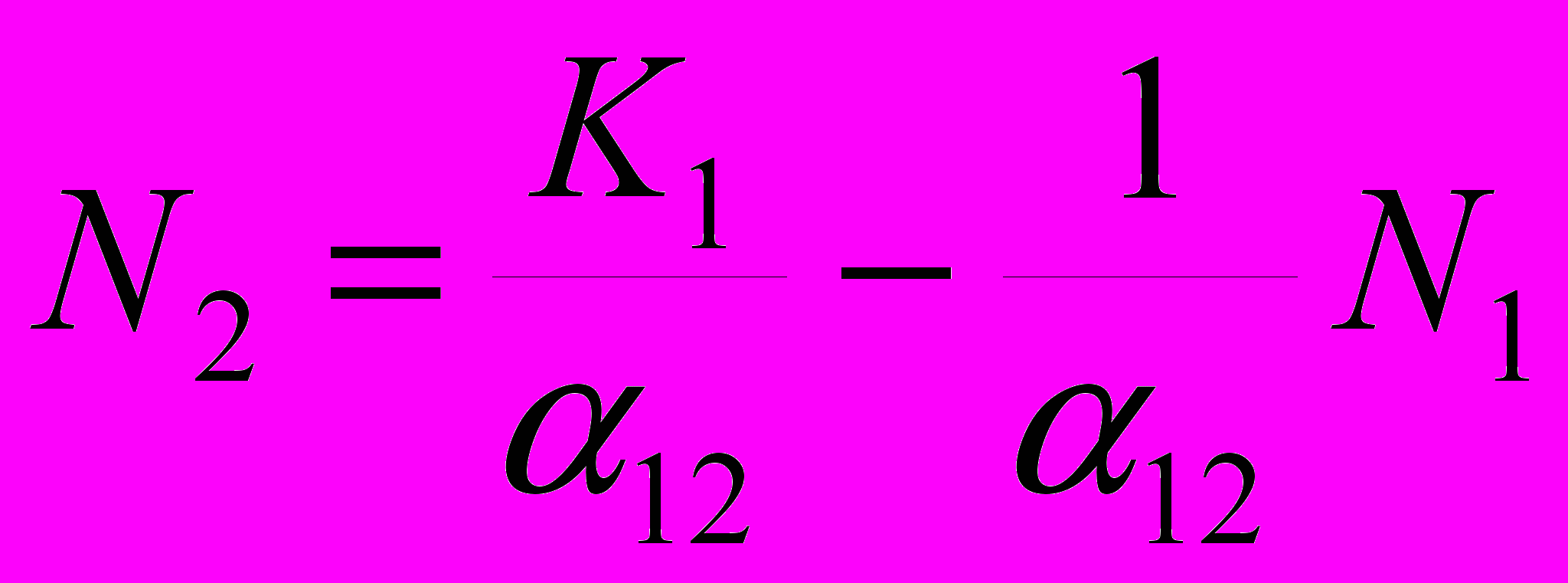
Аналогично, нетрудно вывести и формулы, связывающие численности двух конкурирующих популяций с предельной численностью 2-й популяции и влиянием нашего вида на вид-конкурент:
N2 = K2 – 21N1 и
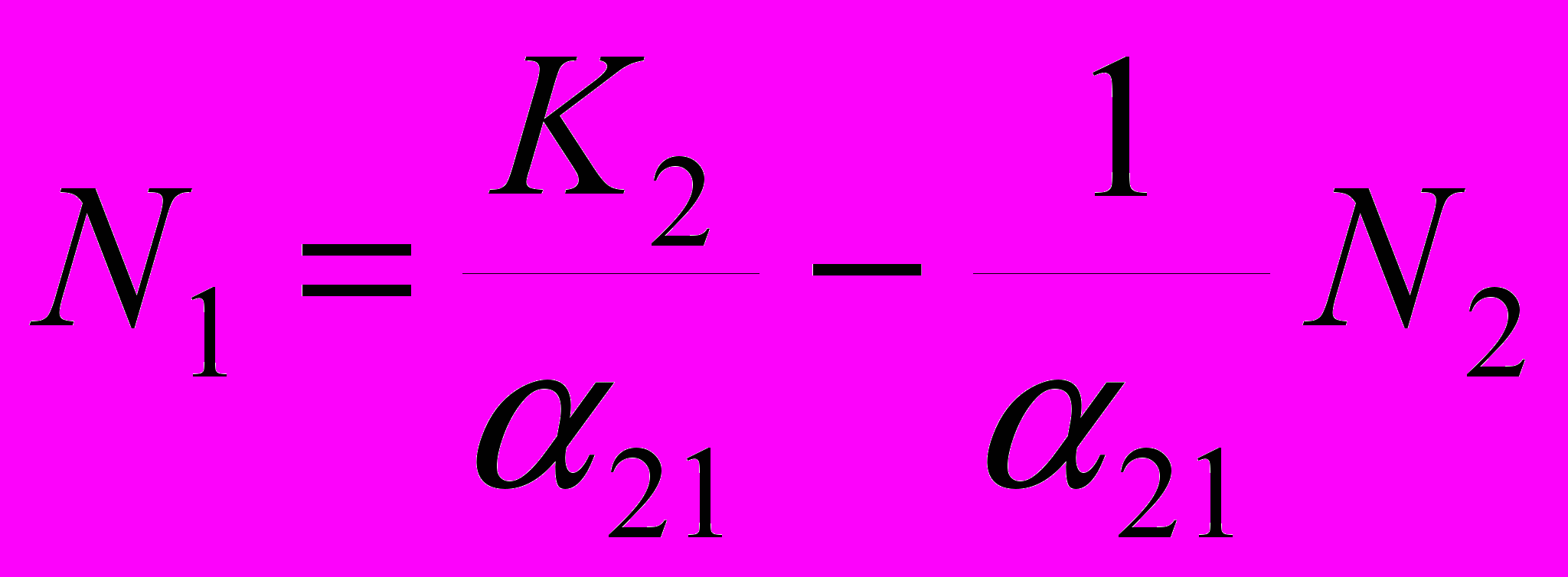
Таким образом, мы можем нанести на одну и ту же координатную плоскость две прямые, связывающие численности двух конкурирующих популяций. Одна линия будет отображать, как должны соотноситься численности двух популяций для того, чтобы на одном уровне поддерживалась численность популяции 1 («нашей»), вторая – как должны соотноситься численности двух популяций для того, чтобы на одном уровне поддерживалась численность популяции 2 («конкурента»).
Например, по оси абсцисс мы будем отсчитывать численность популяции 1 («нашей»), а по оси ординат – численность популяции 2 («конкурента»). Тогда первая линия (описывающая условие стабильности численности «нашего вида») пересечет ось ординат в точке со значением
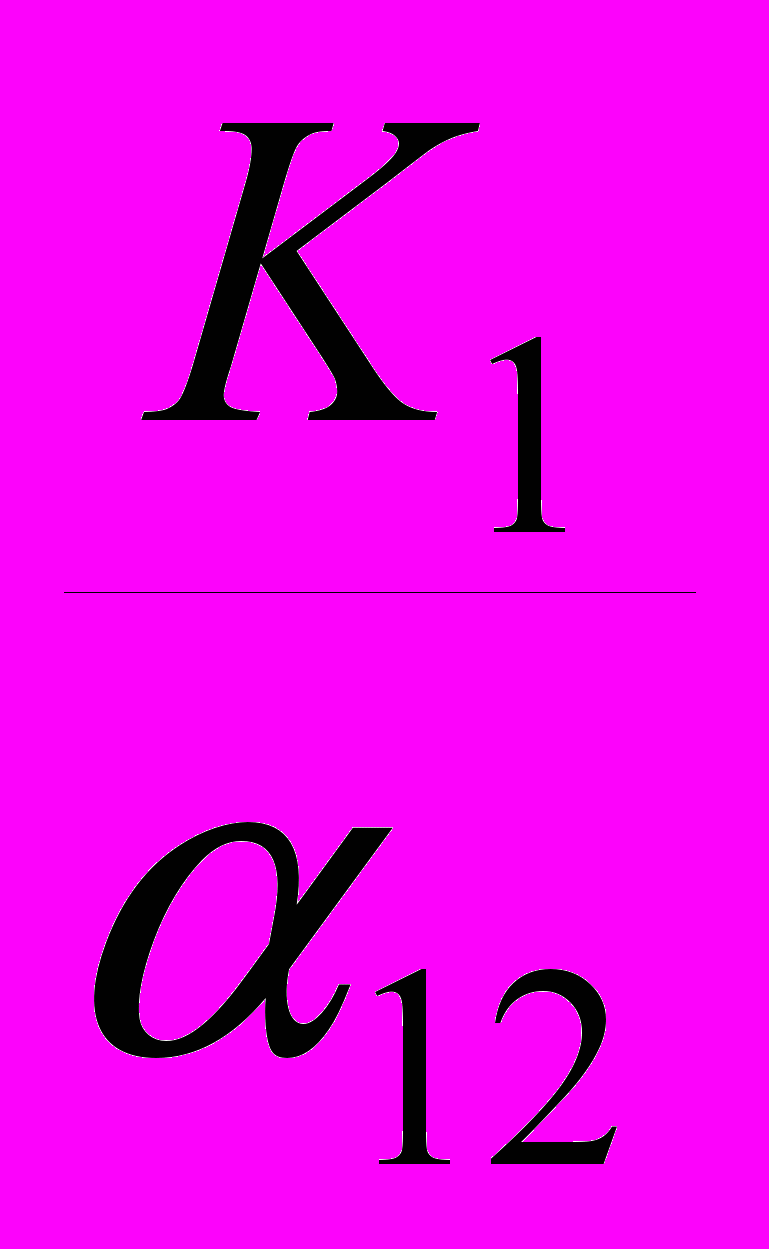 , а ось абсцисс – в точке со значением K1. Вторая линия (описывающая условие стабильности численности «вида-конкурента») пересечет ось ординат в точке со значением K2, а ось абсцисс – в точке со значением
, а ось абсцисс – в точке со значением K1. Вторая линия (описывающая условие стабильности численности «вида-конкурента») пересечет ось ординат в точке со значением K2, а ось абсцисс – в точке со значением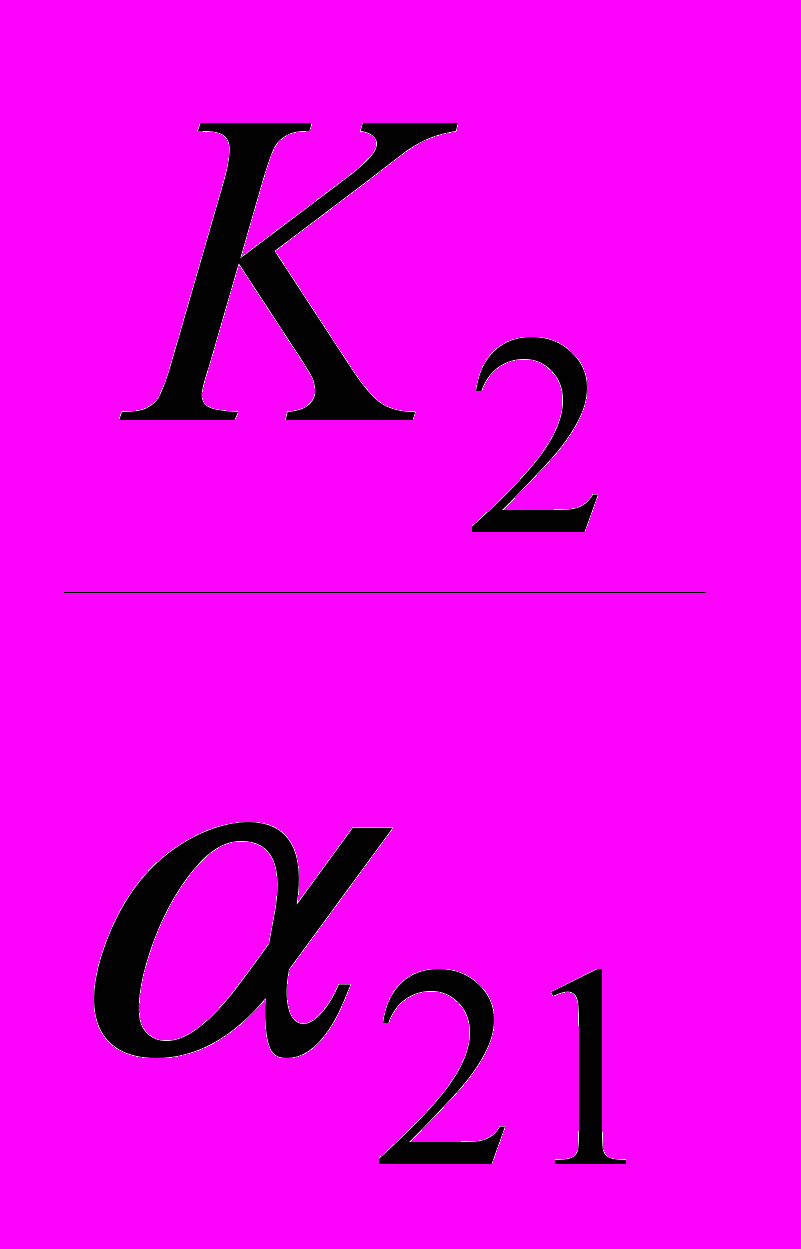 .
. Если численность особей в любой из популяций оказывается ниже равновесной (при сохранении равновесной численности у 2-й популяции), она будет стремиться нарастать, пока не вернется к точке равновесия. Наоборот, если она окажется выше равновесной (также при сохранении равновесной численности у 2-й популяции), она будет снижаться. На иллюстрациях эти направления возврата численности к равновесной показаны горизонтальными стрелками для N1 и вертикальными для N2.
В зависимости от того, как соотносятся эти 4 величины, мы сможем построить целых 4 варианта взаиморасположения этих линий, каждый из которых будет означать свой прогноз последствий данных конкурентных сотношений.
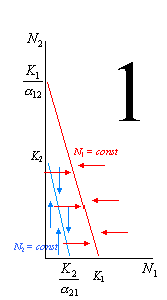
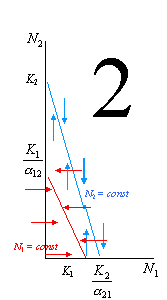
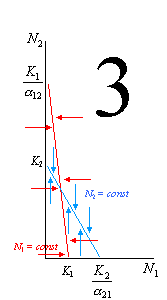
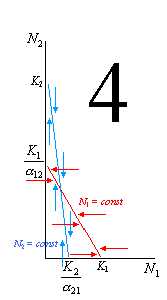
Рассмотрим эти 4 варианта более подробно. На рисунке 1 выполняются следующие условия: K2 / 12 < K1 (то есть вид 1 может сдерживать вид 2) и K1 / 21 > K2 (то есть вид 2 не может сдерживать вид 1). В этой ситуации первый вид может наращивать численность и тогда, когда численность 2-го вида достигла максимума (K2), его влияние на конкурента становится всё сильнее. В итоге вид 1 неминуемо вытесняет своего конкурента. 2-й случай – строго противоположный: K2 / 12 > K1 (то есть вид 1 не может сдерживать вид 2) и K1 / 21 < K2 (то есть вид 2 может сдерживать вид 1). В этой ситуации 1-й вид сам будет вытеснен. Случай 3 отражает ситуацию неустойчивого равновесия, когда оба вида не могут сдерживать численность друг друга (K2 / 12 > K1 и K1 / 21 > K2). При определенных значениях численностей (соответствуют точке пересечения двух прямых) обоих конкурентов эти численности могут стабилизироваться, но после малейшего увеличения ставший более многочисленным вид неминуемо вытеснит конкурента. Наконец, в случае 4 (K2 / 12 < K1 и K1 / 21 < K2) ни один вид не может достичь такой численности, которая соответствовала бы полному отсутствию вида-конкурента, – это устойчивое равновесие.
Г. Ф. Гаузе не только предложил эту переработанную для конкуренции модель Лотки – Вольтерра, он попытался проверить ее экспериментальным путем. Как и в случае проверки аналитической модели хищник – жертва, экспериментальная модель Гаузе представляла собой систему из двух видов инфузорий. Однако теперь уже были взяты два вида инфузории-туфельки, Paramecium caudatum и Paramecium aurelia. Оба вида сосуществовали в одном сосуде и кормились бактериями или дрожжами, которые периодически помещались в среду в ограниченном количестве. Сначала, до определенного предела, численность обоих видов росла по кривой, близкой к логистической, затем у одного из видов начиналось снижение численности, которое заканчивалось его полным исчезновением. Характерной чертой этого явления было то, что при одних и тех же условиях опыта побеждал неизбежно один и тот же вид инфузорий, однако изменение условий опыта могло приводить к смене вида-победителя. Так, при кормлении бактериями Pseudomonas aeruginosa всегда побеждала инфузория P. aurelia, а при кормлении дрожжами Saccharomyces exiguus неизменно побеждала инфузория P. caudatum. Эта разница связана с тем, что P. caudatum плохо переносит продукты метаболизма, выделяемые в среду бактериями.
На основании экспериментальных данных Гаузе сформулировал принцип конкурентного исключения (известный ныне также как принцип Гаузе), согласно которому два вида с совпадающими экологическими нишами сосуществовать не могут. Этот принцип иллюстрируется тремя из четырех приведенных только что графиков. По-видимому, ситуация, отраженная на графике 4, в природе реализуется, действительно, не часто. Примеры вытеснения одного вида другим, приведенные в начале лекции, хорошо согласуются с принципом Гаузе. Однако, очевидно, что достаточно сложно найти, а тем более доказать, примеры полного совпадения экологических ниш у разных видов. Как правило, более-менее детальное изучение конкурирующих видов, даже очень сходных, демонстрирует их различия с точки зрения чувствительности к значениям тех или иных экологических факторов. Это обстоятельство можно истолковать и как трудность доказательства принципа Гаузе на биологических примерах, однако можно подойти к нему и с другой точки зрения. А именно, оно может объяснять механизмы, с помощью которых популяции разных видов, встречающихся в одной экосистеме, могут избегать конкуренции.
Впоследствии мы более детально разберем понятие «экологическая ниша». Сейчас же я объясню механизм избегания конкуренции в самом общем виде. Предположим, у нас имеются популяции двух видов, сходных по своим экологическим требованиям. Если мы представим себе экологические ниши этих двух видов в виде «гиперколоколов», эти области в пространстве экологических факторов в значительной мере пересекутся. Однако почти наверняка останутся области значений экологических факторов, при которых один вид существовать может, а другой – нет (и наоборот) и примыкающие к этим областям зоны (возможно, значительные), при которых этот вид размножается успешнее, чем его конкурент. Если в условиях эксперимента мы лишаем исследуемые организмы выбрать территорию своего обитания, в природе они обычно имеют этот выбор и определенным образом перераспределяются. При этом стóит заметить, что такое перераспределение (между территориями, между стациями и т. п.) обеспечивается не только и не столько целенаправленными миграциями, а большей вероятностью выживания особей того или иного вида на этих разных территориях или в разных стациях. Ну, а коль скоро такое начальное размежевание происходит, далее могут вступать в действие механизмы естественного отбора, в результате которых непересекающиеся области экологических ниш становятся для этих популяций основными и даже смещаются по принципу «взаимоотталкивания».
Феноменология динамики численности
Если говорить о том, какие формы принимает динамика численности живых организмов в реальной природе, а не в моделях, мы столкнемся с определенными трудностями. Если продолжительность жизни отдельной особи очень велика, например, составляет сотни лет, как у некоторых древесных растений, мы не имеем возможности проследить непосредственно смену поколений от начала до конца, однако, по крайней мере, можем с бóльшим или меньшим успехом реконструировать динамику их численности, изучая возрастную структуру популяции на текущий момент. Если продолжительность жизни особи недостаточно велика для того, чтобы в популяции одновременно сосуществовало много поколений, однако непосредственный учет изменения численности затруднен, единственным средством восстановления картины являются статистические данные за много лет и лишь иногда – какие-либо косвенные данные, например – накапливающиеся раковины моллюсков, оболочки диатомовых водорослей и другие поддающиеся датировке следы жизнедеятельности. Наконец, в ряде случаев смена поколений происходит достаточно быстро и при этом численность организмов легко поддается учету, это наиболее удобный для исследования случай. Именно такая ситуация характерна для многих насекомых, при этом исследование и прогнозирование динамики их численности часто оказывается важным в хозяйственном отношении. Рассмотрим этот пример более подробно.
Всё разнообразие динамики численности насекомых, в общих чертах, можно свести к трем основным типам – устойчивому, сезонному и однолетнему.
Устойчивый тип динамики численности означает более-менее постоянную численность в течение всего вегетационного периода. Он характерен для видов, отличающихся высокой выживаемостью при относительно низкой плодовитости (K-стратегов).
Сезонный тип динамики отличается тем, что численность вида значительно возрастает в течение сезона, однако сильно снижается к началу следующего сезона за счет малого количества успешно перезимовывающих особей. Обычно увеличение численности в течение сезона происходит в процессе смены поколений (виды, дающие несколько поколений за сезон, называют поливольтинными, а одно – моновольтинными). В случае моновольтинных видов, такой эффект может достигаться за счет недружного развития и неодновременного созревания. Высокие плодовитость и смертность, характерные для поливольтинных видов, имеющих сезонный тип динамики, свидетельствуют об их следовании r-стратегии.
Наиболее сложный тип динамики численности насекомых – многолетний. Изменение численности и плотности особей в популяции и заселенности стаций охватывает период в несколько лет и может быть подразделено на несколько (4 - 5) фаз, периодически сменяющих одна другую. В наиболее простом варианте эти фазы следующие.
- Фаза минимума, или депрессии – численность минимальна;
- фаза подъема, или нарастания (продромальная) – период наращивания популяцией численности, плотности и количества заселенных стаций;
- фаза максимума, или массовой вспышки, (эруптивная) – численность и плотность популяции, количества занятых стаций достигли максимума, в этот период насекомые-вредители наносят максимальный ущерб сельскому и лесному хозяйству;
- фаза спада, или кризиса, - численность начинает снижаться, иногда очень быстро, под влиянием внешних факторов, в первую очередь биотических (недостаток пищи, хищники, паразиты).
Многолетний тип динамики численности встречается и у моновольтинных, и у поливольтинных видов, от числа поколений в году отчасти зависит продолжительность полного цикла фаз, который может составлять от 2-3 до 6-10 и более лет. Стабильность периода и разница между численностью в фазу депрессии и в фазу максимума очень различны у разных видов. Считается, что стабильность периода и выравненность численности в популяции насекомого-фитофага свидетельствуют о большей сбалансированности в отношениях между этой популяцией и популяцией кормового растения. Так, считается, что наблюдаемые в сибирской тайге нерегулярные вспышки массового размножения сибирского шелкопряда, монашенки и других хвоегрызущих чешуекрылых, носящие порой катастрофический характер (полностью оголяются огромные массивы леса, после чего на ослабленные деревья нападают стволовые вредители), отражают относительную молодость этого типа экосистем, что подтверждается и палеонтологическими и геологическими данными. В случае насекомых-энтомофагов изменения численности в значительной мере связаны еще и с колебаниями численности вида-жертвы (хозяина), которая в свою очередь зависит от колебаний численности его кормового организма.