
Экобиологические механизмы акустического и теплового взаимодействия пчел 03. 02. 08 Экология
| Вид материала | Автореферат |
- Перечислить формы и механизмы взаимодействия национальных культурных центров между, 87.24kb.
- Перечислить формы и механизмы взаимодействия национальных культурных центров между, 37.75kb.
- Докладе рассматриваются две задачи о распространении тепловой волны в образце цилиндрической, 9.26kb.
- План I. Введение. II. Цель и задачи работы. III. Происхождение антибиотиков в пчелином, 649.46kb.
- М. В. Лупанов описывает ульи собственной конструкции с квадратными рамками, которые, 726.39kb.
- «Биология и патология пчел» Специальность 111201 Ветеринария Пояснительная записка, 157.25kb.
- Сессия Американского Акустического Общества, г. Провиденс, 2006, международный, 1000,, 65.95kb.
- Эколого-Токсикологические основы системы защиты медоносных пчел от болезней и вредителей, 608.83kb.
- Эколого-Токсикологические основы системы защиты медоносных пчел от болезней и вредителей, 609.24kb.
- Ветеринарные требования при импорте в российскую федерацию медоносных пчел, шмелей, 49.98kb.
Безрасплодный период в жизни пчел
Толерантность пчел к охлаждению. В отличие от одиночных насекомых, медоносные пчелы, не обладая способностью диапаузировать, вынуждены использовать как индивидуальные, так и социальные этолого-физиологические адаптации. Но основная роль в противостоянии к низким температурам отводится поведенческим адаптациям – образованию холодовой агрегации и поддержанию внутри скопления высокой положительной температуры. Прежде всего, это связано с отсутствием у отдельных пчел физиологических приспособлений к низким температурам. Основные показатели холодоустойчивости пчел – температура оцепенения, температура максимального переохлаждения – зимой и летом сохраняются почти на постоянном уровне.
Связь между устойчивостью к охлаждению и диэлектрической проницаемостью пчел. Установлено, что температура оцепенения пчел летней и осенней генераций не имеет существенных различий, а замерзание жидких фракций тела подвержено возрастной и сезонной изменчивости. Значения температур оцепенения и замерзания определяли по скачкам диэлектрической проницаемости. Температурные пороги холодового оцепенения у пчел весенне-летних генераций находилась на уровне 12,9±0,4оС. Продолжительность жизни этих пчел ограничивается обычно 30-40 сутками. Зимуют и доживают до весны пчелы осенних генераций. У них холодовое оцепенение наступало при температуре 13,3±0,3оС. Сходную тенденцию повышения устойчивости к охлаждению имело температура максимального переохлаждения (ТМП). Пчелы весенне-летних генераций замерзали в среднем при –5,1±2,2, а осенних, способных перезимовывать, выдерживали охлаждение от –7 до –15оС. В течение зимовки происходило некоторое понижение устойчивости к замерзанию. От ноября к январю ТМП повышалось на 1,5-2оС.
Охлаждению пчел от +20 до –25оС сопутствовало неравномерное изменение времени релаксации дипольных моментов, что обуславливает неравномерность динамики диэлектрической проницаемости. Понижение температуры до состояния холодового оцепенения сопровождалось повышением диэлектрической проницаемости тела пчел. Значения действительной части диэлектрической проницаемости (ε') зависели от сезонной и возрастной изменчивости физиологического состояния пчел. В фазе оцепенения пчел летних генераций ε' составляло 54,2±2,4, у осенних – 24,1±1,9, у зимних – 25,2±3,7 (рис. 4).

Рис. 4. Температурная зависимость диэлектрической проницаемости тела пчел в июне (1), ноябре (2) и январе (3)
Дальнейшему охлаждению пчел сопутствовало вначале некоторое понижение значения действительной части диэлектрической проницаемости. Динамика этого процесса нарушалась при охлаждении, приближающемся к началу замерзания жидких фракций тела пчелы. В фазе кристаллизации значение ε' возрастало. У пчел летних генераций ε' достигало 4,3±1,6, у осенних – 20,2±1,1, у зимних – 20,3±0,7 (рис. 4).
Флуктуации внутригнездовой температуры. В течение зимовки сильное влияние на внутригнездовую температуру оказывают колебания внешней температуры. Но зависимость внутригнездовой температуры от внешней уменьшается от начала к завершению зимовки. Так, в ноябре-декабре при изменении внешней температуры от +11 до –15 оС флуктуации температуры теплового центра семьи, содержавшей около 20 тыс. пчел, изменялись в пределах 2,5…2,7 оС. В феврале-марте изменениям внешней температуры от +1 до –17 оС колебания разогрева теплового центра не превышали 1…1,6оС. В соответствии с этим коэффициент корреляции между внешней температурой, варьирующей в пределах
–17…+11оС, и разогревом теплового центра уменьшался с
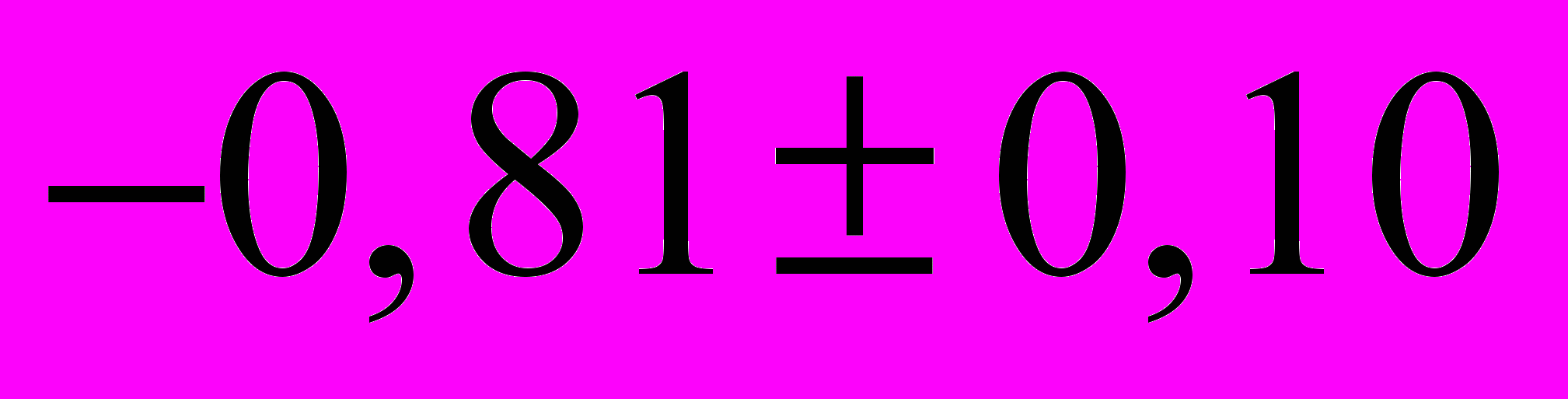 , до
, до  . Но в течение всей зимовки его значение между температурой теплового центра и расположенной над ним периферией скопления пчел поддерживался на уровне 0,82±0,11 (Р>0,95).
. Но в течение всей зимовки его значение между температурой теплового центра и расположенной над ним периферией скопления пчел поддерживался на уровне 0,82±0,11 (Р>0,95).Связь между колебаниями внешней и максимальной внутригнездовой температурой зависит от численности пчел в семье. В семьях с относительно небольшой численностью пчел (≈10 тыс.) сильная отрицательная корреляция между анализируемыми температурами прослеживается только в начале периода зимовки на начальных фазах адаптации к наступившему осеннему похолоданию. С декабря, значение коэффициента корреляции уменьшается до –0,36±0,12. В семьях, содержавших примерно вдвое большее число пчел (≈18 тыс.), колебания температуры в ноябре-декабре достигали 0,8.…2,7оС, а коэффициент корреляции находился на уровне –0,84±0,09. Во второй половине зимовки (с января) семьи почти не изменяли терморегуляторную активность даже на значительные колебания внешней температуры.
Независимо от численности, пчелы реагируют на охлаждение уменьшением поверхности скопления (занимаемого объема) за счет уплотнения, достигающего максимума в зоне соприкосновения с холодным воздухом. При этом потоки тепла в разных направлениях отличаются из-за образующихся неоднородностей структуры скопления. Суммарный поток тепла с его поверхности зависит от эффективной внешней поверхности, т.е. от площади, через которую отдается тепло. Она всегда меньше геометрической поверхности, а ее величина существенно зависит от внешней температуры и локализации пчел в разных межсотовых пространствах и расстояния от леткового отверстия. Через него происходит в основном теплообмен между внутриульевым пространством и внешней средой.
При определенных значениях внешней температуры (зона оптимальности –3,0…+8,8 оС) мощность теплового потока приходящего на 1 см2 поверхности слабо зависит от количества пчел в скоплении. В семьях, содержащих от 8,2 до 19,5 тыс. пчел тепловые потоки находятся в пределах 2,05…3,2110-3 Вт/см2. При этом величина теплового потока зависит от его направления. В пределах указанного диапазона она больше к верхней поверхности и меньше к нижней. В зоне оптимальности не прослеживается существенной вариабельности температуры в тепловом центре семей, что обуславливается сходной тепловой нагрузки на каждую из пчел, среднее значение которой ограничивается 6…8 10-4 Вт.
За пределами зоны оптимальности вариабельность разогрева теплового центра возрастает по мере уклонения от нее и зависит от численности пчел в скоплениях. При прочих равных условиях уменьшению их численности сопутствует возрастание дестабилизации тепловых процессов, что приводит к увеличению теплопродукции за счет дополнительного потребления корма. В частности, мощность основного обмена в зоне оптимальности семьи, содержащей примерно 8 тыс. пчел, составляет в среднем 3,28 Вт (47 кал/мин). Под влиянием охлаждения до –15оС в течение 120 мин. теплопродукция возрастает в 2,6 раза, увеличивая тепловую нагрузку на пчелу от 610-4 до 1,1 10-3 Вт.
Температурные градиенты в направлении от теплового центра к верхней и нижней поверхностям скопления пчел существенно различаются по абсолютным значениям и вариабельности. К верху они меняется незначительно, не превышая в течение зимовки 12оС, а к низу, постоянно изменяясь, достигают 24…28оС. На таком же уровне при –20…–18оС поддерживается разность между внешней температурой и разогревом тела пчел, образующих нижнюю часть их скопления.
Ритмичность терморегуляторной активности у пчел. Проведен анализ ритмичности терморегуляторной активности у пчел в разные периоды годового цикла их жизни. Показано, что доминирующую роль в синхронизации циркадных ритмов и консолидации пчел в единую биосистему играет тепловой шум (флуктуации температуры внутри скоплений пчел). Он формируется в процессе взаимодействия между отдельными особями.
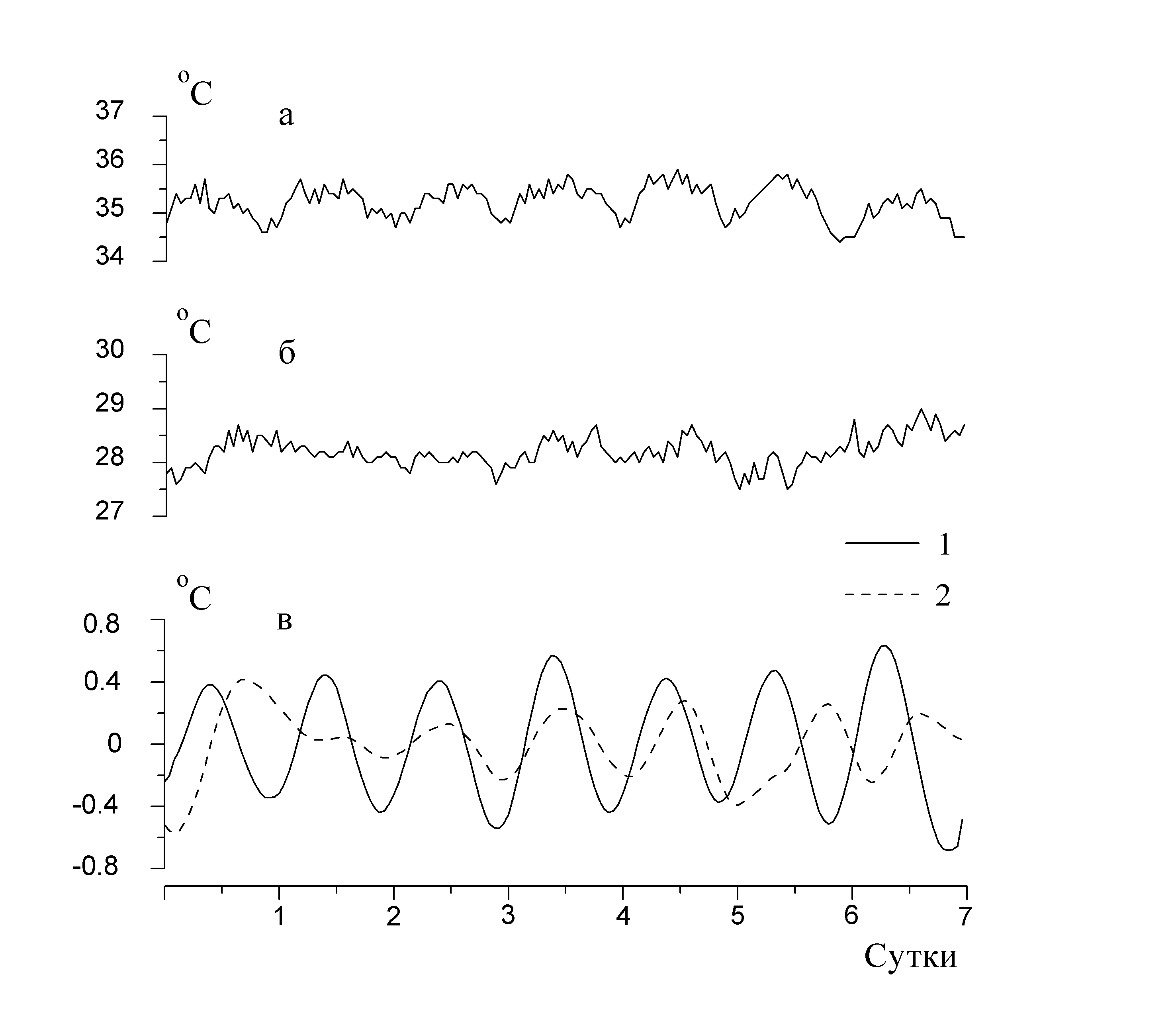
Рис. 5. Недельная динамика изменения температуры в тепловом центре и периоды активности в разное время жизнедеятельности пчелиной семьи: а – лето; б – зима; в – крупномасштабные флуктуации температуры (1 – лето, 2 – зима)
Недельная динамика изменения температуры в зоне расплода показана на рис. 5. В летний период (рис. 5а) четко прослеживается активизация пчел днем, что выражается в увеличении температуры в гнезде, а ночью она уменьшается. В этом выражается ритмичность периодов активности и пассивности пчел. Максимумы периодов соответствуют наибольшей активности пчел, связанной с доставкой в улей нектара, пыльцы или воды. В ходе зимовки суточная ритмичность может нарушиться в результате резких перепадов внешней температуры, изменений физиологического состояния пчел и др. На рис. 5б представлены данные недельной динамики изменения температуры в семье после постановки пчел в закрытое помещение (омшаник), в котором на постоянном уровне поддерживаются освещенность и температура. По ним трудно выявить наличие циркадных или иных периодических компонент.
Анализ суточной динамики внутригнездовой температуры методом дробных моментов указывает на наличие ее крупномасштабных флуктуаций. По характеру изменения этих флуктуаций можно прогнозировать восстановление прежних и появление новых периодических составляющих (Рис. 5в). Семья, помещенная в омшаник, восстановила циркадные ритмы полностью на 10-й день, но произошел фазовый сдвиг пиков активности из-за новых условий содержания. Величина сдвига составляла 0,5…1,5 ч и зависела, прежде всего, от численности пчел в семье.
Время, необходимое для синхронизации ритмов терморегуляторной активности, зависит от физиологического состояния пчел. При объединении безматочной семьи, находящейся под открытым небом, с примерно такой же по численностью пчел семьей с маткой из омшаника, синхронизация ритмов активности происходила через 18-20 ч. За это время в объединенной семье сформировался новый тепловой режим со «средним» ритмом, и процесс сдвига фаз закончился на уровне отдельных пчел. До объединения фазовый сдвиг терморегуляторной активности между этими семьями составлял 2,5 ч.
Временная структура крупномасштабных флуктуаций терморегуляторной активности изменяется в течение суток и в годовом цикле жизни пчелиной семьи. Эти различия четко прослеживаются в весенне-летний и осенне-зимний сезоны, что, вероятно, связано с сезонной изменчивостью локомоторной активности пчел. Она в зонах умеренного и холодного климата достигает максимума в конце весны и сохраняется на высоком уровне до начала осени, а в течение зимовки – приближается к минимуму. Но от весенне-летнего к осенне-зимнему сезонам продолжительность периодов крупномасштабных флуктуаций температуры возрастает в среднем с 2,15±0,10 до 2,4±0,3 ч. В сходных пределах изменяется их суточная продолжительность. В световое время суток весенне-летнего сезона она составляет 2,3±0,1 ч, в темное – 2,1±0,1, а в течение осенне-зимнего, соответственно, 2,5±0,5 и 2,2±0,3 ч.
Периодичность крупномасштабных колебаний температурной активности пчел не имеет выраженной связи с изменениями внешней температуры. Ее стабилизация также не влияет на установившийся ритм внутригнездовых флуктуаций температуры, что установлено на пчелиных семьях, зимовавших под открытым небом. Их вместе с ульями переносили в помещение, температура в котором поддерживалась на уровне 4…4,4оС. По результатам шестикратной повторности опыта установлено, что адаптации пчел к новым условиям жизни сопутствовал фазовый сдвиг пика терморегуляторной активности. Вначале величина сдвига варьировала в пределах 0,5…1,5 ч, находясь в прямой зависимости от численности пчел в семье. Через сутки, независимо от численности пчел в семьях, различие между фазами терморегуляции по отношению к исходным уровням уменьшалось до 0,3±0,2 ч. Полная нормализация ритмичности температурной активности и ее синхронизация во всех семьях, находившихся в одном помещении на расстоянии 40-50 см друг от друга, происходили через 2-3 суток.
В условиях тепловой изоляции двух семей, отличавшихся по численности пчел, ритмичность крупномасштабных флуктуаций в центрах их скоплений могла не совпадать. В частности, у одной из двух семей продолжительность периодов крупномасштабных флуктуаций температуры равнялась 2,2±0,4 ч, у другой – 2,1±0,1 ч. В первой из них находилось около 18 тыс. пчел, во второй – около 10 тыс. Семьи зимовали в одном улье, разделенном сплошной деревянной перегородкой толщиной 2,0 см. После ее замены на перфорированную перегородку с отверстиями 2,5 мм, исключающей прямой контакт и обмен кормом между пчелами, температурные флуктуации в семьях синхронизировались через 60-72 ч. При этом устанавливался ритм семьи, содержавшей большее количество пчел.
Изменчивость температуры тела пчел. Температура разных частей тела пчел, находящихся на периферии их скопления, зависит от места локализации и внешней температуры. При колебаниях в пределах от – 1 до –13оС, наибольшим разогревом чаще всего отличается грудной отдел тела пчелы. Средние значения его температуры на десятые доли градуса отличаются от температуры головного и брюшного отдела. Но температура головного отдела иногда приближается или немного превышает температуру грудного отдела. В отличие от этого среднее значение температуры брюшного отдела всегда ниже грудного.
Понижение внешней температуры отражается на повышении температуры тела пчел, образующих периферию их скопления. Наибольший разогрев пчел происходит в нижней части их скопления, которая подвергается обычно наиболее интенсивному охлаждению. Так, при понижении внешней температуры от –1 до –21оС температура грудного отдела повышалась в среднем на 3,0оС, головного – на 2,6 оС и брюшного – на 2,4оС (Р≥ 0,99), а в верхней части на 1,5 оС, 0,6 оС и 0,9 оС (Р≥ 0,95) соответственно. Самую высокую температуру, достигавшую 27,9 оС, имели брюшные отделы у некоторых пчел, находившихся в нижней части гнезда при –13оС. В этой же зоне у отдельных пчел температура брюшного отдела опускалась до 7,5 оС.
С понижением внешней температуры возрастает представительство пчел, у которых грудные отделы нагреты сильнее головных. Максимальное различие между разогревом головного и грудного отделами возрастает при понижении внешней температуры. При
–1оС оно достигает 5,7оС, при –13 – 8,5оС и при –21 – 10,3 оС, а доля пчел, у которых температура груди превосходит температуру головы не менее, чем на 2оС, равняется, соответственно, 25,4, 36,0 и 47,2%. Но понижению внешней температуры сопутствует уменьшение на нижней поверхности скопления количества пчел, охлаждающихся до температуры стимулирующей холодовое оцепенение. В частности, при –1оС их доля составляет в среднем 52,4±4,9%, при –13оС – 45,7±4,1 и при –21оС – 32,1±3,3%.
В меньшем диапазоне варьирует температура тела у пчел, локализующихся в зоне теплового центра. В частности, при –20 оС температура головных отделов варьировала у разных пчел (n=158) в пределах от 26,8 до 34,3 оС, грудных – от 24,2 до 36,0 и брюшных – от 24,6 до 33,4, а средние значения их разогрева равнялись 29,8, 30,7 и 29,3 оС соответственно.
КОНСОЛИДИРОВАННЫЕ СКОПЛЕНИЯ ПЧЕЛ
Группы пчел при определенной численности (Еськов Е.К., 1992, 1995; Lindauer M., 1954; Free J. B. and Spenser-Booth G., 1959; Harrison J.M., 1987, и др.) агрегируются частично или полностью. Это позволяет им противостоять неблагоприятному воздействию внешней температуры и поддерживать внутри скопления определенную температуру. Регуляция температуры консолидированными скоплениями пчел основана на использовании скоординированных усилий образующих их рабочих особей. Важная роль в этом принадлежит регуляции теплоотдачи и теплосодержания.
Регуляция теплоотдачи. Как и у гомойотермных животных, отдача тепла скоплениями пчел в окружающее воздушное пространство зависит от его температуры. В определенном интервале температур (в зоне температурной оптимальности), зависящем от численности пчел в образуемом ими скоплении, теплоотдача сохраняется на относительно постоянном минимальном уровне. Зона оптимальности для группы, содержащей 550±6 пчел, ограничивается 8,7…10,4оС, в пределах которых тепловыделение поддерживается в среднем на уровне 0,48 Вт (lim. 0,40 – 0,55). Увеличение численности агрегировавшихся пчел примерно в 7 раз (до 3980±26) отражается на расширении зоны оптимальности и ее смещении на область более низких температур. Ее минимальное значение равняется 5,6 максимальное – 9,4оС. В пределах этого диапазона теплоотдача составляет в среднем 2,23 Вт (lim. 2,02 – 2,40).
За пределами нижнего значения зоны оптимальности происходит постепенное нелинейное увеличение теплоотдачи соответственно понижению окружающей температуры. При неограниченной возможности потребления корма потенциальные возможности повышения теплоотдачи в ответ на охлаждение лимитируются численностью агрегировавшихся насекомых. В частности, максимальная теплопродукция группы из 550 пчел при
–3,7±0,4оС достигает 1,82±0,10 Вт. Группа, содержащая около 4000 особей, в состоянии противостоять охлаждению до –21,8±0,5оС при теплопродукции 6,42±0,14 Вт.
Повышение тепловыделения скопления при понижении температуры за пределы нижней границы зоны оптимальности связано с их стремлением противостоять охлаждению увеличением теплопродукции. На это указывает повышение температуры внутри скопления, особенно в его наиболее разогретой области (тепловом центре). В группе, содержавшей около 550 пчел, за 6 ч охлаждения со средней скоростью 2оС/ч теплоотдача возросла в 3,8 раза, а температура в тепловом центре - на 2,1оС (с 24,9 до 27,0оС). В пересчете на одну пчелу тепловыделение возросло в 2,4 раза. Увеличение численности пчел в группе примерно в 7 раз повлияло на увеличение тепловыделения каждой из них в среднем в 1,2 раза. Температура теплового центра возросла на 1,1оС (от 28,1 до 29,2оС).
За пределами верхней границы зоны оптимальности происходит резкое повышение тепловыделения, что связано с влиянием разогрева на активизацию локомоций насекомых. Верхний предел зоны оптимальности для групп, содержащих около 550, 1250, 2600 и 4000 пчел, ограничивается, соответственно, 10,4оС, 9,9оС, 9,6оС и 9,4оС. Отклонение температуры за указанные ее предельные значения на 1,0оС стимулирует увеличение тепловыделения по отношению к таковому в зоне оптимальности в среднем в 1,2, 1,7, 2,0 и 2,3 раза.
Теплосодержание. Температура, поддерживаемая в разных местах скоплений, может использоваться для определения их теплосодержания. По результатам измерений температуры пятью термодатчиками в скоплениях с разной численностью насекомых установлена ее относительно невысокая вариабельность в тех случаях, когда окружающая температура не выходила за пределы зоны оптимальности. В ее пределах флуктуации разогрева теплового центра не превышали 0,5…0,9оС. С этим связана невысокая изменчивость теплосодержания.
Теплоемкость в скоплении пчел изменятся незначительно и определяется в основном содержанием воды в их теле. Теплоемкость гемолимфы, очевидно, близка к теплоемкости воды и составляет 3,77…3,89 кДж/(кг·оС) или 0,9…0,93 кал/(г·оС). Исходя из этого, можно предположить, что удельная теплоемкость всего скопления увеличивается линейно в пределах 2,51…3,35 кДж/(кг·оС) или 0,6…0,8 кал/(г·оС) в зависимости от увеличения плотности пчел. С учетом этих допущений изменение теплосодержания в скоплении, содержащем около 550 пчел, не превышает 0,05 Вт. Под влиянием их 6-часового охлаждения теплосодержание возрастало на 619 кал, а скорость изменения теплоотдачи – на 0,12 Вт (1,72 кал/мин), что обусловлено низким потенциалом теплогенерации скоплениями малой численности. Для указанных значений теплоотдачи и теплосодержания максимальная теплопродукция составляла 1,94 Вт. Тепловая нагрузка на одну пчелу (общая теплопродукция, поделенная на количество особей в скоплении) при максимальном напряжении терморегуляции достигала 0,003 Вт, а области температурного оптимума - около 0,0009 Вт. Такой уровень теплопродукции не выходит за пределы вариабельности ее значений (минимум 0,0005, максимум 0,0015 Вт), определенных по интенсивности метаболизма у индивидуально изолированных пчел, находящихся в состоянии покоя.
Скопления, образованные относительно большим количеством пчел, имели возможность изменять теплонакопление в более широких пределах и в течение большей продолжительности охлаждения на высоком уровне поддерживать терморегуляторную активность. Так, в скоплении, образованном примерно 4 тыс. пчел, находившихся при температуре, колебания которой ограничивалась пределами оптимального диапазона, скорость изменения теплосодержания не выходила за пределы 0,2…0,3 Вт (2,87…4,31 кал/мин), а средняя тепловая нагрузка на пчелу составляла в среднем 0.0006 Вт. В результате охлаждения, продолжавшегося в течение 15,5 ч, максимальная теплоотдача достигала 2.7 Вт. Исходя из этого общая теплопродукция составила 9,12±0,20 Вт. Среднее же значение тепловой нагрузки на одну пчелу, равнявшей 0,0023 Вт, несущественно отличалось от таковой в скоплениях с относительно небольшой численностью.
Таким образом, даже сравнительно малочисленным скоплениям насекомых, находящимся в зоне температурного оптимума, накопленного тепла и небольшой теплопродукции необходимо и достаточно для поддержания устойчивого теплового состояния. Возможность изменения теплосодержания позволяет пчелам мобильно реагировать на изменения внешней температуры, не увеличивая резко теплопродукцию. Биологически целесообразно пчелам сохранять теплопродукцию на постоянном уровне за счет внутреннего накопления тепла, что достигается их уплотнением. Но повышение плотности агрегирования насекомых, стимулируемое охлаждением в пределах температурного оптимума, не существенно влияет на теплоотдачу. Ее изменение не превышает 0,2 Вт. Примерно на столько же изменяется теплопродукция.
За пределами нижней границы зоны температурного оптимума теплоотдача и теплосодержание находятся в тесной зависимости от численного состава агрегирующихся насекомых. С увеличением их численности возрастают возможности изменения теплосодержания при сохранении определенного оптимального соотношения между теплопродукцией и теплоотдачей. Увеличение численности замедляет относительный прирост теплопродукции, но повышение теплоотдачи не имеет прямой связи с понижением внешней температуры. Лишь превышение ею верхней границы оптимального диапазона стимулирует резкое повышение теплоотдачи, что как отмечалось выше связано с активизацией пчел.
В экстремальных ситуациях при угрозе гипертермии функция системы терморегуляции ограничивается в основном удалением избытка тепла. Терморегуляция прекращает функционирование, когда внешняя температура достигает температуры разогрева скопления пчел (средняя объемная температура оболочки сравнивается с температурой камеры биокалориметра). Максимальное значение разогрева пчел зависит от их численности и физиологического состояния. Так, скопление, состоящее из 550±6 пчел, разогревается максимально до 36,4±0,4ОС, а из 3980±26 – до 38,9±0,5оС.
Эффекты скачкообразного изменения внешней температуры. Опыты выполнены в разное время на пчелах весенне-летней (летом) и осенней (зимой) генераций. Подопытные пчелы (примерно по 4,5 тыс. особей) содержались в камере калориметра. Температуру его внешней оболочки поддерживали вначале в пределах оптимального диапазона, а затем резко охлаждали или нагревали. Для охлаждения использовалась холодильная камера, для нагревания – термостат.
Под влиянием охлаждения от +6 до –15оС в течение 120 мин среднее (по пяти термодатчикам) значение температуры в скоплениях пчел, адаптировавшихся к зимовке, повышалось в среднем на 4,1оС, а у летних – на 5,8оС. Это соответствовало увеличению теплопродукции на 7,12 Вт и 11,23 Вт. У зимующих пчел она возрастала постепенно и стабилизировалась на указанном уровне через 80-85 мин, а у летних – через 35-45 мин. Что касается теплоотдачи, то у первых она достигала максимального значения через 60 (lim.54-68) и у вторых – через 40 (lim.34-45) мин.
На повышение температуры от –15оС до +6оС пчелы, адаптировавшиеся к зимовке, реагировали почти линейным уменьшением теплоотдачи, которая в течение 60 мин. (lim. 58-64) устанавливалась на уровне 2,45 Вт. У летних пчел примерно за такое же время теплоотдача достигала 3,78 Вт. Значительное превышение теплоотдачи у летних пчел по отношению к аналогичному показателю зимующих пчел обусловлено разной стратегией их реагирования на охлаждение. Зимние пчелы стремятся равномерно поддерживать температуру во всем объеме скопления, а летние – изменяют ее в относительно широких пределах. Повышенной вариабельности температуры в скоплениях летних пчел соответствует высокая изменчивость теплосодержания и теплопродукции. Поэтому адаптация пчел к охлаждению, приобретенная в ходе зимовки, способствует экономии затрат энергии на терморегуляцию.
Температурные возмущения, стимулируемые введением тепла внутрь скопления. Тепловую инерцию скоплений, образуемых пчелами, определяли при воздействии на них различных температурных возмущений. Для этого с помощью термода, расположенного внутри скопления, в него вводилось тепло. Мощность нагрева термода составляла в среднем 0,85 Вт (12,2 кал/ мин), а его поверхность разогревалась не более чем на 3,5-4,3оС. В течение всего времени температура окружающего воздуха поддерживалась на уровне 5-6оС. За тепловой отклик пчел на нагрев принималось повышение температуры камеры калориметра на 3-4оС через 10-12 мин после включения термода.
Установлено, что пороговые значения вводимого тепла, стимулирующие резкое увеличение теплоотдачи (тепловые отклики), зависят от внешней температуры и количества агрегировавшихся пчел. Скоплению, образуемому примерно 2 тыс. пчел, для стимуляции теплового отклика при –10оС требуется в среднем в 1,26 раза меньше тепла, чем при 5оС. При таком же изменении внешней температуры скоплению, содержащему 3140 пчел, для заметного повышения теплоотдачи необходимо повысить количество тепла, вводимого термодом, в 1,35 раза, а 4400 особям – 1,83 раза (рис. 6, табл. 1). Уменьшение количества тепла, необходимого для стимуляции теплового отклика, в ответ на понижение внешней температуры, очевидно, связано с изменением теплосодержания, возрастающего соответственно повышению плотности агрегирования пчел.

Рис. 6. Изменение теплоотдачи скоплением, содержавшим около 4,5 тыс. пчел под влиянием введения в него 2328 Дж (556 кал) тепла (продолжительность включения термода и развиваемая им мощность показаны пунктирной линией)
Связь между внешней температурой, количеством агрегирующихся пчел и изменением их теплосодержания указывает на наличие зоны физиологически допустимых колебаний теплосодержания, в пределах которой оно может варьировать, не оказывая влияния на изменение терморегуляторной активности насекомых. В пределах зоны оптимальности введение тепла на уровне его пороговых значений, стимулирующих резкое изменение теплоотдачи, несущественно влияет на изменение терморегуляторной активности. Таким образом, увеличение средней температуры на 1оС в скоплениях, содержащих 3±1 тыс. пчел, изменяет теплосодержания в пределах 620-1474 Дж (148-352 кал). Но это стимулирует лишь незначительные сдвиги терморегуляторной активности, которая существенно возрастает при повышении средней температуры разогрева в скоплении до 2±0,1оС теплом, вводимым термодом.
Таблица 1. Динамика пороговых значений вводимого тепла, стимулирующего активизацию терморегуляции (±m рассчитана при уровне значимости р≤0,005) а зависимости от численности пчел и внешней температуры
| Количество пчел | Количество тепла (кал), вводимого при внешней температуре | ||
| -10,0оС | 0оС | 5,2оС | |
| 1850±10 | 106±10 | 120±14 | 134±10 |
| 3140±20 | 132±12 | 164±10 | 178±12 |
| 4400±15 | 175±20 | 226±18 | 320±17 |
Мощность основного обмена в зоне оптимальности для скопления, содержащего примерно 4,5 тыс. пчел, составляет в среднем 2,09 Вт (30 кал/мин). Увеличение мощности основного обмена на 10-20% за 20 мин обеспечивает прибавку тепла на 251-502 Дж (60-120 кал), что соответствует увеличению средней температуры скопления всего на 0,2-0,3оС. Накопление такого количества тепла в скоплении не стимулирует терморегуляторную активности. Но увеличение теплоотдачи достигается в результате уменьшения плотности агрегирования.