
Общая характеристика работы актуальность
| Вид материала | Автореферат |
- Общая характеристика работы актуальность работы, 227.87kb.
- Общая характеристика работы актуальность работы, 227.87kb.
- Общая характеристика работы актуальность, 242.28kb.
- Общая характеристика работы актуальность работы, 336.09kb.
- Общая характеристика работы актуальность работы, 236.99kb.
- I. общая характеристика работы актуальность темы исследования, 263.23kb.
- Общая характеристика работы актуальность работы, 487.01kb.
- Общая характеристика работы. Актуальность работы, 1103.53kb.
- Общая характеристика работы актуальность проблемы, 254.34kb.
- Общая характеристика работы актуальность темы исследования, 305.28kb.
Глава 4. ЭКОЛОГО-ФАУНИСТИЧЕСКИЙ АНАЛИЗ ПАРАЗИТОВ ГОЛЬЦОВ ПАЛЕАРКТИКИ И НЕАРКТИКИ
4.1. Фауна паразитов и экология гольцов арктической группы. У арктического гольца отмечен 91 вид паразитов, выявлены различия в заражённости, связанные с разным характером питания этих рыб в водоёмах Кольского полуо-ва (рис. 2).
У палии в озёрах Карелии найдено 28 видов паразитов (Рыбак, 1985; Румянцев, 1996; Мамонтова и др., 2005), ведущую роль в её питании играют реликтовые амфиподы, рыбы (ряпушка и корюшка) и хирономиды-ортокладиины, что определяет особенности рациона и инвазии гольцов паразитами.
палии в озёрах Карелии найдено 28 видов паразитов (Рыбак, 1985; Румянцев, 1996; Мамонтова и др., 2005), ведущую роль в её питании играют реликтовые амфиподы, рыбы (ряпушка и корюшка) и хирономиды-ортокладиины, что определяет особенности рациона и инвазии гольцов паразитами.
Б
Рис. 2. Дендрограмма различий популяций арктического гольца Кольского полуострова по фауне паразитов, шкала - мера расхождения, построение методом Уорда
оганидская палия бассейна р. Хатанга и оз. Эльгыгытгын на Чукотке заметно различаются по фауне паразитов (нет общих видов) и типу питания (эврифаг и специализированный хищник соответственно). У арктического гольца из озёр Забайкалья найдено
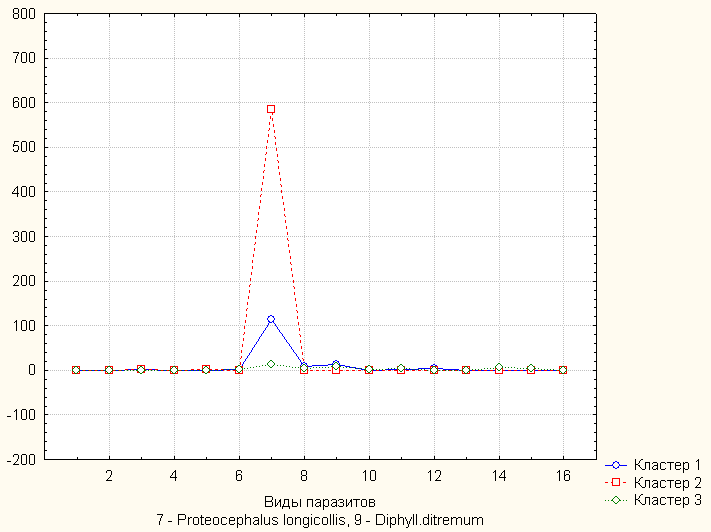
19 видов паразитов, выявлены достоверные различия между популяциями из разных озёр по интенсивности инвазии цестодами Proteocephalus longicollis и Diphyllobothrium ditremum (рис. 3), что обусловлено разным питанием рыб и типом водоёмов (с преобладанием рыб равнинного или предгорного комплексов). В оз. Большое Леприндо отмечена тенденция к расхождению одной формы гольцов по питанию (рис. 4), выявлены очаги опасных для рыб гельминтозов. Фауна паразитов эндемичных гольцов Сибири - гольцов Черского, якутского, таймырского очень сходна (рис. 5) и близка к таковой арктического гольца Забайкалья и гольца Дрягина, по характеру питания это эврифаги с переходом к хищничеству и каннибализму (гольцы якутский, Черского) в связи с ограниченной кормовой базой и экстремальными условиями выживания в тундровых озёрах Заполярья.
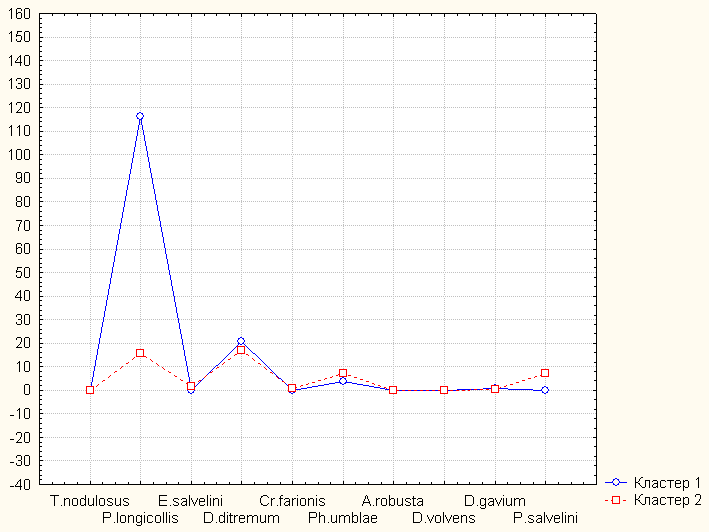
Анализ фауны паразитов гольца Таранца из озёр Чукотки (Атрашкевич и др., 1991, 1993; Шедько и др., 2004), Аляски (Moles, 1982) и Канады (Margolis, Arthur, 1979; McDonald, Margolis, 1995) свидетельствует о наличии озёрного и проходного экотипов, доминировании пресноводных паразитов (72,1%) и широком спектре питания. Проведённое нами паразитологическое изучение гольца Крогиус оз. Дальнее
п
Рис. 3. Различия между популяциями гольцов озёр Забайкалья по заражённости гельминтами, шкала - виды паразитов, метод К-средних
 озволило уточнить его рацион: показало важную роль амфипод, наряду с молодью нерки, амфибиотическими насекомыми и др., отмечено 30 видов паразитов. Первые данные о паразитах Salvelinus sp. оз. Начикинское (табл. 4) свидетельствуют о его эврифагии - питании как рыбой, так и разнообразными беспозвоночными, отмечено значительное обеднение фауны его паразитов в высокогорном озере и экологическое сходство этого гольца с нейвой. Нейва - озёрный эврифаг, близкий к другим арктическим гольцам (рис. 7), с дифференцированным питанием молоди, карликов и половозрелых рыб. Большая или меньшая степень выраженности ихтио- и бентофагии у разных видов гольцов зависит от разнообразия гидроихтиоценоза. Наиболее разнообразна фауна паразитов гольцов, имеющих проходной и туводный экотипы (гольцы Таранца, арктический), (43-91 вид), у озёрных гольцов (нейва, голец Крогиус, даватчан, начикинский и др.) она представлена меньшим числом видов (12-30 видов), наиболее обеднена паразитофауна ледниковых реликтов и эндемиков - палий крупных (Ладожское, Онежское) и мелких тундровых озер Заполярья и якутского гольца (5-20 видов). Преобладание морских паразитов у гольца Леванидова (12 видов паразитов) свидетельствует
озволило уточнить его рацион: показало важную роль амфипод, наряду с молодью нерки, амфибиотическими насекомыми и др., отмечено 30 видов паразитов. Первые данные о паразитах Salvelinus sp. оз. Начикинское (табл. 4) свидетельствуют о его эврифагии - питании как рыбой, так и разнообразными беспозвоночными, отмечено значительное обеднение фауны его паразитов в высокогорном озере и экологическое сходство этого гольца с нейвой. Нейва - озёрный эврифаг, близкий к другим арктическим гольцам (рис. 7), с дифференцированным питанием молоди, карликов и половозрелых рыб. Большая или меньшая степень выраженности ихтио- и бентофагии у разных видов гольцов зависит от разнообразия гидроихтиоценоза. Наиболее разнообразна фауна паразитов гольцов, имеющих проходной и туводный экотипы (гольцы Таранца, арктический), (43-91 вид), у озёрных гольцов (нейва, голец Крогиус, даватчан, начикинский и др.) она представлена меньшим числом видов (12-30 видов), наиболее обеднена паразитофауна ледниковых реликтов и эндемиков - палий крупных (Ладожское, Онежское) и мелких тундровых озер Заполярья и якутского гольца (5-20 видов). Преобладание морских паразитов у гольца Леванидова (12 видов паразитов) свидетельствует
4.1. Фауна паразитов и экология гольцов арктической группы. У арктического гольца отмечен 91 вид паразитов, выявлены различия в заражённости, связанные с разным характером питания этих рыб в водоёмах Кольского полуо-ва (рис. 2).
У
палии в озёрах Карелии найдено 28 видов паразитов (Рыбак, 1985; Румянцев, 1996; Мамонтова и др., 2005), ведущую роль в её питании играют реликтовые амфиподы, рыбы (ряпушка и корюшка) и хирономиды-ортокладиины, что определяет особенности рациона и инвазии гольцов паразитами.Б
Рис. 2. Дендрограмма различий популяций арктического гольца Кольского полуострова по фауне паразитов, шкала - мера расхождения, построение методом Уорда
оганидская палия бассейна р. Хатанга и оз. Эльгыгытгын на Чукотке заметно различаются по фауне паразитов (нет общих видов) и типу питания (эврифаг и специализированный хищник соответственно). У арктического гольца из озёр Забайкалья найдено
19 видов паразитов, выявлены достоверные различия между популяциями из разных озёр по интенсивности инвазии цестодами Proteocephalus longicollis и Diphyllobothrium ditremum (рис. 3), что обусловлено разным питанием рыб и типом водоёмов (с преобладанием рыб равнинного или предгорного комплексов). В оз. Большое Леприндо отмечена тенденция к расхождению одной формы гольцов по питанию (рис. 4), выявлены очаги опасных для рыб гельминтозов. Фауна паразитов эндемичных гольцов Сибири - гольцов Черского, якутского, таймырского очень сходна (рис. 5) и близка к таковой арктического гольца Забайкалья и гольца Дрягина, по характеру питания это эврифаги с переходом к хищничеству и каннибализму (гольцы якутский, Черского) в связи с ограниченной кормовой базой и экстремальными условиями выживания в тундровых озёрах Заполярья.
Анализ фауны паразитов гольца Таранца из озёр Чукотки (Атрашкевич и др., 1991, 1993; Шедько и др., 2004), Аляски (Moles, 1982) и Канады (Margolis, Arthur, 1979; McDonald, Margolis, 1995) свидетельствует о наличии озёрного и проходного экотипов, доминировании пресноводных паразитов (72,1%) и широком спектре питания. Проведённое нами паразитологическое изучение гольца Крогиус оз. Дальнее
п
Рис. 3. Различия между популяциями гольцов озёр Забайкалья по заражённости гельминтами, шкала - виды паразитов, метод К-средних
озволило уточнить его рацион: показало важную роль амфипод, наряду с молодью нерки, амфибиотическими насекомыми и др., отмечено 30 видов паразитов. Первые данные о паразитах Salvelinus sp. оз. Начикинское (табл. 4) свидетельствуют о его эврифагии - питании как рыбой, так и разнообразными беспозвоночными, отмечено значительное обеднение фауны его паразитов в высокогорном озере и экологическое сходство этого гольца с нейвой. Нейва - озёрный эврифаг, близкий к другим арктическим гольцам (рис. 7), с дифференцированным питанием молоди, карликов и половозрелых рыб. Большая или меньшая степень выраженности ихтио- и бентофагии у разных видов гольцов зависит от разнообразия гидроихтиоценоза. Наиболее разнообразна фауна паразитов гольцов, имеющих проходной и туводный экотипы (гольцы Таранца, арктический), (43-91 вид), у озёрных гольцов (нейва, голец Крогиус, даватчан, начикинский и др.) она представлена меньшим числом видов (12-30 видов), наиболее обеднена паразитофауна ледниковых реликтов и эндемиков - палий крупных (Ладожское, Онежское) и мелких тундровых озер Заполярья и якутского гольца (5-20 видов). Преобладание морских паразитов у гольца Леванидова (12 видов паразитов) свидетельствует Рис. 4. Различия в заражённости между 2 группами арктических гольцов оз. Большое Леприндо, метод К-средних

Рис. 5. Дендрограмма различий арктических гольцов из разных частей ареала по фауне паразитов, шкала - мера расхождения, метод Уорда
о его протяжённых морских миграциях. Среди эндемиков наиболее оригинальна фауна паразитов американской палии (122 вида, 47,5% эндемичных), образующей ряд экотипов, в том числе проходной (17,5% морских паразитов), и пресноводной озёрной палии (72 вида, 33,3% эндемичных), что указывает на их древность и длительную изоляцию от гольцов Евразии. В отличие от разнообразного рациона американской палии, озёрная палия использует в первую очередь глубоководных амфипод (инвазия Cystidicola stigmatura), озёрных гаммарид (заражение скребнями), сигов и других рыб (инвазия цестодами).
4.2. Фауна паразитов и экология гольцов тихоокеанской группы. У мальмы в пределах ареала отмечено максимальное разнообразие паразитов - 145 видов, она образует целый ряд экотипов (проходной, озёрно-речной, озёрный, речной и карликовые ручьевой и озёрный). Значительное разнообразие паразитов мальмы на Камчатке (74 вида) и Чукотке (55 видов) свидетельствует об оптимуме для данного вида (Леванидов, 1981) и указывает на её происхождение на севере Азии, вторым центром разнообразия паразитов мальмы является Приморье (53 вида). Северный S. m. malma и южный азиатский S. m. curilus подвиды мальмы имеют почти равное число видов паразитов (75 и 73 соответственно), в том числе 46 общих, но различаются характерными для каждого подвида паразитами (рис. 6). Паразитофауна бентофага-мальмы водоёмов Камчатки (северный подвид) имеет общие черты, но существенно различается в зависимости от кормовой базы водоёма: у речной мальмы р. Кроноцкая она заметно беднее (14 видов), чем у озёрно-речной (30 видов) и ручьевой мальмы (27 видов) мезотрофного оз. Азабачье. В оз. Ушки впервые изучена паразитофауна карликовой озёрной мальмы (Буторина и др., 2009), генетически идентичная северной мальме оз. Азабачье, но морфологически отличающаяся от неё и близкая по экологии к карликовому ручьевому экотипу этого вида. Формирование карликовых форм мальмы - результат внутривидовой конкуренции при ограниченных кормовых ресурсах (Гудков, 2003). Икра тихоокеанских лососей составляет важный элемент их питания и обеспечивает высококалорийной пищей. Полученные данные подтверждают трофическую и
Таблица 3. Видовой состав паразитов гольцов озера Начикинское (Камчатка)
| Вид паразита | Озёрно-речная мальма S. malma | Эндемичный озёрный голец Salvelinus sp. | ||
| Число рыб обследованных / зараженных | Интенсивность: пределы/ средняя | Число рыб об следованных / зараженных | Интенсивность: пределы/ средняя | |
| Leptotheca krogiusi | 6/0 | - | 4/1 | - |
| Chloromyxum coregoni | 6/0 | - | 4/3 | В массе |
| Myxobolus arcticus | 6/6 | - | Нет данных | - |
| M.neurobius? | 6/3 | - | Нет данных | - |
| Proteocephalus longicollis | 6/2 | 1-2/ 1,5 | 4/2 | 1-17/ 9,0 |
| Eubothrium salvelini | 6/0 | 0 | 4/4 | 4-8/ 5,5 |
| Diphyllobothrium sp.1 | 6/0 | 0 | + | + |
| Crepidostomum farionis | 6/3 | 1-236/ 82,3 | 4/1 | 5 |
| Cr. metoecus | 6/0 | 0 | 4/2 | 2-4/ 3,0 |
| Cucullanus truttae | 6/4 | 1-3/ 1,8 | 4/0 | 0 |
| Sterliadochona ephemeridarum | 6/1 | 9 | 4/0 | 0 |
| Anisakis simplex | 6/1 | 1 | 4/0 | 0 |
| Diplostomum sp.1 (хруст.) | 6/5 | 1-3/ 2,0 | Нет данных | - |
| D. volvens (внутр.среда глаза) | 6/6 | 12-114/ 64,2 | Нет данных | - |
1 по данным Савваитовой, Васильева (1976).
т

Рис. 6. Дендрограмма различий между популяциями мальмы по фауне паразитов, шкала - мера расхождения, метод Уорда
опическую дифференциацию мальмы и гольцов-эндемиков в озёрах Дальнее, Начикинское и Кроноцкое. Фауна паразитов ручьевой формы южной азиатской мальмы о. Итуруп и верховьев р. Фроловка в Приморье обеднена (7-8 видов) и очень сходна. Белый голец в оз. Азабачье со второго года жизни - неспециализированный хищник (инвазия (инвазия Diphyllobothrium spp., Eubothrium salvelini), в то время как в оз. Кроноц-
кое это озёрный эврифаг, использующий как рыбу, так и бентос и планктон (Буторина и др., 2008а), что определяет различия в их заражённости некоторыми гельминтами. Фауна паразитов каменного гольца р. Камчатка характеризует его как речного хищника с включением в рацион беспозвоночных. Анализ фауны паразитов эндемичных гольцов оз. Кроноцкое (Буторина и др., 2008а) показывает, что S.
 schmidti - бентофаг с широким спектром питания беспозвоночными (гаммариды, в меньшей степени – другие ракообразные, личинки амфибиотических насекомых, мелкие двустворчатые моллюски), приуроченный к литорали, а S. kronocius - специализированный озёрный ихтиофаг. У кунджи найдено 95 видов паразитов, она имеет более ограниченный ареал, чем м
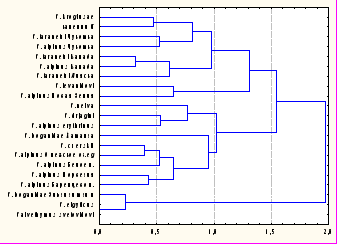
schmidti - бентофаг с широким спектром питания беспозвоночными (гаммариды, в меньшей степени – другие ракообразные, личинки амфибиотических насекомых, мелкие двустворчатые моллюски), приуроченный к литорали, а S. kronocius - специализированный озёрный ихтиофаг. У кунджи найдено 95 видов паразитов, она имеет более ограниченный ареал, чем м Рис. 7. Дендрограмма сходства арктических гольцов по фауне паразитов, шкала - коэффициент корреляции Пирсона, метод Уорда
альма и не образует такого "букета" форм, наибольшее разнообразие паразитов отмечено у неё на Камчатке (56 видов) и в Приморье (40 видов), в Японии у неё отмечено 33 вида паразитов.
4
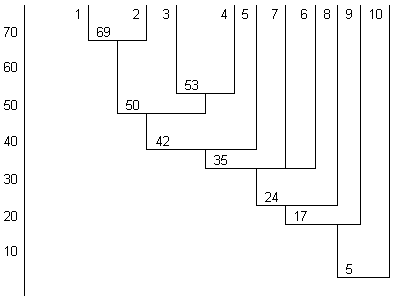
Рис. 8. Дендрограмма сходства популяций мальмы S. malma Голарктики по фауне паразитов. Обозначения: 1 - Чукотка, 2 - Kaмчатка, 3 - Приморье, 4 - бассейн Амура, 5 - Сахалин, 6 - Курильские острова, 7 - Канада, 8 - Япония, 9 - Aляска, 10 - Oрегон
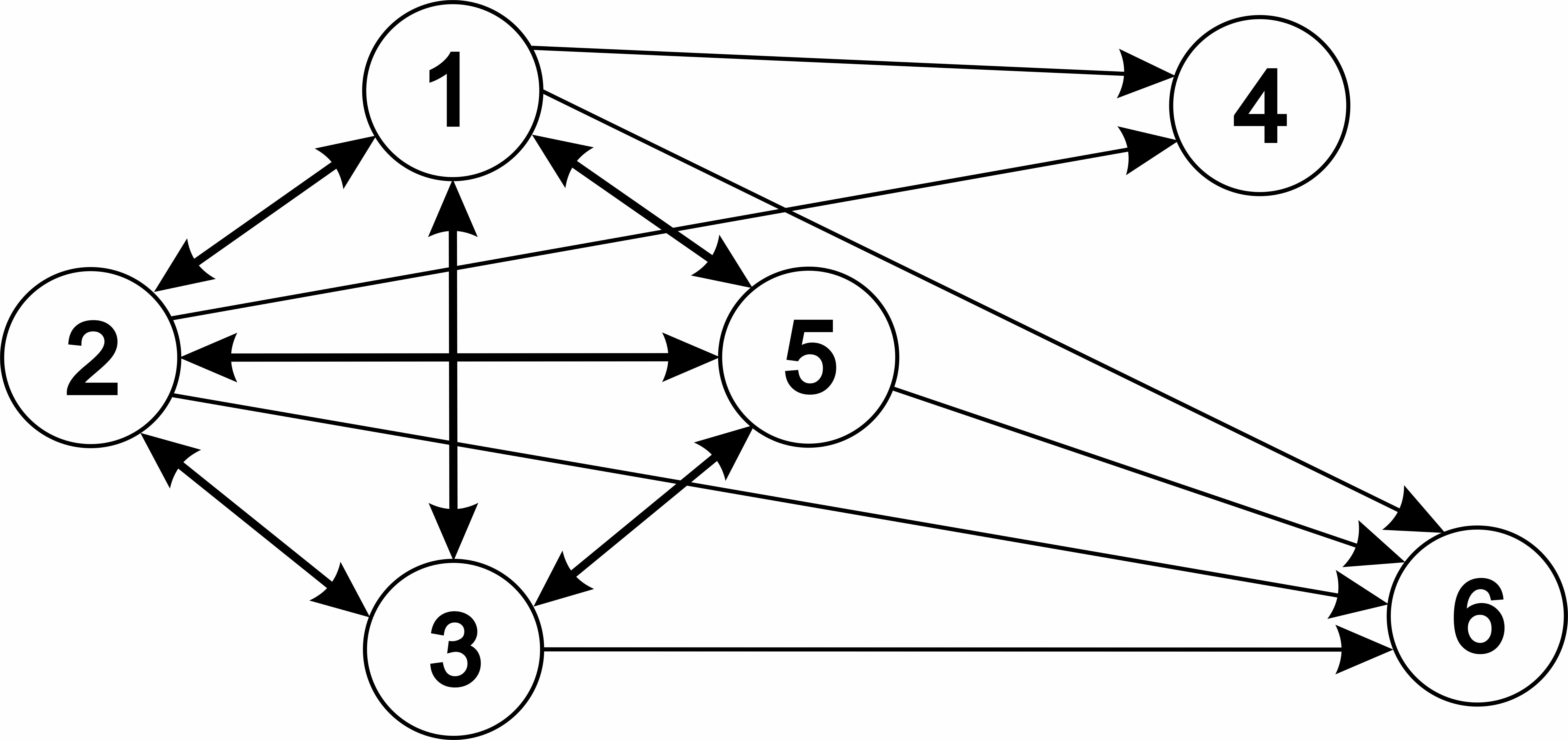
Рис. 9. Ориентированный граф или плеяда сходства популяций кунджи по фауне паразитов. Обозначения: 1 - бассейн Амура, 2 - Приморье, 3 - Сахалин, 4 - Япония, 5 - Курильские острова, 6 - Kамчатка
.3. Сравнительный анализ паразитофауны гольцов с широким ареалом. Сравнение арктических гольцов Евразии по фауне паразитов (рис. 7) позволяет выделить 3 группировки: наиболее обособленную - палий оз. Эльгыгытгын; чукотско-канадскую группу арктического гольца и гольца Таранца, с которыми сходны каменный голец и голец Крогиус Камчатки; европейско-сибирскую, которая объединяет популяции арктических гольцов, боганидскую и других палий и гольцов Дрягина, якутского и Черского, к которым близка нейва Охотского побережья. Третий кластер объединяет проходных арктических гольцов - Новой Земли и гольца Леванидова. Якутский голец по фауне паразитов очень сходен с гольцами Черского и Дрягина, по-следний близок к забайкальским гольцам, а боганидская палия Таймыра - к гольцу Черского. Эти данные указывают на происхождение изученных форм гольцов и соответствуют представлениям о формировании нескольких центров их видообразования (Behnke, 1984).
Самые высокие показатели сходства фауны паразитов получены нами для мальмы Камчатки и Чукотки (северный подвид) (рис. 8). Следующий кластер образует мальма Приморья и бассейна Амура, к которым близка мальма Сахалина (южный азиатский подвид). Фауна паразитов мальмы Курильских о-вов обеднена и наиболее сходна с таковой сахалинской мальмы, от них заметно отличается мальма Японии и Аляски, что может быть результатом изоляции. Наибольшее сходство фауны паразитов выявлено нами (рис. 9) между кунджей Приморья и бассейна Амура (ICS = 54%). Кунджа Приморья, бассейна Амура, Сахалина и Курил образует общий кластер (ICS = 46%) или плеяду. Сравнительный анализ паразитофауны кунджи позволяет дифференцировать разные популяции этого вида по образу жизни, характеру миграций (проходные и жилые, чисто речные и озёрно-речные).
4.4. Анализ паразитофауны гольцов-эндемиков. Сравнение фауны паразитов изученных видов гольцов (рис. 10) позволяет отнести большинство из них к одной из двух основных групп – арктической или тихоокеанской. Голец Леванидова принадлежит к тихоокеанской группе, к которой он близок по образу жизни и другим особенностям экологии (Черешнев и др., 2002). На дендрограмме сходства гольцов по фауне паразитов можно выделить 5 кластеров. Первый из них включает арктические виды: палий оз. Эльгыгытгын, гольца Черского, очень сходных друг с другом якутского гольца и боганидскую палию бассейна р. Хатанга, гольцов Дрягина и Таранца. Второй кластер составляют виды тихоокеанских гольцов – эндемичные гольцы оз. Кроноцкое, виды с ограниченным ареалом - каменный и белый гольцы и широко распространённые мальма и кунджа, к которым присоединяются голец Крогиус и нейва. Обособленное положение в группе занимают арктический голец и два вида американских палий, но больше всего отличается от других близкий к предку голец Леванидова. Сравнение фауны паразитов гольцов показывает их экологическую специфику, позволяет выделить кластеры европейских ледниковых реликтов и сибирских эндемиков (наиболее сходны гольцы Черского, таймырский и якутский, в целом близкие к европейской палии), палий Чукотки, гольцов-хищников Камчатки (каменный, длинноголовый, белый и голец Крогиус), озёрных эврифагов (нейва, начикинский и носатый) и американских палий. Эти данные указывают на единство происхождения европейских ледниковых реликтов и сибирских эндемиков.
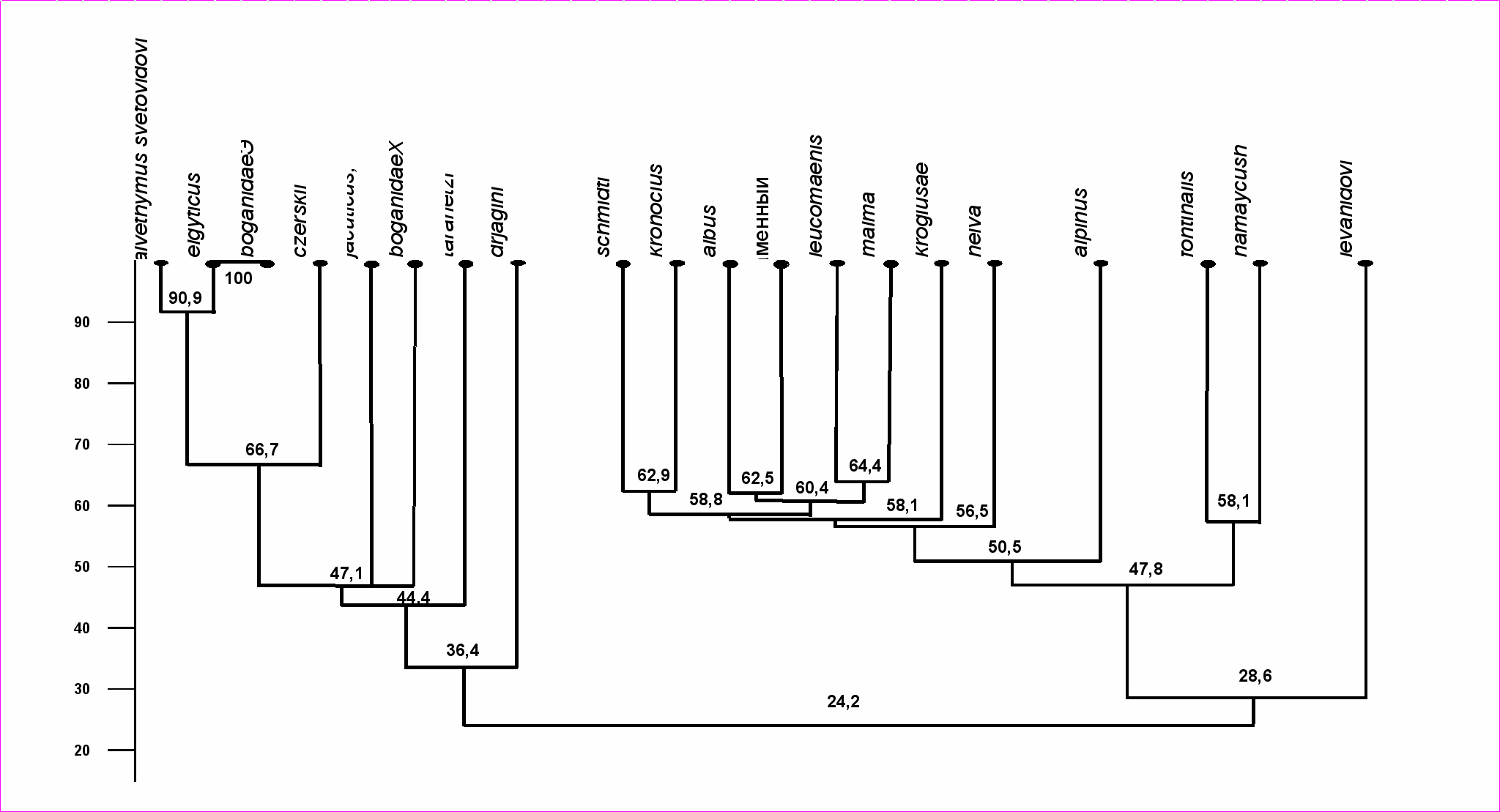
4.5. Сравнение фауны паразитов гольцов Палеарктики и Неарктики. Наглядное представление о степени сходства фауны паразитов гольцов Голарктики дает дендрит сходства (рис. 11), на нём представлены три группы гольцов. Первая объединяет арктические виды - палий оз. Эльгыгытгын и гольцов Черского, якутского, Дрягина и боганидскую палию бассейна р. Хатанга (№№14-20), вторая включает тихоокеанские виды (мальма, кунджа, белый, длинноголовый, носатый и каменный гольцы), а также гольца Крогиус и нейву (№№4-7, 9-12), третья группа - американских палий (№№1-2). Голец Таранца «связывает» эндемиков Сибири и Чукотки с тихоокеанскими видами, а арктический голец служит своеобразным «мостом» между тихоокеанскими гольцами и американскими палиями. Голец Дрягина близок к палиям, а самое обособленное положение занимает один из наиболее древних голец Леванидова. Ориентированный граф включения фауны паразитов гольцов (рис. 12) показывает, что наиболее «оригинальную» (разнообразную по составу) фауну паразитов при принятом пороге 100% имеют мальма, арктический голец и кунджа. В состав их паразитов включено большинство видов, известных у других гольцов. В состав паразитофауны мальмы на 80% включены паразиты 14 видов гольцов, она максимально отражает фауну парази-
Рис. 10. Дендрограмма сходства фауны паразитов гольцов. Шкала – коэффициент Чекановского-Сёренсена, метод максимальной связи
Рис. 11. Дендрит сходства фауны паразитов гольцов по ареалу. Обозначения: 1 - S. fontinalis, 2 - S. namaycush, 3 - S. alpinus, 4 - S. leucomaenis, 5 - S. malma, 6 - S. krogiusae, 7 - S. albus, 8 - S. levanidovi, 9 - S. schmidti, 10 - S. kronocius, 11 - каменный голец, 12 - S. neiva, 13 - S. taranetzi, 14 - S. boganidae Чукотки 15 - S. elgyticus, 16 - Salvethymus svetovidovi, 17 - S. czerskii, 18 - S. jacuticus, 19 - S. boganidae р.Хатанги, 20 - S. drjagini.
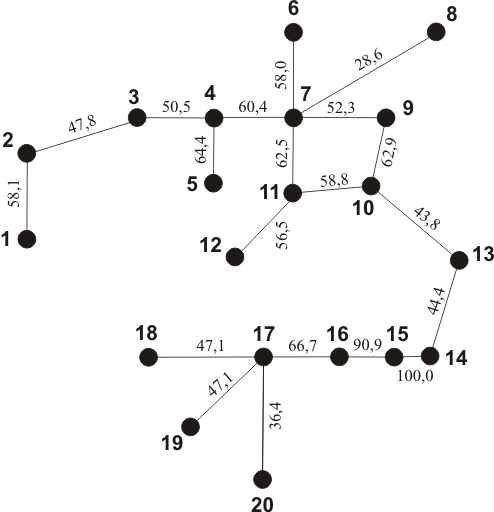
тов гольцов Голарктики, мальма имеет самый широкий спектр питания. Ареал кунджи ограничен, она освоила в основном эстуарии и бассейны крупных рек. В Арктике самый разнообразный состав фауны паразитов у арктического гольца, но он беднее, чем у мальмы. Весьма своеобразна паразитофауна американской палии, остальные эндемики имеют ограниченный ареал и состав паразитов. Американская палия заселила водоёмы различного типа, в том числе прибрежную морскую зону, что способствовало формированию разнообразных экологических форм и приобретению разнообразных видов паразитов (122 вида, не считая приобретённых при акклиматизации в Европе и Японии).
Глава 5. ЗООГЕОГРАФИЧЕСКИЙ АНАЛИЗ ФАУНЫ ПАРАЗИТОВ ГОЛЬЦОВ ГОЛАРКТИКИ
5.1. Распространение паразитов в пределах ареала гольцов. Голарктическое распространение имеют 82 вида паразитов (39,2% из 209 включённых в анализ), только в Евразии отмечены 62 вида (29,8%), в том числе 49 видов обнаружены в Палеарк- тике и 13 видов - в южной части ареала гольцов (Приморье, Япония, Сахалин). Эндемиками Северной Америки являются 65 видов (31,1%), что говорит о формировании фауны пресноводных паразитов каждого континента в условиях изоляции друг от друга. Практически для всех видов морских паразитов, за редким исключением, голь- цы являются случайными хозяевами, большая их часть относится к широко распространённым в Тихом и Атлантическом океанах видам.
Рис. 12. Ориентированный граф включения фауны паразитов гольцов Голарктики. Порог δ = 100%
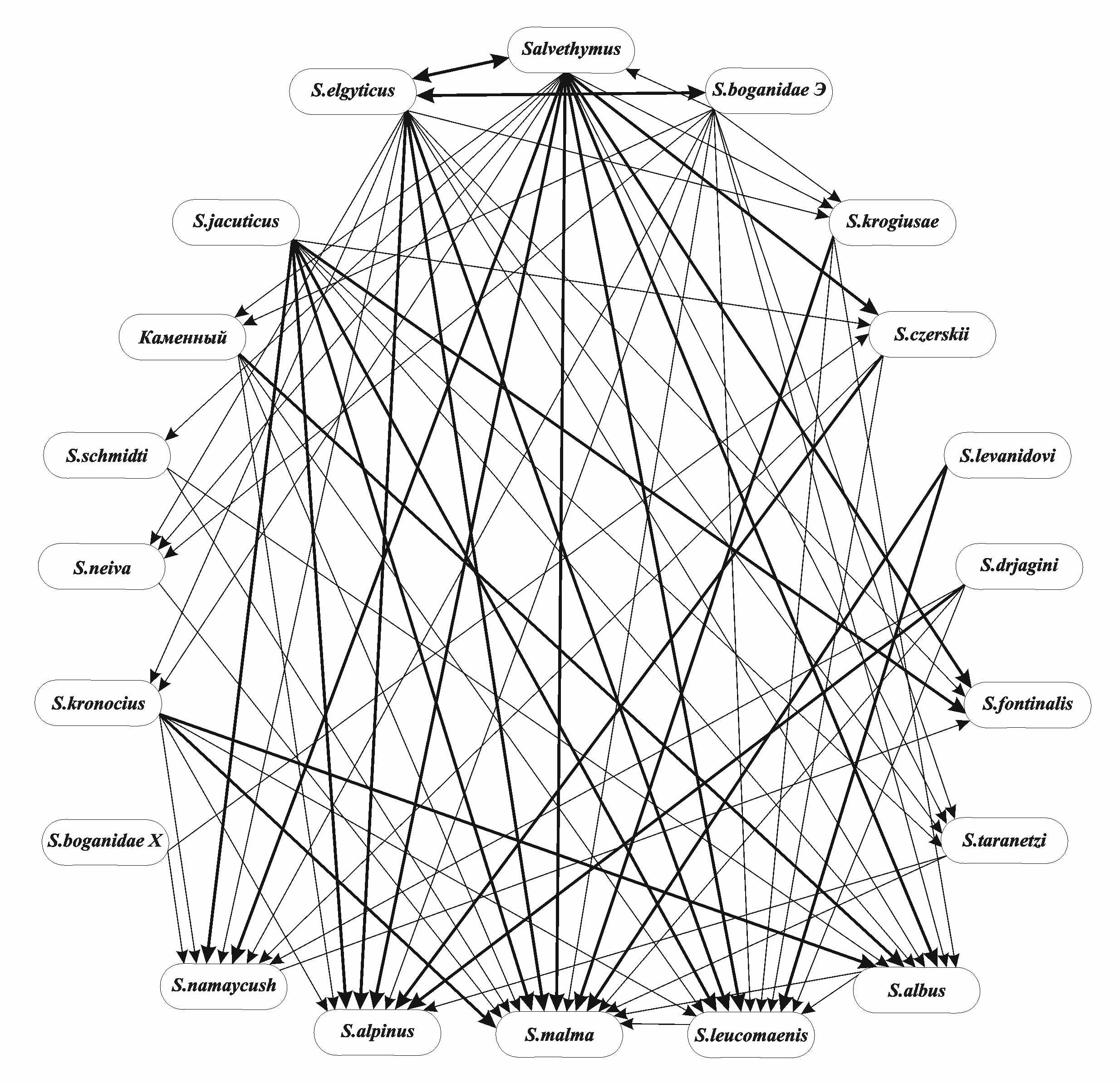
По фаунистическим комплексам распределено 198 видов паразитов гольцов. Из этого числа наибольшую часть 40,9% составляют бореальные равнинные виды (81 вид). Основной вклад в этот комплекс вносят паразиты палий Северной Америки и их гибрида, у них представители этого комплекса составляют 43,2%. Вторым по значимости в фауне паразитов гольцов является арктический пресноводный комплекс (51 вид или 25,8%), тихоокеанские предгорные (26 видов) составляют 13,1% и бореальные предгорные - 19,2% (38 видов). На долю представителей теплолюбивого китайского равнинного комплекса приходится 1,0%. Эти данные показывают, что гольцы, с одной стороны, исторически и экологически тесно связаны с родственными группами лососеобразных и имеют с ними значительное число общих холодолюбивых паразитов. С другой стороны, в процессе широкого расселения по Голарктике они обогатили свою паразитофауну за счёт включения в неё представителей бореальной равнинной фауны.
Паразитофауна северного и южного азиатского подвидов мальмы очень сходна по фаунистическому составу, морские элементы составляют основную долю у обоих подвидов (34,5-34,7%), однако процент арктических пресноводных видов выше у северной мальмы (29,8% и 17,3% соответственно). Находки 46-ти общих для обоих подвидов паразитов указывают на общее происхождение и неоднократные контакты между ними.
Cравнение фауны паразитов 25 видов и экотипов гольцов позволило выделить некоторые виды паразитов в качестве видовых дифференцирующих признаков: только для американской палии - как минимум 6-ти эндемичных видов паразитов (возможно, до 9), у южной азиатской мальмы - 5 видов, кунджи - 3 вида, у гольцов Сибири - 2 эндемичных вида паразитов.
Нематоды Сystidicola farionis и Salvelinema salmonicola являются видами, которые замещают друг друга с севера на юг ареала гольцов. В северных популяциях обычна Cy. farionis, на Камчатке их ареалы перекрываются, в Приморье и Японии у мальмы встречается только S. salmonicola (Ермоленко и др., 1998). На Камчатке в озёрах Ушки и Дальнее распространена S. salmonicola, но не встречается Cy. farionis. Южная граница ареала Cy. farionis – оз. Курильское на юге Камчатки, северная граница S. salmonicola доходит до р. Утхолок на западном побережье Камчатки (Соколов, 2005). Скребень Echinorhynchus salmonis заражает гольцов Камчатки и Чукотки, а E. cryophilus отмечен у мальмы только в Приморье и бассейне Амура. Трематода Phyllodistomum umblae - паразит гольцов Камчатки и Чукотки (Рудминайтене, Рудминайтис, 1979; Буторина 1980), американской палии и арктического гольца (возможно, гольца Таранца) в Канаде, южная граница его распространения в Азии - оз. Кроноцкое на Камчатке. Преимущественно на юге ареала (Приморье, южный Сахалин, Япония) у гольцов распространена Rhabdochona oncorhynchi, северной границей её ареала является нижнее течение р. Камчатка.
5.2. Влияние изоляции и краевых условий ареала хозяина на фауну паразитов гольцов. В оз. Кроноцкое наблюдается необычайно сильная по сравнению с другими озёрами Камчатки инвазия гольцов некоторыми гельминтами (Crepidostomum spp., E. salvelini, C. truttae, N. salmonis и др.) и ракообразными рода Salmincola (от нескольких сотен до нескольких тысяч). Мы полагаем, что здесь проявляется островной эффект Кребса – вспышка численности паразитов в изолированном озере при невозможности удаления части паразитов из-за отсутствия в озере проходных форм. Совершенно иная картина наблюдается в периферийных популяциях реликтовых палий оз. Эльгыгытгын и других в верховьях притоков р. Анадырь, которые отличаются экстремальными климатическими условиями, близкими к условиям первичных ультраолиготрофных озёр (Румянцев, 1996). Для этих маловидовых сообществ характерен чрезвычайно бедный состав ихтиофауны (Черешнев, Скопец, 1993) и фауны паразитов (всего 5 видов). У палий доминируют паразиты, промежуточными хозяевами которых служат планктонные ракообразные, планктонные трофические цепи доминируют здесь над донными (Черешнев, Скопец, 1993). Боганидская палия, как хищник, накапливает паразитов. Возникающий в таких условиях каннибализм, связанный с сезонной нехваткой пищи (Hammar, 2000), - необходимая стратегия выживания изолированных популяций гольцов в высоких широтах. Обеднение фауны паразитов наблюдается и у гольцов островных популяций - S. alpinus в Гренландии (Due, Curtis, 1995), S. fontinalis на о. Ньюфаундленд (Marcogliese, Cone, 1991). Причиной этого авторы считают отсутствие соответствующих промежуточных хозяев или контакта гольцов с другими видами рыб. У мальмы Орегона на краю ареала отмечено 3 вида (Myxosoma sp., Bunodera luciopercae и Tubulovesicula lindbergi) паразитов. Сильное обеднение фауны паразитов и приобретение неспецифичных паразитов наблюдается и при акклиматизации гольцов - значительно обеднена паразитофауна S. fontinalis в Японии - 6 видов, из которых лишь два специфичны для лососёвых (Rh. oncorhynchi, Acanthocephalus echigoensis).
Глава 6. ИСТОРИЯ ФОРМИРОВАНИЯ ПАРАЗИТОФАУНЫ ГОЛЬЦОВ
Паразитологические данные подтверждают распространённую точку зрения на эволюцию гольцов (Викторовский, 1978а; Глубоковский, 1995; Пугачёв, 1984, 1999; Черешнев, 1986). Формирование и расселение гольцов и их паразитов происходило в периоды резкого похолодания климата, интенсивного горообразования, переориентации речных систем, многократных трансгрессий и регрессий океана. Не случайно они отличаются повышенной адаптационной способностью, холодолюбивостью, реофильностью и эвригалинностью. C водоёмами горного типа связана эволюция не только лососей рода Salmo, охридских форелей рода Salmothymus и тайменей, но и гольцов (Salvelinus a. salvelinus озёр Альп, даватчана горного Забайкалья, гольцов Камчатки и др.). Подобно хариусам и форелям, предки гольцов обитали в горных водотоках. На расхождение этих групп и их дальнейшую эволюцию повлияли геогидрократические колебания уровня Мирового океана (Линдберг, 1972), которые способствовали дифференциации гольцов и форелей и формированию у тех и других полупроходных и проходных форм (Смирнов, 1979). Конкуренция с другими пресноводными видами способствовала тому, что популяции гольцов, сохранившие чисто пресноводный образ жизни, благодаря широким адаптивным возможностям, занимали верховья рек и высокогорные озёра, недоступные для других рыб. Эта способность проявляется у многих видов гольцов, нередко обитающих в водоёмах горного типа. В реках и ручьях с обеднённым планктонным населением молодь могла питаться в основном бентосом: личинками и куколками амфибиотических насекомых, падающими в воду воздушными насекомыми (озёрно-речные системы Камчатки). В пользу горного происхождения свидетельствуют находки у большинства видов гольцов бореальных предгорных элементов фауны паразитов, а также то, что в онтогенезе озёрно-речной мальмы и белого гольца происходит смена ручьевого (горного) образа жизни, типа питания и паразитофауны в первые годы жизни на озёрно-речной и озёрный, затем – на проходной. Миграционная способность мальмы, белого гольца озёр Кроноцкое, Азабачье и других видов гольцов развилась в связи с необходимостью расширить спектр питания. Эти изменения в онтогенезе гольцов, на наш взгляд, отражают исторический путь, пройденный предками в процессе становления (Буторина, 2006). Поэтому бореальные предгорные элементы в паразитофауне гольцов, по-видимому, более древние, чем арктические пресноводные. Паразитологические данные согласуются с мнением Р.М. Викторовского (1978а) и гипотезой М.Ю. Пичугина с соавторами (2006) о ручьевой мальме как исходной форме. Бореальные предгорные элементы в паразитофауне гольцов специфичны для семейства в целом. К ним относятся: Myxidium salmonis, Myxobolus neurobius, Sterliadochona ephemeridarum, Pseudocapillaria salvelini, Crepidostomum metoecus и ряд других.
Миграции на север Евразии, на что указывает значительная доля представителей арктического пресноводного комплекса в паразитофауне гольцов (25,8%), привели к освоению гольцами арктического бассейна и значительному расширению ареала. Это дало гольцам значительное преимущество перед форелями и другими холодолюбивыми рыбами бореального предгорного комплекса и позволило занять зону арктических тундр. Сопоставление ареалов арктических гольцов (палии, гольцов арктического, таймырского, Дрягина, Черского) со схемой распространения ледникового покрова на севере Европы в период его максимального развития и последующей деградации (Гроссвальд, 1999) показывает, что области распространения гольцов приходятся на бывшие приледниковые районы и зоны формирования обширных озёрно-речных систем в результате таяния ледников. Важную роль в распространении гольцов и их паразитов сыграли и морские миграции. Таким путем мальма заселила пресноводные водоёмы островов и континентальной части тихоокеанского побережья. В целом, морские миграции имели меньшее значение в освоении гольцами своего обширного ареала, чем пресноводные. На это указывает ограниченное число морских видов в фауне паразитов гольцов (17,9%). Однако доля морских элементов в фауне паразитов разных видов гольцов заметно различается. Морские миграции сыграли наиболее важную роль в становлении гольца Леванидова, обоих подвидов мальмы и кунджи, гольца Таранца, связанного в своём возникновении с пограничной между Арктикой и Пацификой зоной Берингии, эти виды и в настоящее время больше связаны с морем. Для гольцов Арктического бассейна морской путь расселения, по-видимому, имел несколько меньшее значение, они используют для нагула в основном эстуарии и опреснённые морские заливы и сравнительно слабо инвазированы морскими паразитами.
Миграция на восток - следующая волна миграции гольцов. Паразитологические данные свидетельствуют о сходстве нейвы, проникшей на тихоокеанское побережье, с арктическими гольцами. В Сибири сформировался обширный центр видообразования гольцов, включая Забайкалье, на что указывает заметное сходство фауны паразитов эндемичных гольцов Сибири и Забайкалья. В дальнейшем он подвергся значительной редукции из-за активной вулканической деятельности в этом регионе, тектонических процессов, изменений климата. На арктическом побережье также происходило распространение гольцов на восток и возникновение новых популяций, которые были описаны как самостоятельные виды и формы арктических гольцов. На Северо-Востоке Азии сформировался Чукотско-Берингийский центр видообразования гольцов. Бэлон (Balon, 1980) предполагал, что предками гольцов были небольшие специализированные рыбы, которые жили в конце третичного периода и питались планктоном. Умеренно-теплый климат первой половины плиоцена на Северо-Востоке Азии (Бискэ, 1975, Бискэ, Баранова, 1976) был благоприятным для существования в древних озёрах (типа оз. Эльгыгытгын) планктоноядных форм, аналогичных длиннопёрой палии Световидова, близкой к предку арктических гольцов (Черешнев, Скопец, 1993; Черешнев и др., 2002). Формирование нескольких центров покровных оледенений на севере Евразии и Северной Америки и арктических островах в плиоцене-плейстоцене приводило к неоднократным разрывам ареала гольцов и возникновению рефугиумов, где они находились в значительной изоляции друг от друга, что способствовало образованию эндемичных видов и форм. В дальнейшем они стали центрами расселения гольцов.
В периоды депрессии ледников по их краям образовывались многочисленные приледниковые озёра, возникла сеть озёрно-речных систем (Флинт, 1968; Митенёв, 2000 и др.), связи между которыми неоднократно исчезали и появлялись, происходила миграция и обмен гидробионтами. В этих водоёмах формировалась холодолюбивая фауна, представителями которой являются гольцы и большинство их характерных паразитов. Существенная доля арктических видов в паразитофауне гольцов показывает, что они длительное время населяют тундровые и близкие к ним северные водоёмы с обеднённым видовым составом гидробионтов. Не случайно и обилие арктических видов и форм гольцов. Арктический пресноводный комплекс связан с «реликтовыми» озерами Голарктики (Онежское, Ладожское, озёра бассейна Байкала, Карелии, Швеции, бассейна рек Обь, Енисей, Лена, Великих Североамериканских озёр и других в Северной Америке) (Иешко и др., 1985; Румянцев, 1996). В то же время мальма, распространяясь в тихоокеанском бассейне, освоила самые разнообразные биотопы, дала южные подвиды, что нашло свое отражение в максимальном разнообразии фауны паразитов. По мнению Р.М. Викторовского (1978), в тихоокеанском бассейне быстрой эволюции гольцов способствовали трансгрессии океана, тектонические процессы, вулканизм, которые уничтожали пресноводную ихтиофауну (конкурентов гольцов). Трансгрессии благоприятствовали распространению проходных форм гольцов (у мальмы, кунджи, белого гольца, гольца Леванидова). В бассейне Тихого океана возник тихоокеанский предгорный комплекс видов паразитов, связанный с родом Oncorhynchus и тихоокеанскими гольцами. К нему можно отнести: Kabatana takedai, Myxidium salvelini, Zschokkella orientalis, Leptotheca krogiusi, Chloromyxum wardi, Chloromyxum salvelini, Myxobolus krokhini, Myxosoma dermatobia, Plagioporus imanensis, Nanophyetus salmincola, Salvelinema salmonicola, Philonema oncorhynchi, Rhabdochona oncorhynchi, Echinorhynchus cryophilus, Acanthocephalus echigoensis, A. minor, Salmincola markewitschi, S. californiensis. Таким образом, у гольцов сформировался смешанный состав фауны паразитов, разных по возрасту и происхождению, ведущую роль в котором играли арктические пресноводные, бореальные и тихоокеанские предгорные холодолюбивые виды.
Приведённые в работе данные показывают, что гольцы способствовали видо- и формообразованию во многих связанных с ними специфическими и экологическими связями таксономических группах паразитов и формированию их жизненных циклов. Из 8 видов рода Salmincola, отмеченных у гольцов, S. markewitschi специфична для кунджи, S. carpionis и S. edwardsii – для мальмы и белого гольца (Шедько, Шедько, 2002), S. siscowet отмечена у арктического гольца, мальмы, озёрной палии и гибрида американской и озёрной палий (Hoffman, 1999). Находки у гольцов «сиговых» видов (S. extensus и S. corpulentus) обусловлены их совместным обитанием в одних и тех же водоёмах и трофическими связями между ними. Специфичный для арктического гольца Salmonchus alaskensis освоил разные виды гольцов и перешёл на других лососевидных (Герасев, 2004). С гольцами связано возникновение G. birmani, специфичного для гольцов бассейна Тихого океана. Миксоспоридии Chloromyxum salvelini характерны для мальмы и кунджи Японии, Ch. wardi - для мальмы и гольца Крогиус на Камчатке, американской палии и других лососёвых. Возможно, они произошли от представителя бореальной предгорной фауны Ch. truttae. Миксоспоридии Myxidium salvelini характерны для рода Salvelinus. Zschokkella salvelini найдена только у S. fontinalis в Северной Америке, а Z. orientalis отмечена у рода Salvelinus и других лососёвых азиатского побережья Пацифики. Cпецифичный для гольцов Myxobolus arcticus, по-видимому, произошёл от M. neurobius, представителя бореальной предгорной фауны, в большей степени связанного с родом Oncorhynchus. Связанный с арктическим гольцом Eubothrium salvelini (Kennedy, 1978), очевидно, дал начало эстуарному виду E. crassum, широко распространённому у лососеобразных. Pseudocapillaria salvelini - паразит арктического (Полянский, 1952) и других видов гольцов, возможно, произошёл от морфологически очень близкого к нему P. tomentosa (Moravec, 1987). Цестода Proteocephalus longicollis, исходно паразитирующая у сиговых (Аникиева, 2000), перешла на гольцов и дала мальмовую экоформу (Аникиева, 2000). Дивергенция нематод рода Philonema тесно связана с дифференциацией сиговых и лососёвых рыб - палеарктический Ph. sibirica отмечен у арктического гольца и гольца Дрягина, Ph. agubernaculum – у гольцов Канады, Ph. oncorhynchi – у гольцов Пацифики, экологически связанных с тихоокеанскими лососями рода Oncorhynchus. Cystidicola farionis отмечена в Северной Азии у разных видов гольцов, в Северной Америке она в большей степени приурочена к сиговым. Cy. stigmatura, напротив, тяготеет к гольцам, реже отмечена у сиговых и тихоокеанских лососей (Black, 1983). Очевидно, Cy. stigmatura возникла после проникновения гольцов на американский континент и специализировалась к роду Salvelinus (особенно к S. namaycush), а Cy. farionis, по-видимому, и в Палеарктике характерна для сиговых, с которых перешла на гольцов. Становление цикла развития Cy. farionis проходило в первичных олиготрофных озерах и связано с реликтовыми амфиподами (Румянцев, 1996). Нематоды Rhabdochona oncorhynchi и R. kisutchi возникли в тихоокеанском бассейне и связаны с гольцами специфическими связями. Возможно, палеарктическая R. denudata, найденная у арктического гольца и мальмы, является исходной формой для «лососёвых» видов этих нематод. Бореальный предгорный вид Crepidostomum metoecus, очевидно, является исходным для более широко распространённого у лососёвых и близких к ним рыб Cr. farionis. С гольцами связаны и скребни рода Echinorhynchus. Филогенетически самым молодым из них является E. cryophilus, представитель тихоокеанской предгорной фауны, один из самых древних – E. salmonis, формирование его жизненного цикла связано с эвтрофикацией озёр и массовым развитием реликтовых амфипод в озёрах олиготрофного типа (Румянцев, 1996). E. truttae связан в первую очередь с палеарктическими форелями и гольцами и, возможно, ещё более древний, чем E. salmonis. Пиявки Paracanthobdella livanowi и Acanthоbdella peledina произошли от общего предка, близкого к A. peledina, эволюция P. livanowi шла по пути усиления свободноживущей стадии (Пронин, 1971).
Проникновение предка американской и озёрной палий на американский континент, освоение свободных экологических ниш и пространственная изоляция послужили стимулом для эволюции самих гольцов и большой группы их специфичных паразитов. У американской палии найдено наибольшее число паразитов, специфичных к виду или роду: Dactylosoma salvelini, Haemogregarina salvelini, Sarcocystis salvelini (вид сомнителен), Eimeria salvelini, Loma fontinalis, Zschokkella salvelini, Myxosoma ovoidalis, Henneguya fontinalis, Gyrodactylus salmonis, Sanguinicola fontinalis. Другие паразиты американской палии имеют более широкий круг хозяев.
Гольцы сыграли важную роль и в расширении круга хозяев коэкологичных с ними видов паразитов. Включившись в жизненный цикл диплостоматид, стригеид, цестод рода Triaenophorus и многих других гельминтов в качестве дополнительных или резервуарных хозяев, гольцы приняли участие в становлении их жизненных циклов во многих озёрно-речных системах Голарктики.
Глава 7. ЗНАЧЕНИЕ ПАРАЗИТОЛОГИЧЕСКИХ ДАННЫХ ДЛЯ
ХОЗЯЙСТВЕННОГО ИСПОЛЬЗОВАНИЯ ГОЛЬЦОВ
Интерес к гольцам как объектам рыбохозяйственной деятельности определяется не только их промысловым значением, но и перспективностью разведения в холодноводных водоёмах Европы, Сибири и Северной Америки (Кудерский, 1984; Шевцова, 1987; Сорокин и др., 1992; Jobling et al., 1998). Американскую палию, естественный ареал которой включает только Виргинию, акклиматизировали по всему американскому континенту и Европе преимущественно для спортивного и любительского рыболовства (Fausch, White, 1981). Удобными объектами товарного рыбоводства являются быстрорастущие гибриды S. alpinus х S. fontinalis и Salmo salar х S. alpinus (Савваитова, 1989). На Северо-Западе России, в Карелии традиционным остаётся искусственное воспроизводство ладожской палии для пополнения её запасов. В странах арктического бассейна, особенно Исландии, Скандинавских странах, Канаде, США большое внимание уделяется выращиванию арктического гольца. В связи с этим, особое значение приобретает паразитологический мониторинг. Среди паразитов гольцов можно выделить около 30 видов, имеющих эпизоотическое и медицинское значение. Массовую гибель молоди лососёвых могут вызывать Hexamita truttae, Chilodonella piscicola, Trichodina truttae (Богданова, 1976; Карманова, 1998), Cryptobia salmositica (Ardelli et al., 1994; Зайцев, 2005), Gyrodactylus salaris (Bakke et al., 2007), Diphyllobothrium dendriticum (Hoffman, Dunbar, 1961). У половозрелых гольцов на Камчатке отмечены воспаление желчного пузыря (миксоспоридии рода Chloromyxum) (Буторина, 1997), ерошение чешуи (Myxosoma dermatobia) (Буторина, Горовая, 2007), язвенная болезнь (Henneguya zschokkei), циатоцефалёз, эуботриоз (Eubothrium salvelini), в Европе - пролиферативная почечная болезнь, в Японии и на Сахалине - заражение Kabatana takedai (Nagasawa et al., 1987), в Забайкалье - триенофороз (Triaenophorus nodulosus), дифиллоботриоз (Diphyllobothrium ditremum) и диплостомоз (Пронин, 1967; Буторина и др., 2007). Нематоды вызывают тяжёлые поражения плавательного пузыря гольцов (Cystidicola farionis, C. stigmatura) (Black, 1985), висцеральную адгезию и уплотнение гонад, частичную или полную деструкцию икры (Philonema oncorhynchi, Ph. agubernaculum) (Margolis, 1970; Nagasawa, 1985; Атрашкевич, Орловская, 1993). Petromyzon marinus был одной из основных причин массовой гибели озёрной палии S. namaycush в оз. Гурон (Dechtiar et al., 1988; Muzzall, Bowen, 2000).
ЗАКЛЮЧЕНИЕ
В работе дан сравнительный анализ фауны паразитов гольцов в пределах Голарк-
тики и показано, что паразитологический метод позволяет улавливать как черты сходства, так и тонкие экологические различия между разными видами и экотипами
гольцов, выявлять их экологическую специализацию, трофические особенности и биоценотические связи, специфику конкретных водоёмов. Показана возможность использования данных о паразитах для дифференциации гольцов и выяснения их происхождения. Полученные в работе данные могут быть использованы при выборе гольцов – объектов разведения, их важно учитывать при организации рыбоводных заводов для пополнения запасов гольцов, охране эндемичных и редких популяций гольцов.
Из 276 видов паразитов, отмеченных у гольцов в Голарктике, конформными для рода являются лишь некоторые: Salmincola carpionis, S. edwardsii, у тихоокеанских мальмы, кунджи и белого гольца - Gyrodactylus birmani, у кунджи - Salmincola markewitschi, у озёрной палии – Cystidicola stigmatura, у арктического гольца – Salmonchus alaskensis и Eubothrium salvelini. Дивергенция и расселение гольцов послужили стимулом для возникновения новых паразитохозяинных связей.
Выявление видов паразитов, характерных для большинства видов гольцов и других лососёвых, свидетельствует о том, что гольцы получили их от предка, эти связи сложились на начальных этапах эволюции группы. Внешнее сходство озёрной и боганидской палий, каменного гольца с древними представителями рода Hucho дополняется близким составом фауны паразитов. Наибольшее сходство паразитофауны отмечено для гольцов и благородных лососей родов Salmo, Parasalmo, что подтверждает представление об их близком родстве. Ручьевая карликовая форма свойственна разным видам гольцов (мальма, кунджа, голец Таранца, американская палия) (Иванков, Броневский, 1976; Гриценко и др., 1999), как и форелям и некоторым другим лососеобразным, что указывает на её исходный характер. Формирование у гольцов внутривидовых группировок наследственно обусловлено и генетически закреплено, как у близких к ним форелей (Дорофеева, 1988). Сходство фауны паразитов сахалинского тайменя и гольцов свидетельствует в пользу представлений С.С. Линдси (Lindsey, 1964) об общем предке тайменей и гольцов, который, по его мнению, обитал в Евразии и Северной Америке. Каменный голец напоминает древнейшего представителя гольцов – озёрную палию не только по экстерьеру (Lindsey, 1964), но и по образу жизни и характеру питания.
Чрезвычайное сходство фауны паразитов разных видов гольцов, её общность у гольцов Евразии и Северной Америки (IJ=29,9), значительная доля (41,2%) голарктических видов паразитов свидетельствуют о существовании их единого ареала (Behnke, 1980) и широкого обмена паразитами, косвенно подтверждая вывод о едином предке-основателе гольцов Евразии от Балтики до Таймыра и Забайкалья (Осинов и др., 1996).
В результате освоения гольцами водоёмов Голарктики у них сформировался смешанный состав фауны паразитов, ведущую роль в котором играют пресноводные виды (81,9%) - арктические, бореальные предгорные и тихоокеанские предгорные. Изучение паразитофауны гольцов Камчатки в возрастном аспекте (Буторина, 1980) и присутствие бореальных предгорных видов (19,2%) в фауне паразитов гольцов от европейского Севера до Канады свидетельствует об освоении их предками водоёмов горного типа. Существенная доля арктических пресноводных видов (25,8%) в фауне паразитов гольцов указывает на тесную связь гольцов с пресноводными водоёмами арктического бассейна. Многие из них представлены реликтовыми и эндемичными, изолированными друг от друга популяциями. Анализ их паразитофауны показывает заметное обеднение состава фауны паразитов, что свойственно более древним формам. Гольцы сыграли важную роль в становлении в бассейне Тихого океана паразитов тихоокеанского предгорного комплекса (13,1%). В процессе освоения гольцами пространств Евразии и Северной Америки они приобрели значительное число паразитов, представляющих бореальную равнинную фауну (40,9%). Эти данные говорят о возможно более широком ареале гольцов в прошлом в пресноводных водоёмах Голарктики.
Изучение состава фауны паразитов и особенностей питания гольцов свидетельствует об их изначальной эврибионтности и эврифагии. В разных условиях они используют массовые и наиболее доступные виды кормовых объектов: планктонных ракообразных (гольцы озёр Забайкалья, малоротая и длиннопёрая палии оз. Эльгыгытгын), беспозвоночных бентоса (гольцы большинства озёр Камчатки), в том числе глубоководных амфипод (озёрная палия, голец Крогиус, мальма оз. Азабачье), рыбу (кунджа, бычья форель, озёрная палия, нейва, палия озёр Карелии, голец Крогиус и белый, Salvelinus sp. оз. Начикинское, американская палия). Нами выделены паразиты, которые позволяют различать северный и южные подвиды мальмы, тихоокеанскую и арктическую группы видов гольцов, эндемичных гольцов Сибири, боганидскую палию озёр Таймыра и Чукотки.
Арктический голец, мальма, кунджа, американская палия отличаются от эндемичных, реликтовых и специализированных видов гольцов (озёрная палия) наличием проходного и одного или нескольких жилых экотипов и наиболее разнообразной паразитофауной. Белый голец и голец Таранца близки к ним по наличию нескольких экотипов, но имеют ограниченный ареал и меньшее число видов паразитов. Эндемичные виды гольцов и ледниковые реликты имеют обеднённый состав паразитофауны и наиболее часто занимают озёрные биотопы.
Паразитологические данные подтверждают представления о формировании нескольких центров видообразования гольцов. Заметное сходство фауны паразитов гольцов Таймыра и Забайкалья свидетельствует о существовании единого сибирского центра их формообразования, включавшем и Забайкалье. Нейва, озёрный голец оз. Начикинское, голец Крогиус по фауне паразитов во многом близки к арктическим, что указывает на пресноводный путь распространения их предков через верховья сибирских рек к тихоокеанскому побережью. Анализ паразитофауны подтверждает, что на Северо-Востоке Азии сформировался центр видообразования гольцов, давший начало гольцу Таранца, малоротой и длиннопёрой палиям и, по-видимому, боганидской палии оз. Эльгыгытгын.
Приведённые в работе новые данные о фауне паразитов гольцов некоторых озёр Камчатки (Ушки, Кроноцкое, Начикинское, Дальнее, Курсин) и Забайкалья (Фролиха, Большое Леприндо, Лёша и Камканда) дают новые свидетельства экологической специализации гольцов в пределах одного водоёма и разнообразия в выборе объектов питания. Комплексное изучение ушковского гольца показало его своеобразие как одной из форм мальмы - карликовой озёрной, близкой к ручьевому типу, позволило разграничить его от речного хищника - каменного гольца из бассейна р. Камчатка.
ВЫВОДЫ
1. Находки одних и тех же паразитов у разных видов гольцов Голарктики указывают на то, что их общий предок уже имел сложившиеся биоценотические (трофические и топические) связи с холодолюбивыми лимнофильными и реофильными беспозвоночными, благодаря которым приобрёл определённый набор паразитов, характерный для этих лососёвых.
2. Сходство фауны паразитов гольцов Евразии и Северной Америки (IJ = 29,9%) и доминирование видов с голарктическим распространением (41,2%) свидетельствуют о существовании в прошлом единого ареала гольцов и широком обмене паразитами в пределах Голарктики. Значительное преобладание пресноводных видов паразитов (81,9%) говорит о становлении гольцов в пресноводных водоёмах и использовании преимущественно пресноводных путей при расселении по ареалу.
3. Анализ фауны паразитов гольцов Голарктики показывает, что виды, представленные проходными и жилыми экотипами (арктический голец, мальма, кунджа, американская палия) имеют максимальное число видов паразитов (91-145), эндемичные и реликтовые - обеднённое (от 5 до 27 видов), специализированные (озёрная палия) - промежуточное между ними (72 вида).
4. Выявлена наибольшая общность фауны паразитов гольцов рода Salvelinus и представителей родов Salmo и Parasalmo (ICS = 58,7%), что свидетельствует об их экологическом сходстве и близком родстве. Cходство фауны паразитов гольцов и сигов (ICS = 51,5%) обусловлено их совместным обитанием в прошлом в одних и тех же приледниковых озёрах, где возникли трофические связи между ними.
5. Паразитологический метод позволил получить новые данные об экологической специфике эндемичных видов гольцов Камчатки: голец Крогиус оз. Дальнее является неспециализированным хищником, в питании которого важную роль играют гаммариды; речная северная мальма р. Кроноцкая - планкто-бентофаг, длинноголовый голец оз. Кроноцкое - специализированный хищник; Salvelinus sp. оз. Начикинское - озёрный эврифаг. В условиях изоляции у гольцов-эндемиков проявляются как обеднение состава паразитов (палии оз. Эльгыгытгын), так и вспышки численности некоторых видов (гольцы оз. Кроноцкое). Среди эндемичных видов гольцов выявлена новая форма северной мальмы - карликовая озёрная (оз. Ушки), близкая к карликовой ручьевой, показано её существенное отличие по фауне паразитов и экологическим особенностям от одиночного речного хищника каменного гольца.
6. Паразиты могут быть использованы как дифференцирующие признаки для гольцов, имеющих разное происхождение и распространение - арктических и тихоокеанских видов, эндемиков Неарктики и гольцов Палеарктики, северного и южных азиатского и американского подвидов мальмы, боганидской палии Таймыра и эндемичных чукотских палий.
7. Гетерогенный состав фауны паразитов гольцов отражает историю их становления и освоения водоёмов разного типа. Бореальные предгорные элементы (19,2%) связаны с периодом обитания предка в горных водотоках, значительная доля представителей арктического пресноводного комплекса (25,8%) отражает период освоения приледниковых водоёмов и арктических озёр, тихоокеанские предгорные виды паразитов (13,1%) приобретены в бассейне Пацифики. Преобладание представителей бореального равнинного комплекса (40,9%) обусловлено широким расселением гольцов по Голарктике.
8. Фауна паразитов северного и южного азиатского подвидов мальмы представлена почти равным числом видов (73 и 75) и сходна по видовому (46 общих видов) составу, что указывает на происхождение от общего предка и неоднократные контакты между ними, но различается долей арктических пресноводных видов.
9. Сравнительный анализ фауны паразитов гольцов Голарктики свидетельствует о существовании нескольких центров видообразования – в Европе и Сибири, включая Забайкалье (S. alpinus, S. a. erythrinus, S. boganidae, S. drjagini, S. taimyricus, S. jacuticus, S. lepechini), на Северо-Востоке Азии (S. taranetzi, S. malma malma, S. albus, S. schmidti, S. kronocius, S. krogiusae, S. elgyticus, Salvethymus svetovidovi), в бассейне Японского моря (S. leucomaenis, S. malma curilus) и Северной Америке (S. fontinalis, S. namaycush).
СПИСОК РАБОТ, ОПУБЛИКОВАННЫХ ПО ТЕМЕ ДИССЕРТАЦИИ
Статьи, опубликованные в ведущих рецензируемых научных журналах:
1. Буторина (Маховенко) Т.Е. Об особенностях паразитофауны различных форм гольца (Salvelinus alpinus L.) Камчатки // Паразитология. - 1972. - Т. 6. - Вып. 4. - С. 369-375.
2. Буторина Т.Е. Динамика паразитофауны разных форм гольцов Salvelinus
alpinus L. бассейна озера Азабачьего // Паразитология. - 1975. - Т. 9. - Вып. 3. - С. 237-246.
3. Буторина Т.Е. Изучение паразитофауны молоди лососей рода Oncorhynchus в Охотском море //Паразитология. - 1976. - Т. 10. - Вып. 1. - С. 3-8.
4. Глубоковский М.К., Буторина Т.Е. Некоторые методические подходы к изучению таксономически сложных групп лососёвых // Биология моря. -1978. - № 3. - С. 31-38.
5. Буторина Т.Е. Численность и миграции гольцов озера Азабачьего (Камчатка) // Биология моря. - 1983. - № 1. - С. 29-36.
6. Буторина Т.Е. Новая фуркоцеркария из пресноводных озёр Камчатки // Паразитология. - 1988а. - Т. 22. - Вып. 3. - С. 247-250.
7. Буторина Т.Е. О роли миног в жизненном цикле нематод лососёвых рыб Камчатки // Биология моря. - 1988б. - № 4. - С. 66-67.
8. Ермоленко А.В., Буторина Т.Е. Паразитофауна симы Приморского края // Паразитология. - 1988. - Т. 22. - Вып. 4. - С. 278-284.
9. Буторина Т.Е., Шедько М.Б. Об использовании паразитологических данных для дифференциации молоди нерки в озере Азабачье // Паразитология. - 1989. - Т. 23. - Вып. 4. - С. 302-308.
10. Буторина Т.Е. Ксифидиоцеркарии из пресноводных моллюсков Камчатки // Паразиты морских и проходных рыб бассейна Тихого океана // Изв. ТИНРО. - 1994. - Т. 117. - С. 164-168.
11. Буторина Т.Е. О фауне паразитов экологически эквивалентных видов и экотипов гольцов водоёмов Камчатки // СЭЖ. - 2003. - № 3. - С. 279-287.