
Экобиологические механизмы акустического и теплового взаимодействия пчел 03. 02. 08 Экология
| Вид материала | Автореферат |
- Перечислить формы и механизмы взаимодействия национальных культурных центров между, 87.24kb.
- Перечислить формы и механизмы взаимодействия национальных культурных центров между, 37.75kb.
- Докладе рассматриваются две задачи о распространении тепловой волны в образце цилиндрической, 9.26kb.
- План I. Введение. II. Цель и задачи работы. III. Происхождение антибиотиков в пчелином, 649.46kb.
- М. В. Лупанов описывает ульи собственной конструкции с квадратными рамками, которые, 726.39kb.
- «Биология и патология пчел» Специальность 111201 Ветеринария Пояснительная записка, 157.25kb.
- Сессия Американского Акустического Общества, г. Провиденс, 2006, международный, 1000,, 65.95kb.
- Эколого-Токсикологические основы системы защиты медоносных пчел от болезней и вредителей, 608.83kb.
- Эколого-Токсикологические основы системы защиты медоносных пчел от болезней и вредителей, 609.24kb.
- Ветеринарные требования при импорте в российскую федерацию медоносных пчел, шмелей, 49.98kb.
МОДЕЛИРОВАНИЕ МЕХАНИЗМОВ ТЕРМОРЕГУЛЯЦИИ
Агрегировавшиеся пчелы находятся в определенном теплофизическом отношении с окружающей средой и, так же как и любое физическое тело, подчиняются законам теплообмена. Однако существование физиологических реакций теплопродукции и теплоотдачи затрудняет выяснение основных теплофизических закономерностей, изучение которых принципиально для понимания механизмов терморегуляции, используемых пчелами и другими социальными насекомыми.
Теплофизическая модель консолидированного скопления пчел.
Моделировали консолидированные скопления пчел, регулирующих температуру при заданных изменениях внешней температуры. В модель заложена гипотеза регуляции температуры в центре и на периферии гнезда по ее выходу из определенной зоны. Поддержание температурного гомеостаза осуществляется двумя идентичными регуляторами, настроенными на разные интервалы температур, значения которых можно задавать в зависимости от реализуемой модели. «Комфортными» для пчел взяты значения температур в центральной части гнезда 24…32 оС (в отсутствие расплода). С появлением расплода этот диапазон должен составлять 33,5…35,5 оС. На периферии гнезда температура регулировалась в пределах 8…13,5 оС.
Электрическая схема предлагаемой модели показана на рис. 7. Система регулирования температуры теплового центра Р1 настроена на температурный интервал 24…32 оС (пассивный период). При выходе температуры из этого интервала включается или дополнительный нагреватель
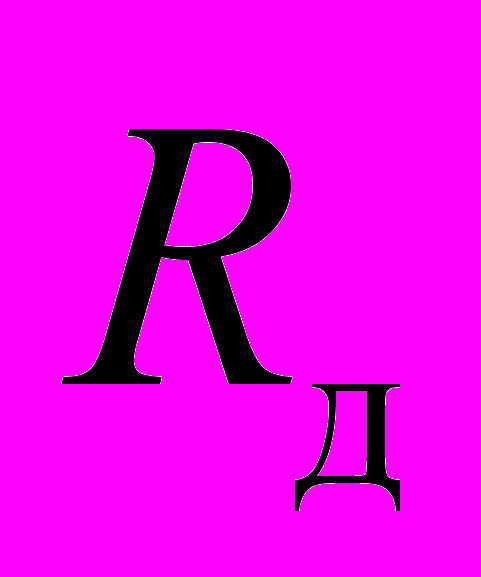 , имитируя увеличение теплопродукции пчел, или резистор
, имитируя увеличение теплопродукции пчел, или резистор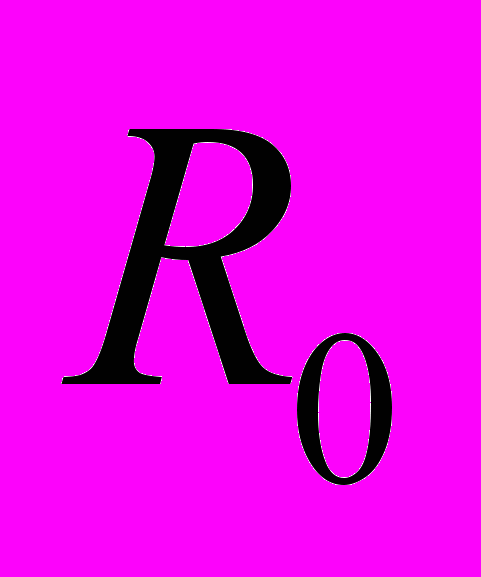 , что соответствует увеличению теплоотдачи при перегреве. Аналогично работал регулятор периферической температуры, поддерживая ее значение в пределах 8…13,5 оС. Мощность нагревателя
, что соответствует увеличению теплоотдачи при перегреве. Аналогично работал регулятор периферической температуры, поддерживая ее значение в пределах 8…13,5 оС. Мощность нагревателя 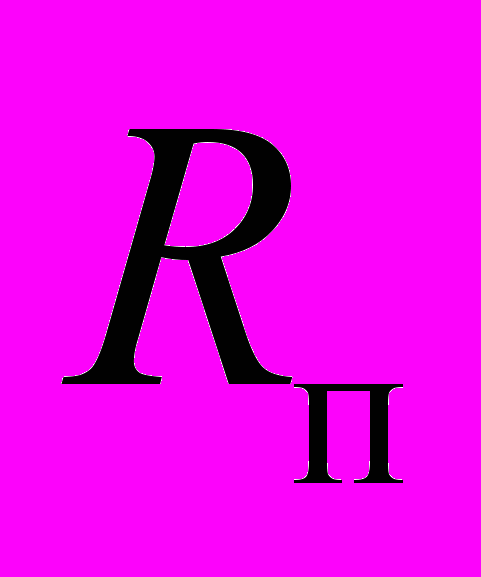 выбирали таким образом, чтобы средний уровень метаболизма покоя составлял 3…5 Вт.
выбирали таким образом, чтобы средний уровень метаболизма покоя составлял 3…5 Вт. 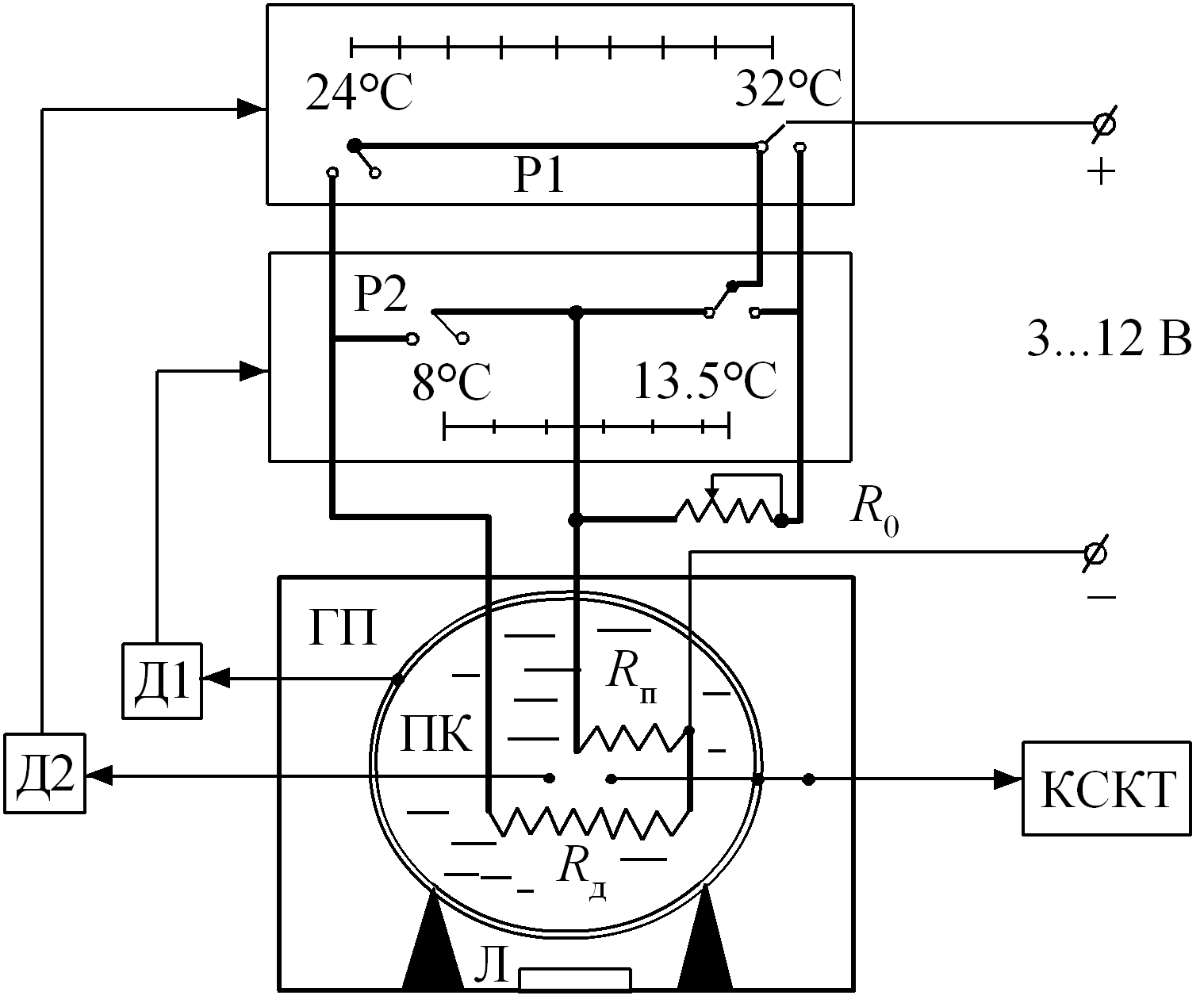
Рис. 7. Принципиальная схема терморегуляции при холодовой агрегации пчел. Принятые обозначения: Р1 – регулятор температуры теплового центра гнезда; Р2 – регулятор температуры на периферии; Д1 – датчик периферической температуры; Д2 – датчик центральной температуры; ГП – гнездо пчел; ПК – пчелиный клуб; Л – леток; КСКТ – компьютерная система контроля температуры;
– переменный резистор, находящийся вне гнезда пчел и имитирующий теплоотдачу с поверхности скопления; – нагреватель, моделирующий метаболизм покоя пчел; – нагреватель, моделирующий увеличение теплопродукции за счет микровибраций грудных мышц при охлаждении гнездаВ ответ на охлаждение включался нагреватель
, увеличивая общий уровень метаболизма до 10 Вт. Имитация реакции увеличения теплоотдачи осуществлялась включением резистора , который уменьшает мощность нагрева примерно на 40%. Эти значения мощностей подбирались изменением напряжения питания нагревателей и сопротивления резистора . Питание нагревателей осуществлялось от стабилизированного источника питания.На рис. 8 приведены характерные изменения температуры в центре модели агрегации пчел в ответ на охлаждение окружающей среды. Как видно из рисунка, возможно большое разнообразие в поведении температуры в центральной зоне, особенно в начальный период возникновения ответных реакций терморегуляции в зависимости от исходного теплового состояния. Температура в центре может изменяться в направлении температурного стимула, оставаться на прежнем уровне или изменяться в противоположном направлении. Изменения температуры в центре и на периферии гнезда приводят к изменению и средней температуры, энергетической мерой которой может служить изменение теплосодержания.

Рис. 8. Динамика изменения температуры в центре модели холодовой агрегации пчел на скачкообразное (
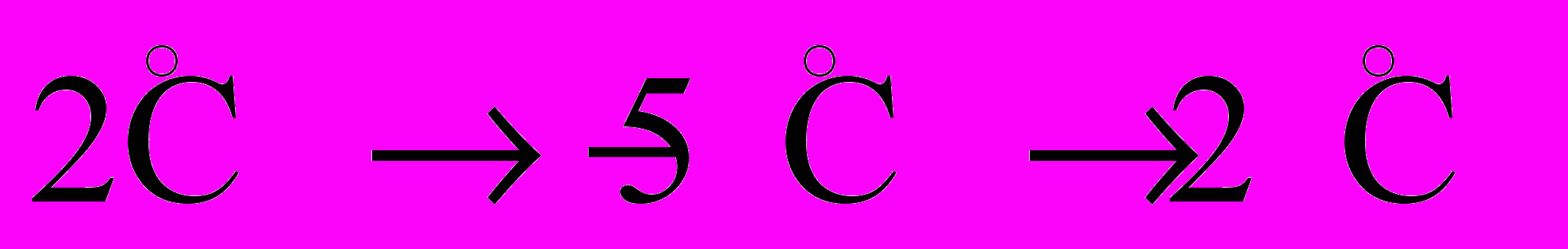 ) изменение температуры внешней среды (сплошная линия). Линии (2,3,4) соответствуют изменению температуры в центре модели агрегации при разных начальных значениях теплосодержания. Стрелкой показан момент включения нагревателя
) изменение температуры внешней среды (сплошная линия). Линии (2,3,4) соответствуют изменению температуры в центре модели агрегации при разных начальных значениях теплосодержания. Стрелкой показан момент включения нагревателя При этом реакция модели зимующих пчел на холодовое воздействие зависит от начального уровня теплосодержания. Как повышение теплосодержания, так и его снижение на определенную пороговую величину приводит к включению терморегуляторной реакции, направленной на отдачу тепла или на его сохранение.
Анализ специфики функционирования модели позволяет объяснить ряд противоречивых тенденций в поведении механизмов терморегуляции пчелиных семей. Функционирование агрегировавшихся пчел, согласно данной модели, направлено на сохранение такого соотношения между теплопродукцией (
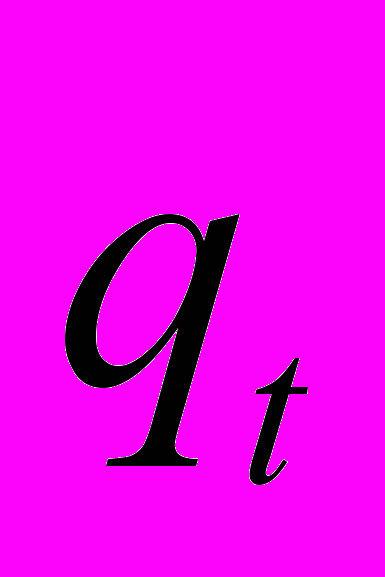 ) и теплоотдачей (
) и теплоотдачей (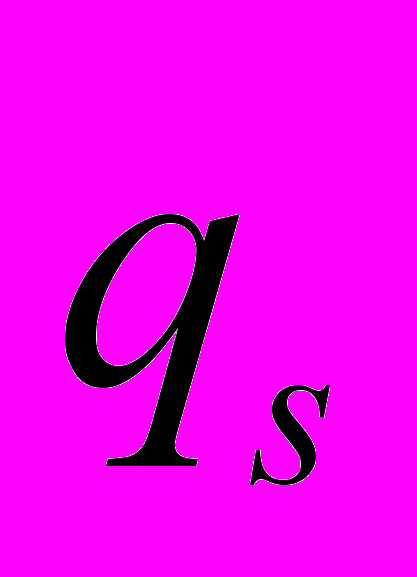 ), когда изменение теплосодержания остается постоянным (
), когда изменение теплосодержания остается постоянным (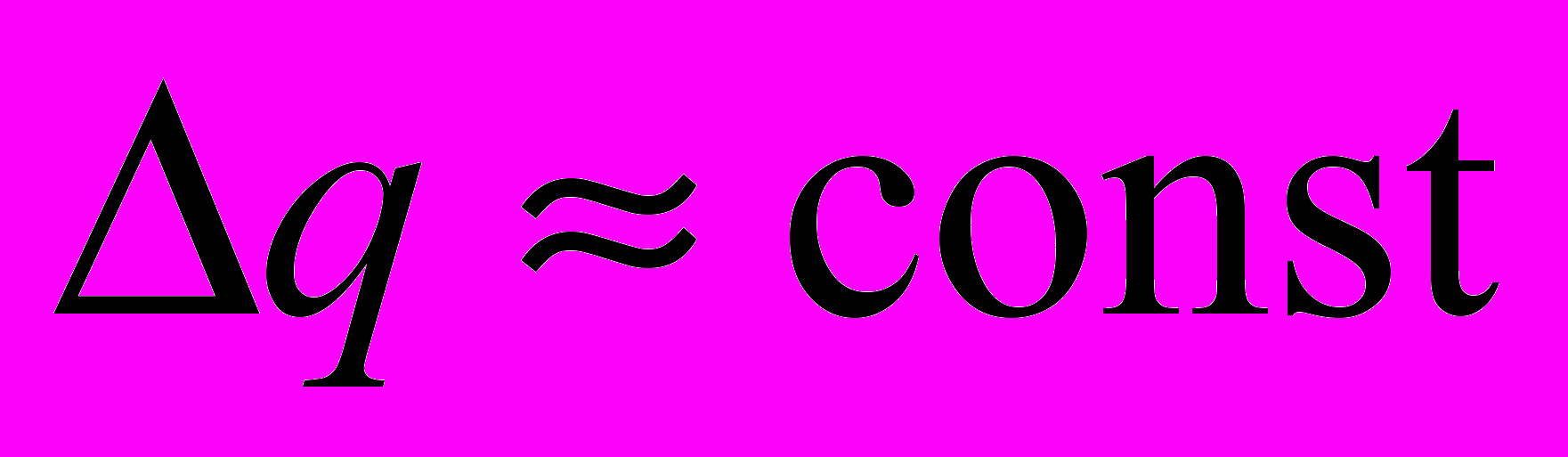 ) при определенных температурах. Другими словами, в определенном интервале температур (зона оптимальности) должно осуществляться динамическое равновесие между теплопродукцией и теплоотдачей, обеспечивающее соответствующие и относительно постоянные для каждого периода зимовки температуры внутри и на поверхности скопления. Теплопродукция и теплоотдача выступают в качестве регулирующего воздействия по отношению к теплосодержанию. Вне зоны оптимальности теплосодержание, определяемое, прежде всего, геометрией агрегации и массой пчел в нем, претерпевает весьма заметные колебания вследствие неравенства теплопродукции и теплоотдачи. Следует отметить, что изменение теплосодержания может происходить под действием не только внешних, но и внутренних причин, например при нарушениях влажностного режима, увеличении концентрации углекислого газа в тепловом центре скопления, при патологических изменениях.
) при определенных температурах. Другими словами, в определенном интервале температур (зона оптимальности) должно осуществляться динамическое равновесие между теплопродукцией и теплоотдачей, обеспечивающее соответствующие и относительно постоянные для каждого периода зимовки температуры внутри и на поверхности скопления. Теплопродукция и теплоотдача выступают в качестве регулирующего воздействия по отношению к теплосодержанию. Вне зоны оптимальности теплосодержание, определяемое, прежде всего, геометрией агрегации и массой пчел в нем, претерпевает весьма заметные колебания вследствие неравенства теплопродукции и теплоотдачи. Следует отметить, что изменение теплосодержания может происходить под действием не только внешних, но и внутренних причин, например при нарушениях влажностного режима, увеличении концентрации углекислого газа в тепловом центре скопления, при патологических изменениях.Рассмотренная теплофизическая модель позволяет утверждать, что теплосодержание является интегральной величиной, отражающей тепловое состояние зимнего скопления пчел. В этом случае система терморегуляции управляет уровнем теплонакопления в скоплении, а не регулирует непосредственно температуру в определенной точке.
При определении адекватного соотношения между теплопродукцией, изменением теплосодержания и температурой окружающей среды разработанная модель позволяет воспроизводить и анализировать процессы теплового взаимодействия агрегирующихся пчел с внешней средой в статических и динамических условиях.
Математическая модель холодовой агрегации пчел.
Построение и исследование математических моделей распределения температурных полей основаны на изучении натурных процессов теплопродукции и теплоотдачи, происходящих в скоплениях пчел. Для изучения распределения тепловых полей и тепловыделения скоплениями агрегирующихся пчел применяли тепловизор ИРТИС-2000. На основании выполненных тепловизионных исследований предложена математическая модель распределения температурных полей, позволяющая прогнозировать механизмы генерации, накопления и рассеяния тепла в самоорганизующихся биосистемах, анализировать динамические связи и взаимодействия в них.
Моделируя механизмы терморегуляции, исходили из того, что скопления агрегирующихся пчел представляют собой сферически симметричные пространственные структуры, внутри которых теплоотдача происходит посредством проводимости через пчел и частично конвекцией через воздушные зазоры между пчелами. Основным стимулом, на который агрегирующиеся насекомые реагируют изменением занимаемого объема, является внешняя температура. От нее и локализации в скоплении зависит стратегия поведения каждой пчелы, ее участие в терморегуляции. Пороговое значение для стимуляции теплогенерации посредством микровибраций грудной мускулатуры непрямого действия находится на уровне 18оС, но при 13.5 оС начинается неглубокое холодовое оцепенение, отражающееся на резком снижении обменных процессов.
Регулирование температуры в определенных внутренних зонах скоплений рассматривается как результат деятельности каждой из пчел, стремящейся остаться при температуре, удовлетворяющей ее физиологическим потребностям. Рассмотренная модель количественно передает основные закономерности, характеризующие динамику тепловых процессов в скоплениях, стимулируемых охлаждением агрегирующихся насекомых. В частности, с высокой надежностью предсказываются повышение температуры в центре в ответ на возрастающее охлаждение периферической части модели, ход температурных профилей (рис. 9), а также условия, стимулирующие дополнительное тепловыделение и терморегуляторную функцию пчел, непосредственно примыкающих к тем из них, которые образуют периферию скопления. Модель, реагируя на колебания внешней температуры, изменяет форму и плотность агрегирования всего скопления. Уплотнение скопления, связанное с интенсификацией охлаждения, отражается на уменьшении теплоизоляции поверхностного слоя агрегирующихся насекомых.
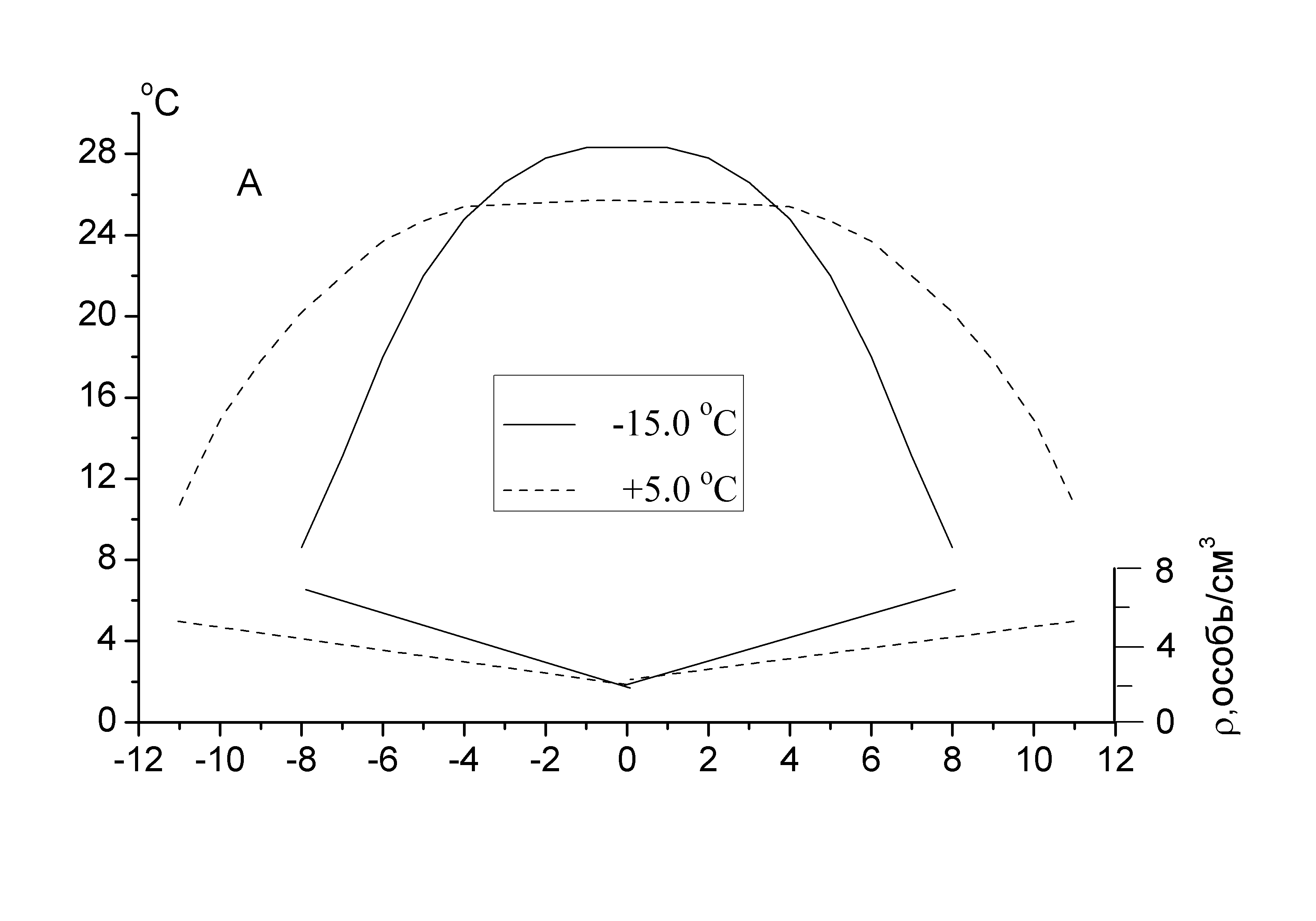
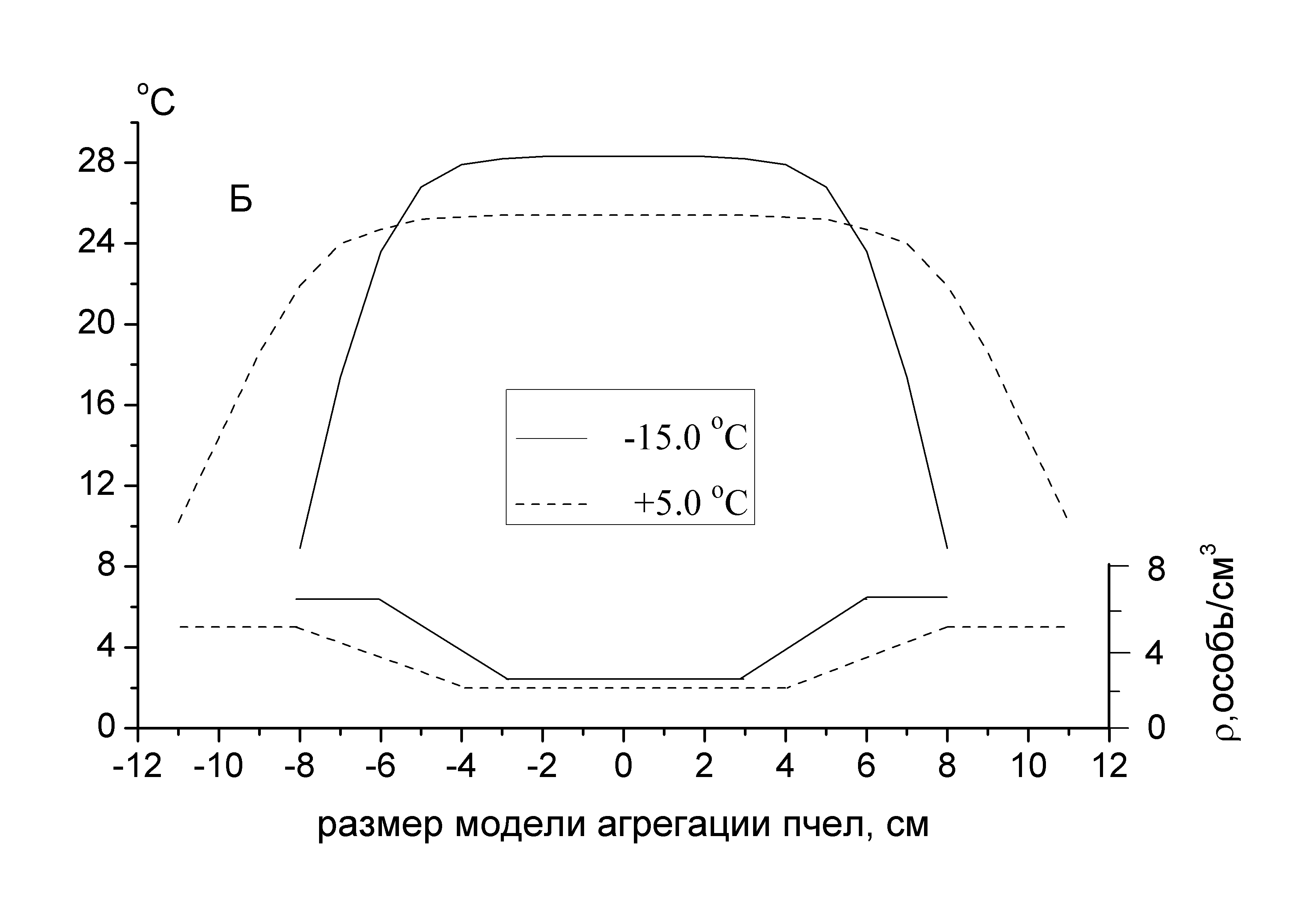
Рис. 9. Температурные профили при заданной плотности распределения пчел (особь/см3) внутри модели (А – линейная, Б – ступенчатая) в зависимости от внешней температуры. В расчетах приняты следующие значения параметров модели:
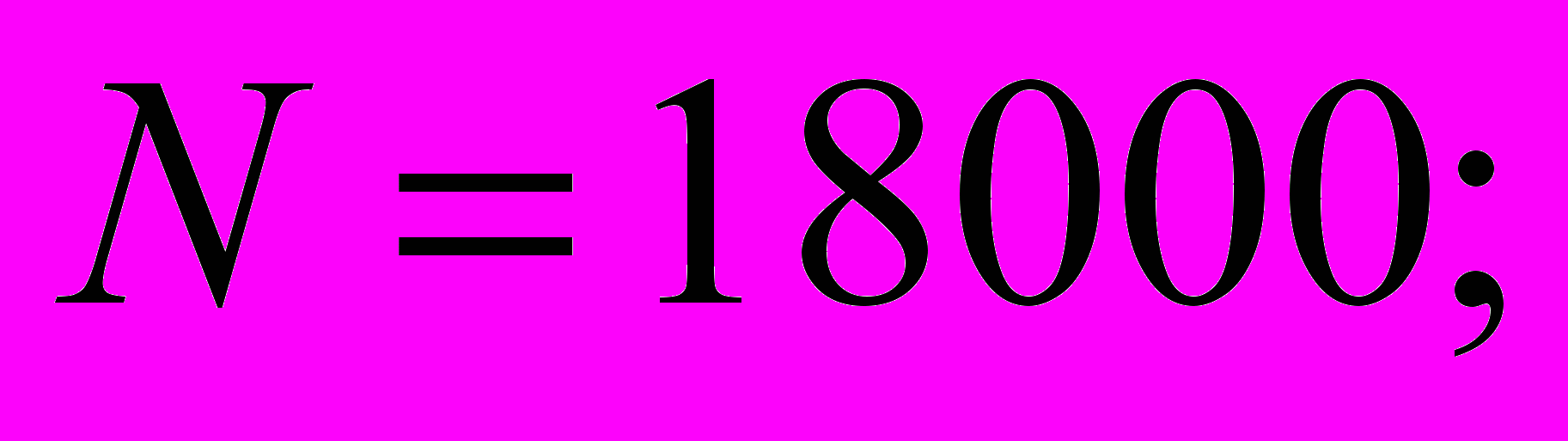
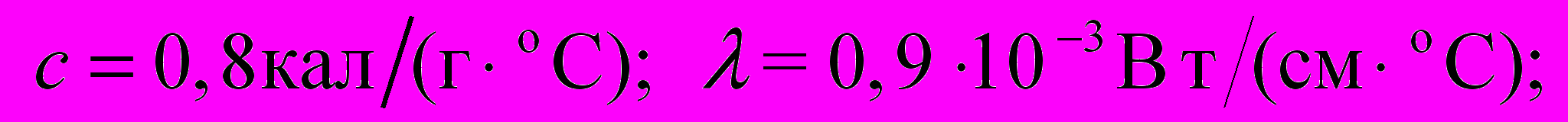
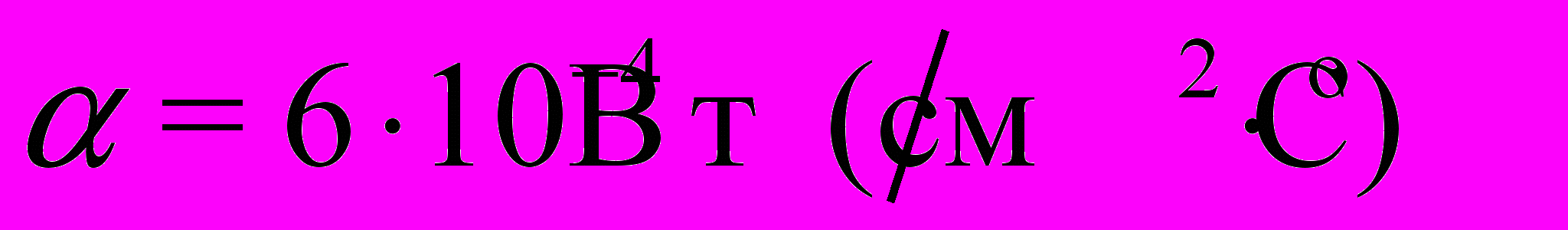 .
.Из результатов моделирования также следует, что экономии затрат энергии саморегулирующейся системой способствуют случайные перераспределения составляющих ее элементов (пчел), но закономерные изменения интенсивности их функционирования обусловлены локализацией в системе (в скоплении).
АКУСТИЧЕСКИЕ ПРОЦЕССЫ В ГНЕЗДАХ ПЧЕЛИНЫХ СЕМЕЙ
Известно, что передача информации (коммуникации) внутри пчелиной семьи осуществляется с помощью разнообразных сигналов: химических, электрических, акустических, тактильных, оптических. Из всех этих каналов связи наибольшей оперативностью обладают акустические сигналы (вибрации субстрата и звуки, передаваемые по воздуху). Скоординированная акустическая коммуникация отдельных групп пчел, выполняющих различные функции, способствует распространению информации и обмену ею между особями, что позволяет пчелиной семье функционировать как единый организм. Поэтому любые изменения состояния семьи, связанные с внешними воздействиями или внутренними биологическими процессами, отражаются на характере генерируемых пчелами акустических сигналов. По-видимому, «способность слышать» у пчел отнюдь не ограничивается восприятием только звуковых волн сжатия, передаваемых (распространяющихся) по воздуху. Рассматриваемые нами сигналы традиционно обозначаются как «звуковые», так как в их состав входит составляющая, которую может слышать человеческое ухо и которая может быть измерена с помощью микрофона.
Общая характеристика акустического шума пчелиной семьи
Суммарный акустический шум (звуковой фон пчелиной семьи) складывается из очень многих составляющих, которые генерируются отдельными особями. Их интенсивность, временная и частотная структура зависят от физиологического состояния пчел. В результате наложения собственных колебаний (микровибраций) торакса и взмахов летательного аппарата отдельных особей с меняющейся во времени амплитудой и различными периодами, а также происходящей между ними интерференции сам суммарный акустический шум и его спектр получаются нестационарными. Из них очень сложно выделить устойчивые информативные признаки, по которым можно было бы проводить идентификацию состояния пчелиных семей.
Повышению надежности анализа акустического шума способствует применение предлагаемого нами неинвазивного метода, основанного на выделении статистически однородных и часто повторяющихся фрагментов, которые представляют собой «истинный» общий фон, структура которого связана с физиологическим состоянием пчел. Выделенные фрагменты шума имеют одинаковую структуру и не содержат звуков отдельных пчел, представляют собой локальные участки с неупорядоченными и нехарактерными компонентами. Составленные таким образом участки в дальнейшем используются для идентификации множества физиологических состояний пчелиных семей, что позволит формировать их коллективные и статистически однородные «акустические портреты». Участки с одинаковой статистической структурой шума (кластеры) анализируются во временной и в частотной областях. Основная идея предлагаемого метода связана с выделением статистически однородных (наиболее сильно коррелированных) и часто повторяющихся фрагментов акустического шума, которые представляют «истинный» шум для данного распознаваемого физиологического состояния пчел.
Предложенный метод анализа акустических шумов включает в себя три основных этапа.
Этап 1. Выделение особенностей и выбор оптимальной длины фрагмента (участков) шума. На первом этапе методом рекуррентного интегрирования выявляются основные тенденции изменения амплитудных значений в выделенных участках анализируемого шума по их убыванию или возрастанию. Это позволяет находить статистически однородные участки, сравнивать их по величине функции корреляции и детектировать внутренние (т.е. не навязанные исследователем) диагностические признаки. Они выделяются по последовательности ранжированных амплитуд относительных флуктуаций (ПРАОФ) и трендам, полученным с помощью процедуры оптимального линейного сглаживания по минимальной относительной ошибке. Длина участков фрагментации и корреляционная зависимость между трендами различных участков определяется по обобщенной функции корреляции Пирсона.
Этап 2. Кластеризация выделенных фрагментов по статистически однородному признаку (признаковое описание). Использование последовательности ранжированных амплитуд позволяло выражать количественно относительные флуктуации в терминах некоего «универсального» набора редуцированных (подгоночных) параметров, входящих в приближенное аналитическое выражение для ПРАОФ. Это требуется для количественного сопоставления произвольных участков акустического шума. Увеличивая число подгоночных параметров, можно при необходимости выявить количественные различия между двумя участками шума. Функция обобщенного среднего (ФОС), выраженная в терминах высших и дробных моментов, позволяет предсказывать возможное поведение анализируемого случайного шума.
Этап 3. Изучение полученных закономерностей на основе кластеризации фрагментов шума и визуализация найденных кластеров методами, упрощающими анализ полученных результатов.
Для визуализации и количественного сравнения различных участков акустического шума помимо вышеуказанных пунктов предлагается использовать статистику дробных моментов.
Анализ коротких фрагментов неупорядоченного акустического шума пчел, описывающих относительные флуктуации, позволяет выявлять закономерности изменения его свойств, определяемых алгоритмом фрагментации. Нахождение кластеров (статистически однородных участков) позволяет определить во временной области фоновые участки сигнала, скрытые периодичности, нехарактерные (маргинальные) особенности. В частотной области открывается возможность сравнения традиционным спектроскопическим методом (по интенсивности и полуширине) динамики изменения основных пиков Фурье-компонент сигнала и нахождения геометрически подобных (фрактальных) участков спектра для выборок разного объема. По динамике изменения трендов (сглаженной кривой) и последовательности ранжированных амплитуд относительных флуктуаций открывается возможность прогнозирования вероятных изменений физиологического состояния пчел. Подчеркнем, что при расчетах такого рода не вносится никаких неконтролируемых ошибок и не принимается никаких априорных гипотез о статистической природе звукового шума пчел.
Акустические процессы в размножающихся пчелиных семьях
В течение периода подготовки пчелиных семей к размножению, занимающего в зависимости от их состояния и экологической ситуации обычно от двух до трех недель, происходит изменение генерируемых ими звуков. В размножавшихся семьях значительные изменения временной структуры акустических шумов наблюдались за 15 суток до вылета роев. По результатам анализа акустических процессов в терминах новых количественных характеристик (ПРАОФ и ФОС) установлено, что приближению отделения от материнской семьи роя сопутствует увеличение в их временной структуре статистически однородных участков. Их длина и оптимальная длительность разбиения фрагментов опт возрастает. В частности, за 10 дней до выхода роя опт равнялась 48 мс, а за сутки – достигала 128 мс. Общая продолжительность статистически однородных участков увеличилась в среднем с 0,3 до 0,5 с в каждом фрагменте анализируемого акустического процесса длительностью 1 с. По-видимому, это связано с возрастающей синхронизацией звуков, генерируемых пчелами, готовящимися отделиться от материнской семьи, что подтверждают результаты анализа трендов и функций обобщенного среднего. Непосредственно перед выходом роя интегрированные последовательности выделенных участков акустического шума становятся детрендированными, что выражается в уменьшении до несущественных значений сглаженных значений Js относительно оси абсцисс.
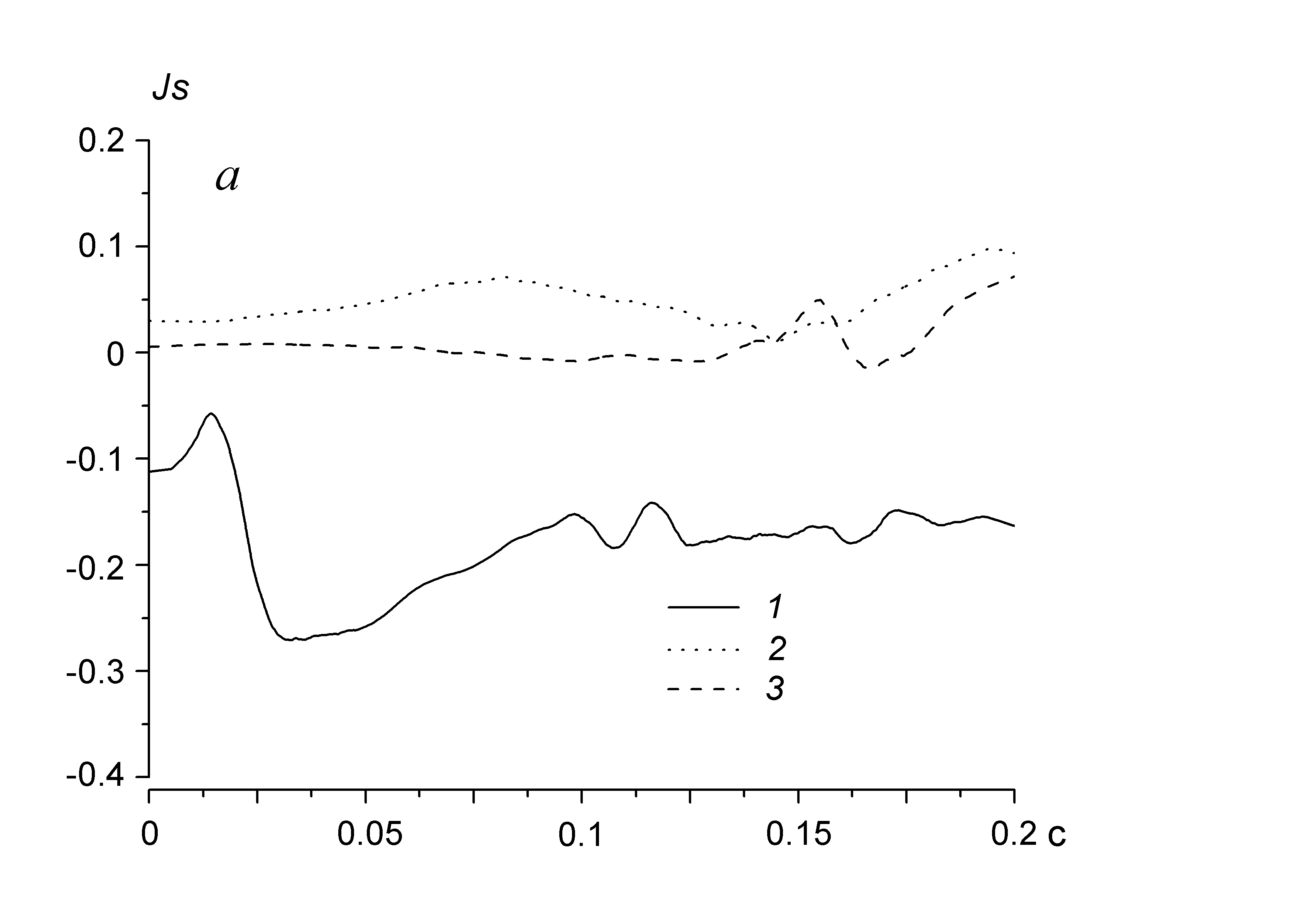
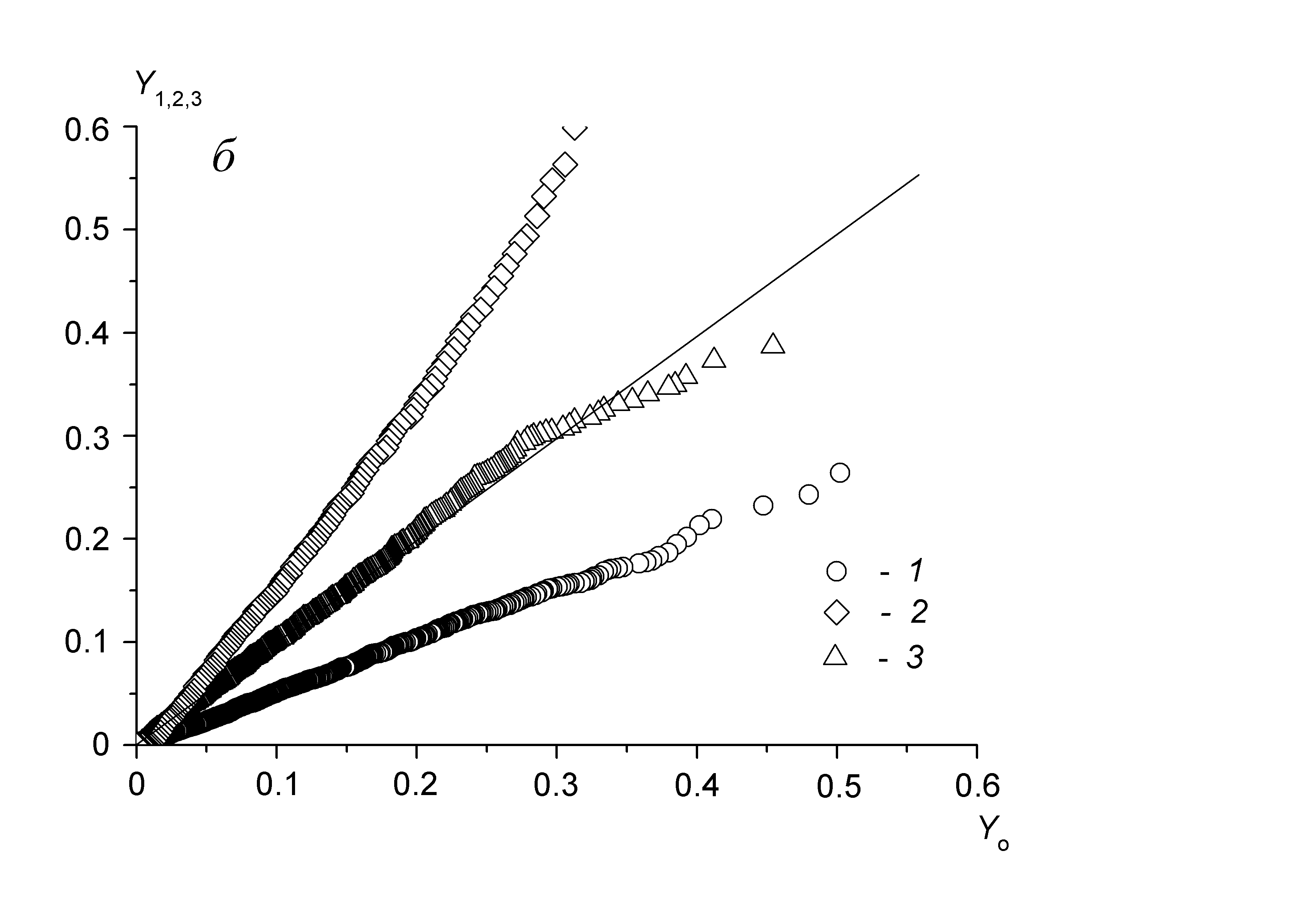
Рис. 10. Динамика усредненных значений (трендов Js) проинтегрированной последовательности (а) для выделенных участков акустического шума и взаимная корреляция ПРА относительных флуктуаций (б): 1 – семья, не размножавшаяся и не готовившаяся к размножению в течение сезона наблюдений (контроль); 2 – семья в период подготовки к размножению за 10 дней до выхода роя; 3 – эта же семья за день до роения
Сильная корреляция шума пчел (в терминах ПРАОФ) определялась линейной зависимостью
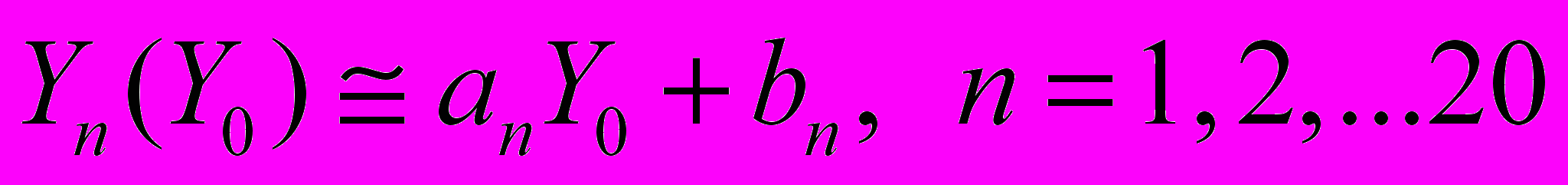 ,
,где
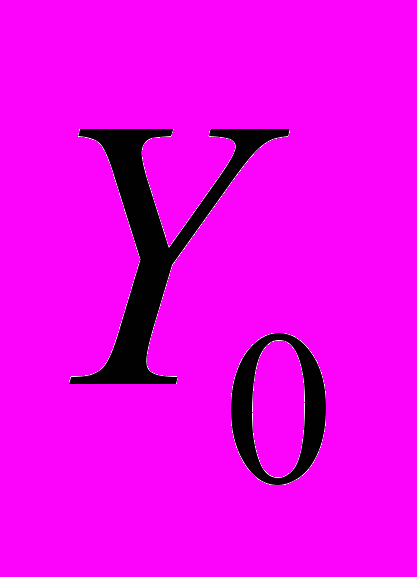 соответствует последовательности ранжированных амплитуд относительных флуктуаций акустического шума в день выхода роя, числовые характеристики которой получены статистической обработкой звуковых шумов размножавшихся семей (объем выборки N=30), а Yn – относительные флуктуации акустического шума семьи за n дней до выхода роя (n=20). По мере приближения дня выхода роя в динамике флуктуаций звукового шума, представленной функциональной зависимостью Yn(Yo) относительно эталонного значения Yо, прослеживается увеличение степени корреляции. Коэффициенты a (или тангенс угла наклона соответствующей прямой) и b (начальная отсечка прямой), характеризующие изменение акустического шума пчел в ходе подготовки к размножению, вычислены методом наименьших квадратов.
соответствует последовательности ранжированных амплитуд относительных флуктуаций акустического шума в день выхода роя, числовые характеристики которой получены статистической обработкой звуковых шумов размножавшихся семей (объем выборки N=30), а Yn – относительные флуктуации акустического шума семьи за n дней до выхода роя (n=20). По мере приближения дня выхода роя в динамике флуктуаций звукового шума, представленной функциональной зависимостью Yn(Yo) относительно эталонного значения Yо, прослеживается увеличение степени корреляции. Коэффициенты a (или тангенс угла наклона соответствующей прямой) и b (начальная отсечка прямой), характеризующие изменение акустического шума пчел в ходе подготовки к размножению, вычислены методом наименьших квадратов. Что касается надежности анализа, то ее можно оценить по относительной величине тангенса угла наклона прямой и величине отсечки по отношению к ПРАОФ, отнесенного к эталонным значениям относительных флуктуаций Yо (рис. 10б). Поскольку функции
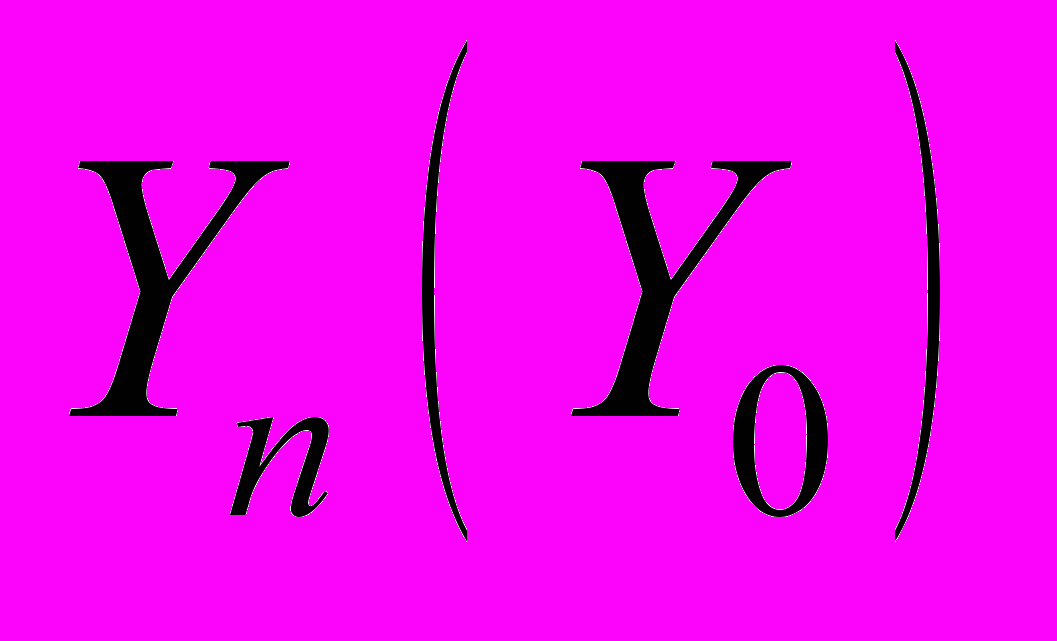 различны и ФОС для трендов и относительных флуктуаций шума также имеют различные количественные характеристики, то их можно использовать для идентификации и количественной дифференциации подготовки пчелиных семей к размножению. Начальные фазы этого процесса четко прослеживаются по значительному увеличению угла наклона прямой, значение параметра a которого начинает превышать единицу.
различны и ФОС для трендов и относительных флуктуаций шума также имеют различные количественные характеристики, то их можно использовать для идентификации и количественной дифференциации подготовки пчелиных семей к размножению. Начальные фазы этого процесса четко прослеживаются по значительному увеличению угла наклона прямой, значение параметра a которого начинает превышать единицу.Динамика изменения параметра a такова, что с приближением дня выхода роя, он стремится к единице. За пять дней до выхода роя среднее значение тангенса угла наклона прямых равнялось 1,12±0,04, за три – 1,04±0,02, а за сутки – 1,01±0,01. В акустических процессах всех восьми семей, завершивших подготовку к размножению вылетом роев, величина отсечки В (нормированное значение ординаты прямой в нулевой точке) в течение пяти дней перед вылетом роя не превышала 0,02±0,01.
Сравнение относительных флуктуаций с помощью функций обобщенного среднего ФОС в пространстве дробных моментов позволяет найти полосу корреляций
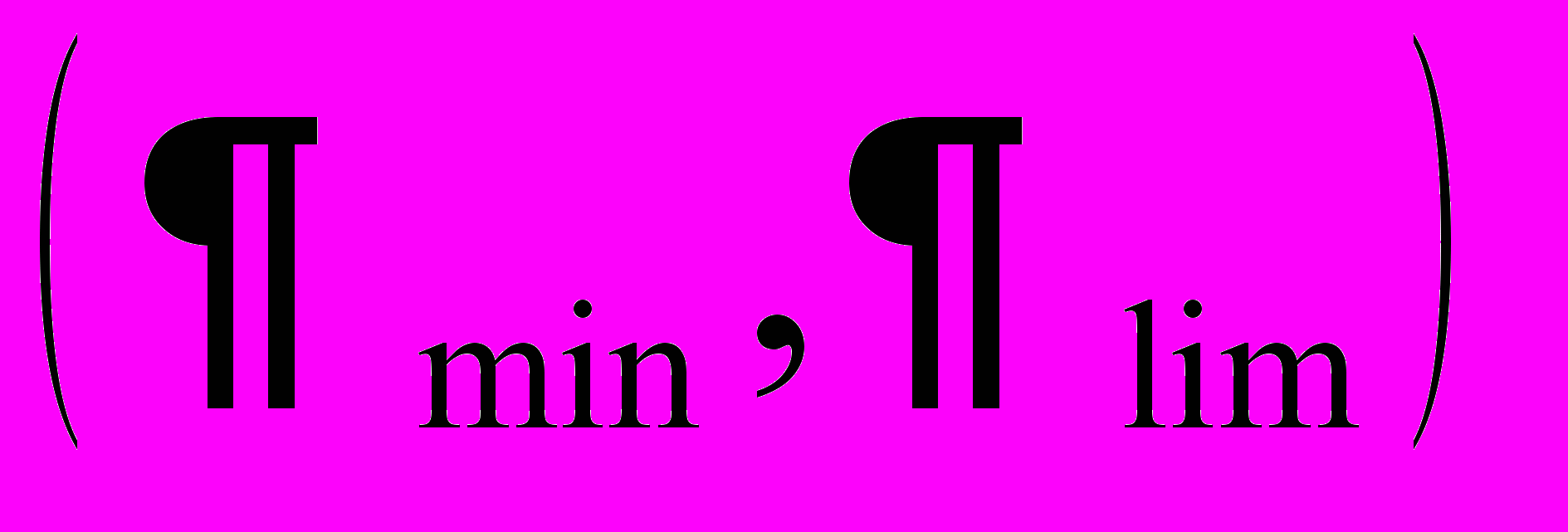 , характеризующую близость отделения части пчел от основной семьи. Чем ближе день выхода роя, тем больше значение величины
, характеризующую близость отделения части пчел от основной семьи. Чем ближе день выхода роя, тем больше значение величины 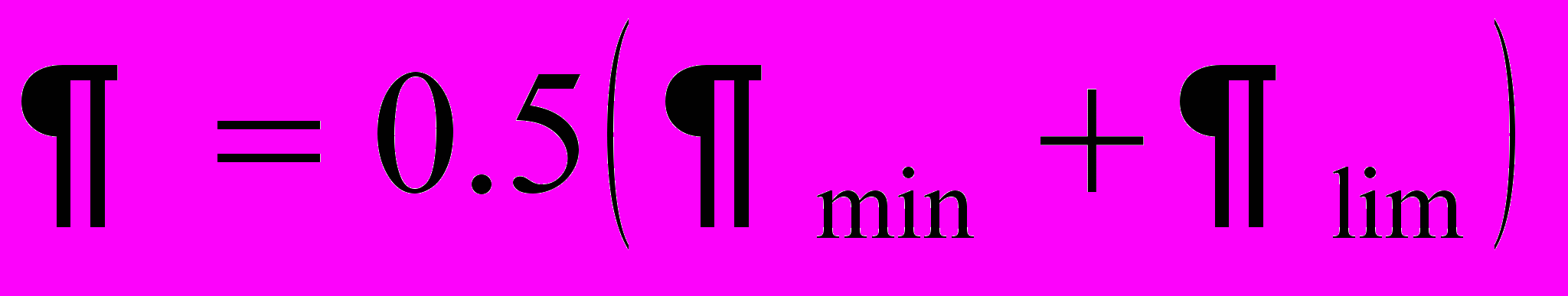 стремится к единице.
стремится к единице. По четырем параметрам кластеров построена динамическая модель, отражающая изменение акустических процессов, сопутствующих подготовке семьи к размножению. Для визуализации и количественного сравнения изменений параметров статистически однородных участков акустического шума в этот период по отношению к дню разделения семьи построен эллиптический классификатор
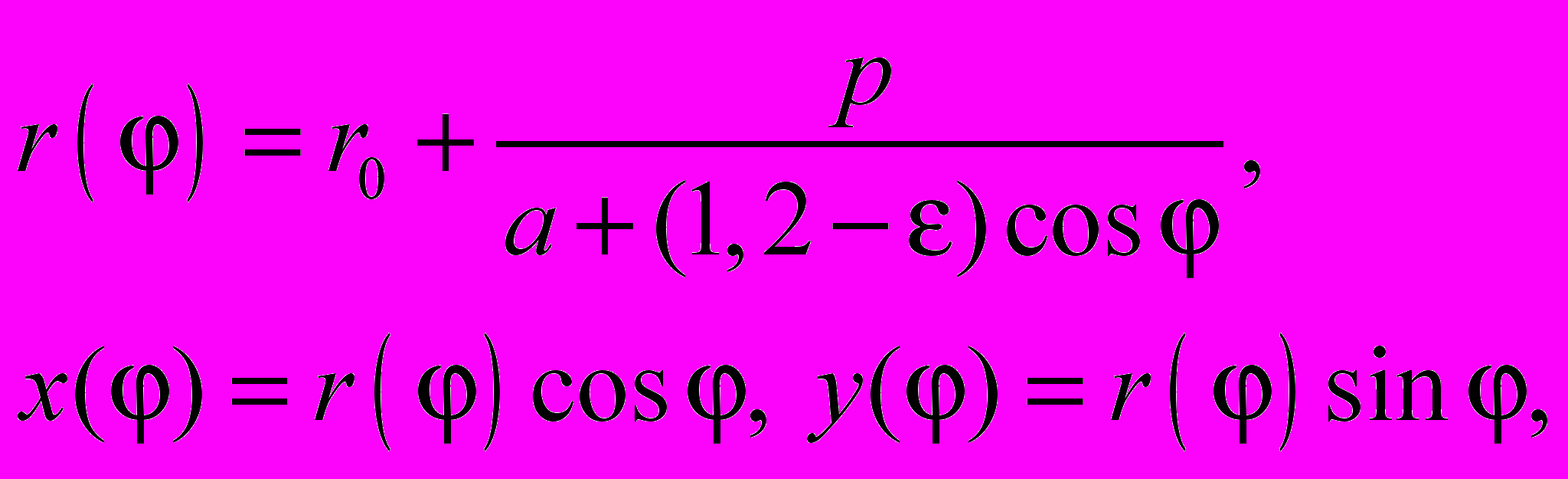 (2)
(2)в котором значение r0 характеризует относительную ошибку подгонки прямой, а e - класс корреляций ОФКП. Величина p равна тангенсу угла наклона при сравнении относительных флуктуаций в системе координат ФОСn-ФОС0 (или Yn-Yо). Значение a, определяющее отсечку прямой на оси ординат, выбрано так, чтобы при прохождении прямой
через начало координат а=1. Акустические процессы в гнездах зимующих пчел
Структура звуков, генерируемых зимующими семьями. В зимний период интенсивные составляющие общего акустического шума пчелиной семьи находятся в диапазоне 100-600 Гц и имеют два отчетливых максимума в областях 150-250 и 350-450 Гц. Первый диапазон порождается звуками, возникающими при вентилировании улья, второй – соответствует работе пчел по обогреву гнезда. Предложенный алгоритм обработки акустических шумов пчел, выраженный в терминах ПРАОФ и ФОС, позволял выявлять и классифицировать внутреннюю структуру сравнительно коротких фрагментов, что необходимо для определения динамики изменения физиологического состояния пчелиных семей в течение зимовки. Статистически близкие реализации для относительных флуктуаций звукового шума пчелиной семьи в течение пяти месяцев (декабрь-апрель) говорили о благополучной зимовке. В ходе анализа флуктуаций акустического шума в течение зимовки эталонному значению
соответствовали флуктуации акустического шума в начальный месяц зимовки (выбран декабрь), а Ym – остальным четырем: январь-апрель (m=1,2,3,4). Так для января и февраля коэффициент a соответственно равен 1,03 и 0,95, а b≈0. Появление расплода в семье приводит к изменению высокочастотной составляющей шума пчел, что сказывается на уменьшении тангенса угла наклона прямой (3) до 0,84 в марте и 0,79 в апреле. Сильную коррелированность соответствующих ПРАОФ между собой, следует важный предварительный вывод о возможности предсказать ход зимовки, если известны относительные флуктуации для его первого месяца. В частности, если она проходит без значительных колебаний внешних климатических параметров (зимовник), то в качестве можно выбрать эталонные значения, полученные в результате многолетних наблюдений, остается учесть ее конкретные особенности подготовки семей к зимнему периоду. Кроме того, если
является последовательностью ранжированных амплитуд с известными числовыми характеристиками (например, акустический шум благополучно зимующей семьи или семьи с известными исходными физиологическим состоянием), а 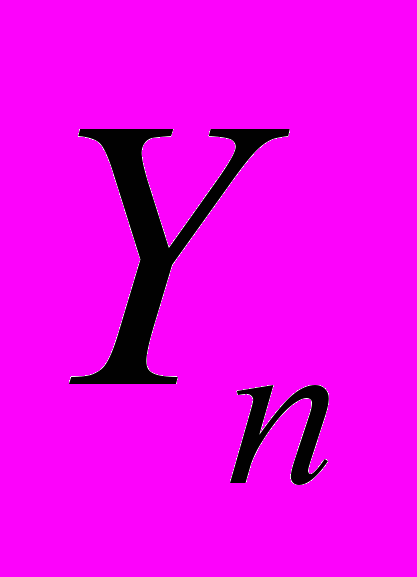 - исследуемая последовательность, то зависимость
- исследуемая последовательность, то зависимость 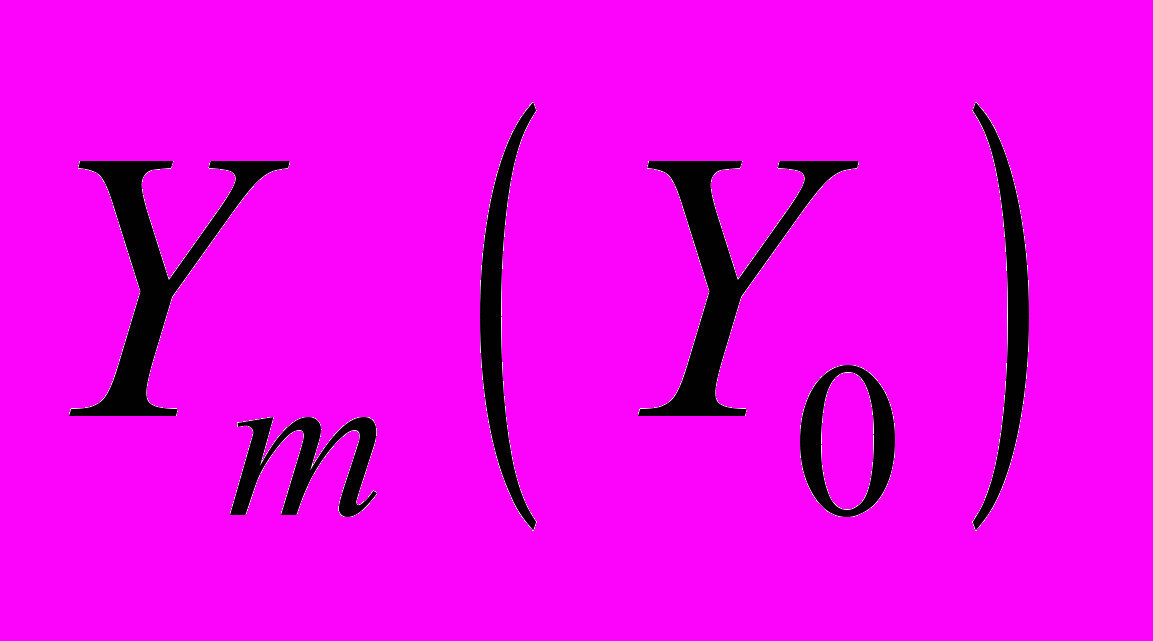 позволяет их сравнить. Для каждой пчелиной семьи отношение различно, поэтому может быть использованы для идентификации и количественного отделения состояния одной от другой. Степень предсказуемости можно оценить по относительной величине тангенса угла наклона прямой и величине отсечки по отношению к ПРАОФ, отнесенной к благополучно зимующей семье. Так, сильная активизация пчел, вызванная повышением влажности в улье, приводит к уменьшению угла наклона зависимости , а для голодающих семей, наоборот, наклон возрастает. Для благополучно зимующих семей временная структура выбранных участков акустического шума практически одинакова и параметр а не выходит за пределы интервала (1,10; 0,85).
позволяет их сравнить. Для каждой пчелиной семьи отношение различно, поэтому может быть использованы для идентификации и количественного отделения состояния одной от другой. Степень предсказуемости можно оценить по относительной величине тангенса угла наклона прямой и величине отсечки по отношению к ПРАОФ, отнесенной к благополучно зимующей семье. Так, сильная активизация пчел, вызванная повышением влажности в улье, приводит к уменьшению угла наклона зависимости , а для голодающих семей, наоборот, наклон возрастает. Для благополучно зимующих семей временная структура выбранных участков акустического шума практически одинакова и параметр а не выходит за пределы интервала (1,10; 0,85). Дополнительную информацию о состоянии зимующих семей можно получить, анализируя сглаженные средние значения (тренды) предварительно проинтегрированных участков шума. Для каждой они различны и поэтому также могут быть использованы для отделения показателей одной семьи от другой (рис. 14). При благоприятных условиях зимовки тренды представляют собой линии, практически совпадающие с осью абсцисс (кривые 0 и 1). Значительные отклонения от них могут быть обусловлены различными возбуждениями пчел (кривая 2) или недостаточной обеспеченностью кормовыми запасами (кривая 3). Тренды сравниваются с помощью функции обобщенных средних, учитывающих их парные корреляции в пространстве дробных моментов.
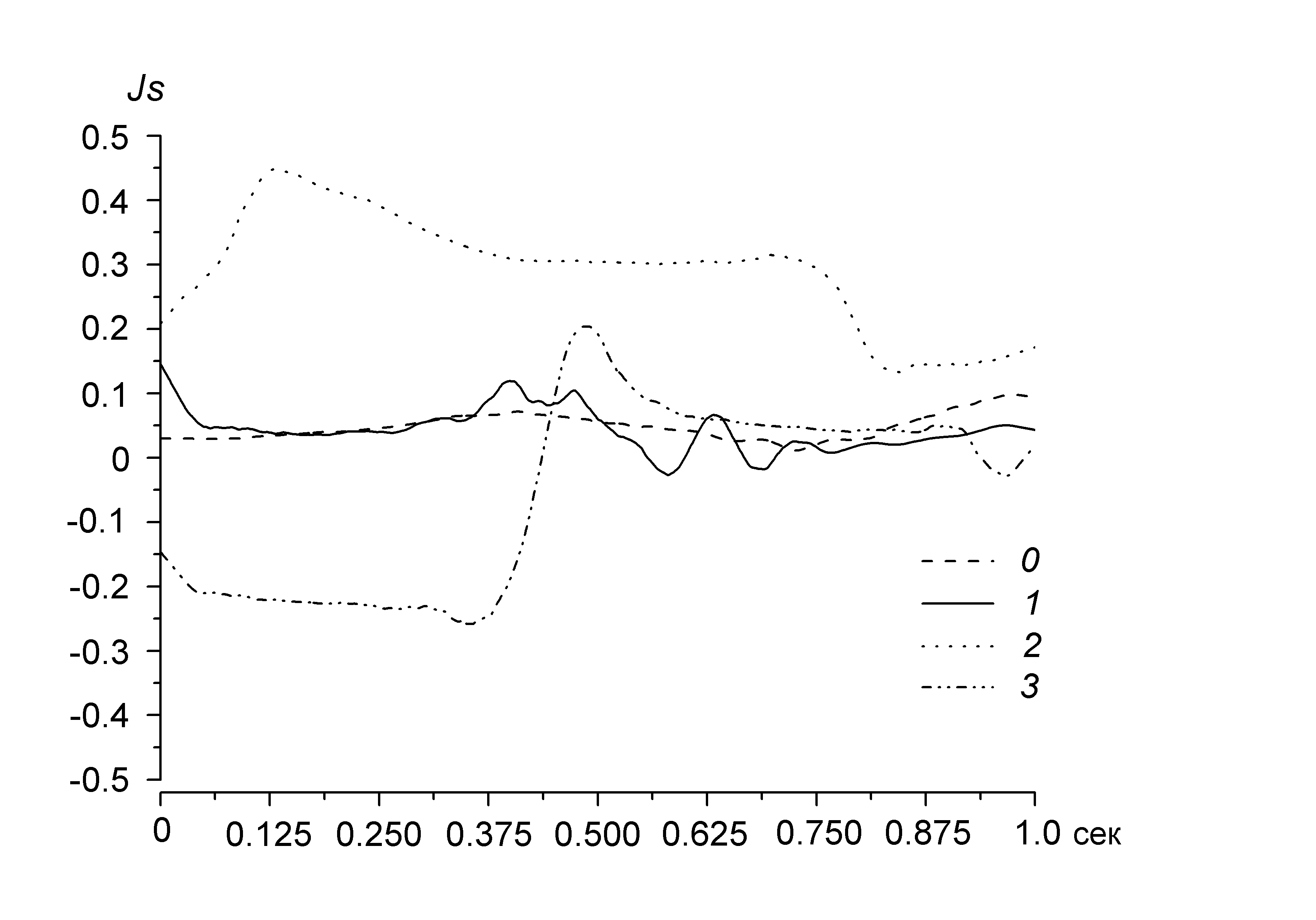
Рис. 14. Сглаженные средние значения (Js – тренды) акустического шума четырех зимующих семей, полученные в результате численного интегрирования участков длительностью 1 с: 0, 1 — благоприятные условия зимовки; 2 — возбуждение; 3 — голодание
Разработанный нами метод дает возможность использовать динамику сильно-флуктуирующих величин общего звуковой шума пчелиной семьи для определения соответствия ее физиологическим потребностям внутригнездовой температуры, концентрации углекислого газа и других показателей микроклимата гнезда.
Акустические отклики на вибрационную стимуляцию. Исследования выполнены в период зимовки пчелиных семей, отличавшихся по физиологическому состоянию. На вибрационный импульс пчелы реагировали резким повышением интенсивности генерируемых звуков, которые можно принять в качестве акустического отклика. Его временная структура связана с физиологическим состоянием пчелиных семей. От него зависят крутизна фронта акустического отклика и его последующее развитие. Семьи, находящиеся в нормальном состоянии (благополучно преодолевающие зимовку), реагируют на одиночный вибрационный импульс резким усилением генерируемых звуков. Их интенсивность за 5-10 мс возрастает в 1,6 ±0,5 раза, затем в течение 4,5 ± 0,4 с уменьшается по отношению к исходному уровню в 1,3 ±0,1 раза. Продолжительность этого периода в звуковом отклике составляет 3,8 ± 0,6 с. За этим следуют некоторое усиление звуков, которое чаще всего не достигает исходного уровня, и некоторое изменение их частоты. Общая продолжительность акустического отклика, изменяющегося по амплитуде и частоте, составляет 8,2 ± 0,3с.
Отклонение состояния зимующих семей от нормы отражается на структуре акустического отклика. Семьи, испытывающие голодание, связанное с недостатком кормовых запасов, реагируют на вибрационную стимуляцию увеличением продолжительности и уменьшением фронта нарастания интенсивности акустического отклика по сравнению с его изменением у семей, находящихся в нормальном состоянии. Сходную структуру имеет акустический отклик у семей, в гнездах которых имеются отклонения от нормы температуры и/или влажности воздуха. Семьи, зимующие в ульях с повышенной температурой, слабо или совсем не реагируют изменением генерируемых звуков на вибрационную стимуляцию. Это связано с исходно высоким уровнем возбуждения пчел, обусловленного неблагоприятным воздействием термофактора, независимо от его природы. Она может быть связана с повышением внешней температуры за пределы оптимального диапазона для зимовки или активизацией тепловыделения самими пчелами, что чаще всего связано с развитием инфекций или инвазий.
Структура акустических ответов на вибрационную стимуляцию, зависящая от состояния пчелиных семей, четко дифференцируется по близости функций обобщенного среднего. Вычисление ФОС для различных участков акустического ответа позволяет получить статистический кластер параметров, образующих семейство однородных его значений. В частности, сравнение в терминах ФОС участков исходного звука, генерируемого пчелами, и его изменений под действием вибрационной стимуляции можно провести по двум параметрам: тангенсу угла наклона прямых и отсечке прямых на оси ординат. В случае статистической близости этих участков (схожести их временной структуры) функции обобщенного среднего, представленные друг относительно друга на координатной плоскости, образуют отрезок прямой линии:
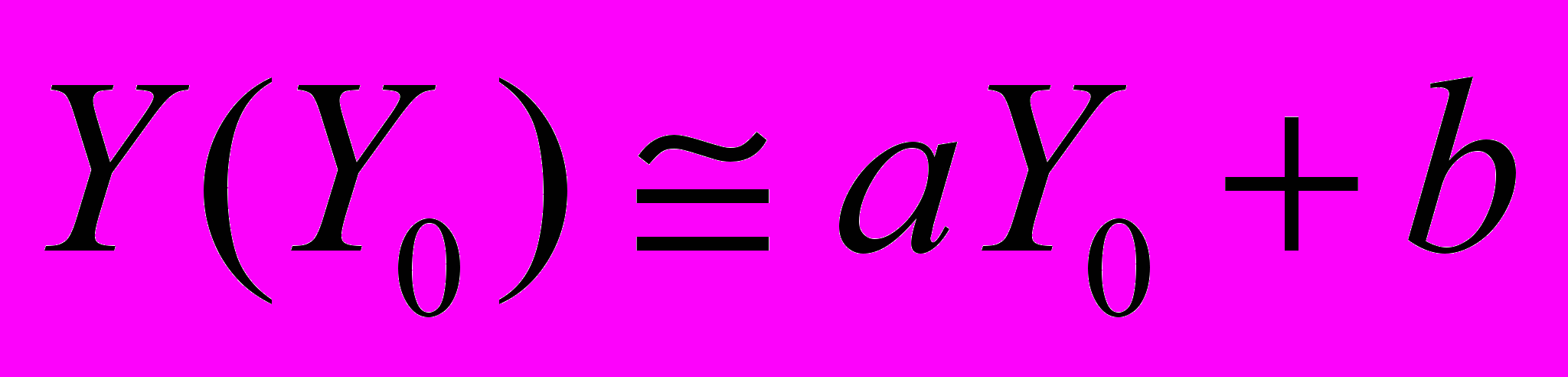 , где соответствует флуктуациям исходной структуры звуков, а Y – после вибрационной стимуляции. Коэффициенты a (или тангенс угла наклона соответствующей прямой) и b (начальная отсечка прямой), определенные методом наименьших квадратов, характеризуют изменение акустических процессов. При совпадении их временных структур перед и после воздействия стимула первый коэффициент равняется единице, второй – нулю. Эти параметры у нормально зимующих семей варьируют в незначительных пределах. Так, значения a меняются в пределах 1,05…0,84, а отсечка прямых близка к нулю b0. Минимальных значений корреляционные связи достигают через 3-5 с после вибрационной стимуляции пчел в то время, когда интенсивность звуков оказывается ниже их исходных уровней. Флуктуации акустических ответов пчел на вибрационную стимуляцию приближаются к минимуму при неблагоприятной для них внутриульевой температуре и влажности воздуха. В таких ситуациях значения a варьируют от 1,1 до 0,45, а отсечка прямых – от 0 до 0,03.
, где соответствует флуктуациям исходной структуры звуков, а Y – после вибрационной стимуляции. Коэффициенты a (или тангенс угла наклона соответствующей прямой) и b (начальная отсечка прямой), определенные методом наименьших квадратов, характеризуют изменение акустических процессов. При совпадении их временных структур перед и после воздействия стимула первый коэффициент равняется единице, второй – нулю. Эти параметры у нормально зимующих семей варьируют в незначительных пределах. Так, значения a меняются в пределах 1,05…0,84, а отсечка прямых близка к нулю b0. Минимальных значений корреляционные связи достигают через 3-5 с после вибрационной стимуляции пчел в то время, когда интенсивность звуков оказывается ниже их исходных уровней. Флуктуации акустических ответов пчел на вибрационную стимуляцию приближаются к минимуму при неблагоприятной для них внутриульевой температуре и влажности воздуха. В таких ситуациях значения a варьируют от 1,1 до 0,45, а отсечка прямых – от 0 до 0,03.Использование функций обобщенного среднего для выделенных статистически однородных участков акустических процессов позволяет прогнозировать ход зимовки пчелиных семей, для чего необходимо ФОС нормально зимующих семей. Исходя из того, что функции
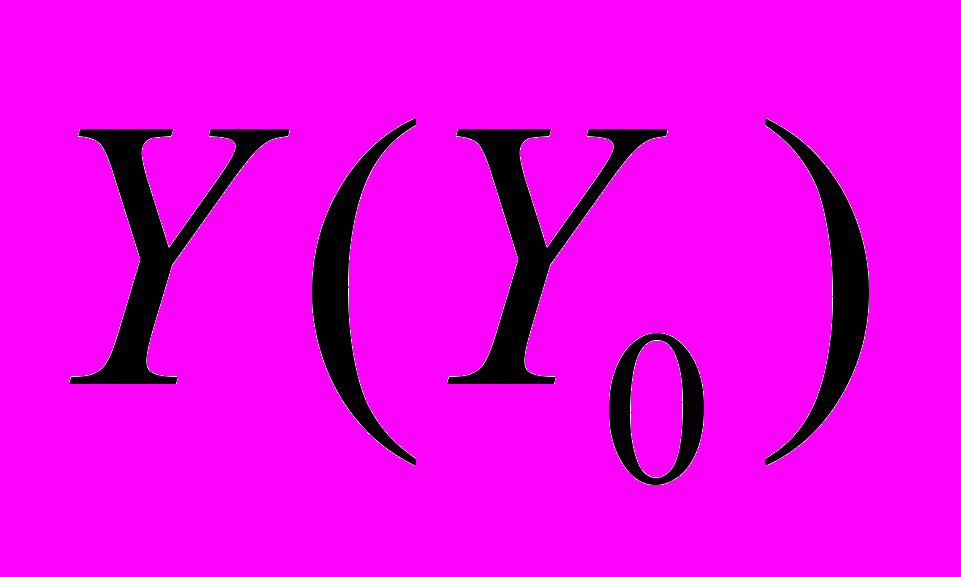 различны, то их можно использовать для идентификации и количественной дифференциации пчелиных семей по их физиологическому состоянию. Сильная корреляция прослеживается при сопоставлении зависимостей флуктуаций акустических откликов семей, испытывающих недостаток корма, и нормально зимующих семей. Различия между этими семьями выражаются в динамике изменений флуктуаций звуковых откликов на вибрационные импульсы.
различны, то их можно использовать для идентификации и количественной дифференциации пчелиных семей по их физиологическому состоянию. Сильная корреляция прослеживается при сопоставлении зависимостей флуктуаций акустических откликов семей, испытывающих недостаток корма, и нормально зимующих семей. Различия между этими семьями выражаются в динамике изменений флуктуаций звуковых откликов на вибрационные импульсы. Значительному увеличению интенсивности акустического отклика у семей, испытывающих недостаток корма, сопутствует изменение его временной структуры. Эти изменения можно анализировать по коэффициентам a и b. Примерно через 30 с после возбуждения корреляция относительно нормально зимующей семьи значительно уменьшается (а=0,31, b=0,03). В ульях с повышенной влажностью, наоборот, значение коэффициента а увеличивается от 0,54 до 0,76.
Анализ частотных спектров выделенных участков акустического шума до и после воздействия стимула позволяет получить дополнительную информацию о физиологическом состоянии пчел. Несмотря на то, что спектр акустического шума является нестационарным, предварительное выделение статистически однородных участков в шуме и использование процедуры оптимального сглаживания их частотных спектров позволяет анализировать динамику изменения основных пиков традиционным спектроскопическим методом (по интенсивности и полуширине). Интенсивные составляющие спектра до и после вибрационного стимула находятся в диапазоне 100-450 Гц с двумя максимумами на частотах около 200 и 400 Гц. При этом возбуждение пчел незначительно изменяет положение максимумов в спектре, приводя лишь к уширению полуширины. К такому же результату приводят и сравнения ФОС для сглаженных спектров выделенных участков. Интересно отметить, что при нормальных условиях зимовки интенсивность звукового шума пчел после возбуждения всегда меньше первоначального.
Акустические отклики пчел на электромагнитные воздействия
Изменения амплитуд частотных составляющих акустических процессов в пчелиных семьях указывают на зависимость их физиологического состояния от суточной динамики нагрузки ЛЭП. Интенсивные составляющие частотных спектров получены после выделения статистически однородных участков для каждого выделенного участка шума. В утренние часы интенсивные составляющие в спектрах звуков, генерируемых пчелами, находятся в диапазоне 220-250 Гц. В дневное время возмущения электромагнитных полей (ЭМП) приводят к увеличению интенсивных составляющих на частотах 400-450 Гц. Этому же сопутствует увеличение летной активности пчел. В вечернее время возрастает интенсивность частотных составляющих в диапазоне 300-350 Гц.
Использование функций обобщенного среднего для анализа шумов, генерируемых пчелами, позволило выявить статистически значимое влияние ЭМП на физиологическое состояние пчел. На это указывает наличие связи между флуктуациями шумов пчелиных семей и изменениями нагрузки трассы. Сравниваемые интегрированные участки акустического шума сравнивались по близости их функций обобщенного среднего к отрезкам прямых линий. Количественная характеристика этой близости оценивается по четырем параметрам: D, Tg, B,
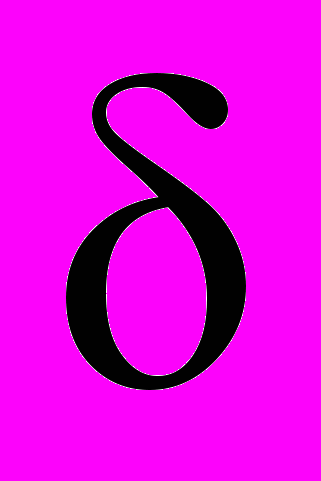 ,
, Детализация анализа достигается путем количественного сравнения параметров фитинга последовательности ранжированных амплитуд относительных флуктуаций звукового шума семей в течение суток. Значения параметров фитинга степенной зависимостью для них приведены в таблице 2. Показатели в последней графе иллюстрируют качество подгонки гипотезы
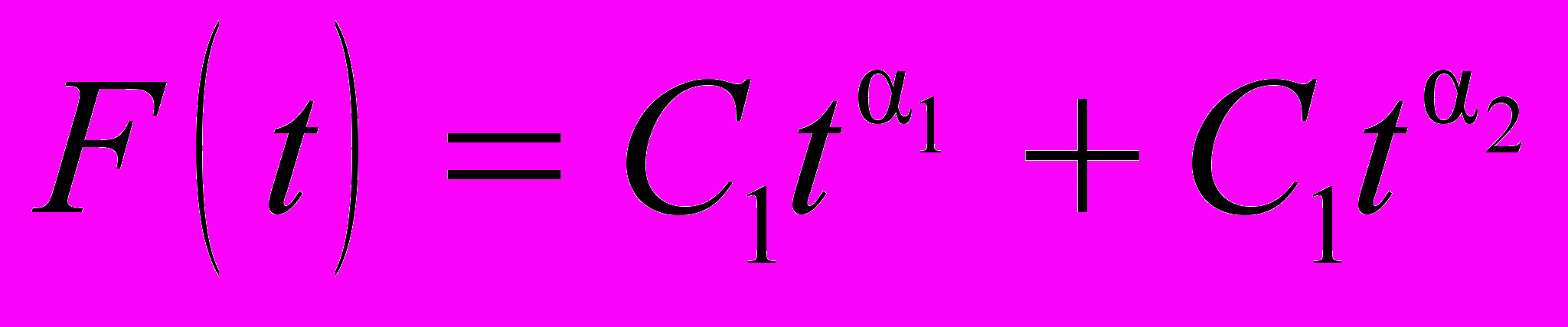 для заданной ПРАОФ. Относительная ошибка (δ) вычисляется как отношение стандартного отклонения между ПРАОФ и подгоночной функцией к величине среднего значения подгоняемой функции
для заданной ПРАОФ. Относительная ошибка (δ) вычисляется как отношение стандартного отклонения между ПРАОФ и подгоночной функцией к величине среднего значения подгоняемой функции Таблица 2. Подгоночные параметры, полученные для ПРАОФ, описывающие ненулевые относительные флуктуации звукового шума пчелиной семьи при разных значениях нагрузки трассы.
-
Время
суток
C1
α1
C2
α2
δ
Утро
-0,045
-0,985
0,756
-0,345
0,034
День
-0,012
-0,873
0,853
-0,412
0,041
Вечер
-0,53
-0,768
0,711
-0,562
0,021
В таблице приведены наиболее различимые подгоночные параметры. Они могут быть использованы в качестве декодирующих ключей для изучения малых воздействий ЭМП на физиологическое состояние пчелиной семьи.