
Содержание Глава Эволюционное моделирование. 2
| Вид материала | Документы |
- Курсовая работа по бухгалтерской (финансовой) отчётности «отчёт о прибылях и убытках:, 1262.12kb.
- Наименование и краткое содержание лекций в соответствии с Рабочей программой Тема лекций,, 195.58kb.
- Автор Горбань Валерий (соsmoglot). Украина Парадигма мироздания. Содержание: Глава, 163.17kb.
- В. Е. Латов Тульский государственный университет mibo@klax tula ru Эволюционное моделирование, 65.56kb.
- Моделирование и формализация Моделирование как метод познания Моделирование, 143.04kb.
- Информатика, вычислительная техника и инженерное образование 2011, №1(3) эволюционное, 635.81kb.
- Анализ финансовых операций. Глава Имитационное моделирование инвестиционных рисков, 859.72kb.
- Как стать предпринимателем содержание Глава I. Общие сведения о предпринимательской, 5487.38kb.
- Календарный план учебных занятий по дисциплине Моделирование информационных процессов, 24.12kb.
- Узоры Древа Жизни Глава Десять Сфир в четырех мирах Глава 10. Пути на Древе Глава 11., 3700.54kb.
Содержание
Глава 1. Эволюционное моделирование. 2
1.1 Общие понятия эволюционного моделирования. 2
1.2 Эволюционное моделирование. 10
1.2.1 Модели возникновения молекулярно-генетических систем. 10
1.2.2 Общие модели эволюции. 12
1.2.3 Эволюционные модели искусственной жизни. 14
1.2.4 Прикладное эволюционное моделирование. 16
1.3 Модели Вселенной. 19
1.3.1 Модель Фридмана. 19
1.3.2 Модель де Ситтера. 20
1.3.3 Модель раздувающейся Вселенной. 20
1.3.4 Модель квазистатической Вселенной. 21
Глава 2. Генетические алгоритмы. 23
2.1 История появления эволюционных алгоритмов. 23
2.2 Генетические алгоритмы. 26
2.3 Когда следует применять ГА. 30
2.4 Решение Диофантова уравнения. 33
Список используемой литературы. 37
Глава 1. Эволюционное моделирование.
1.1 Общие понятия эволюционного моделирования.
Хочу начать изложение своего реферата с терминологии эволюционного моделирования, а именно с термина моделирование. Моделирование - ведущий принцип современного научного познания. Человек не может видеть предмет познания целиком, во всех его проявлениях. Поэтому он ограничивает свои притязания и стремится познать какую-либо сторону этого предмета, в зависимости от стоящей перед человеком задачи. Моделирование опирается на следующие основные принципы научного познания:
- принцип редукционизма – возможность сведения более сложного к более простому. Это значит, что изучение более простого может что-то сказать и о самом объекте.
- принцип эволюции – все высшие формы постепенно развились из низших форм. Это значит, что, анализируя поведение низших форм, можно прогнозировать поведение высших форм.
- принцип рациональности – говорит, что объекты реального мира можно познавать с помощью логики и математики.
Таким образом, моделирование – это инструмент науки для познания чего-либо. При помощи моделирования мы можем увидеть многие процессы с различных сторон. Но следует отметить, что методом моделирования надо пользоваться с большой осторожностью. Любая модель отражает только какой-то фрагмент реальности и перенос закономерностей одной части на все целое может иметь непредвиденные последствия.
Чтобы перейти к терминам эволюции, рассмотрим что такое развитие.
Развитие - это необратимое, направленное, закономерное изменение материи и сознания, их универсальное свойство; в результате развития возникает новое качественное состояние объекта - его состава или структуры. Развитие - всеобщий принцип объяснения природы, общества и познания, как исторически протекающих событий.
Различают две формы развития, между которыми существует диалектическая связь: эволюционную, связанную с постепенными количественными изменениями объекта (эволюция), и революционную, характеризующуюся качественными изменениями в структуре объекта (революция). Выделяют прогрессивную, восходящую линию развития (прогресс) и регрессивную, нисходящую линию (регресс). Прогресс - направленное развитие, для которого характерен переход от низшего к высшему, от менее совершенного к более совершенному. Развитие, как бы повторяет уже пройденные ступени, но повторяет их иначе, на более высокой базе, так сказать, по спирали, а не по прямой линии; развитие скачкообразное, катастрофическое, революционное превращение количества в качество; внутренние импульсы к развитию, даваемые противоречием, сталкиванием различных сил и тенденций, действуют на данное тело или в пределах данного явления; непрерывная связь всех сторон каждого явления, связь, дающая единый, закономерный мировой процесс движения, - таковы некоторые черты диалектики, как более содержательного учения о развитии (А.К.Айламазян, Е.В.Стась).
Основной особенностью, отличающей развитие от других динамических процессов, например, от процесса роста, является качественное изменение во времени переменных, характеризующих состояние развивающейся системы (для процесса роста обычно говорят лишь о количественном изменении этих переменных). Причем качественное изменение носит скачкообразный характер. Постепенное монотонное изменение некоторого параметра в течение заметного времени сопровождается соответствующим постепенным изменением состояния системы, но в определенный момент происходит разрыв постепенности: состояние системы меняется скачком, система переходит на новый качественный уровень, количество переходит в качество. Затем повторяется все заново, но уже на новом качественном уровне (А.И.Яблонский).
В изучении развития материи современной наукой сделаны такие серьезные шаги, что сейчас можно с полным правом говорить о превращении идеи развития, эволюции в норму научного мышления для целого ряда областей знания.
Термин "эволюция" имеет несколько значений, однако чаще всего он используется как синоним развития. Так, И.И.Шмальгаузен определяет эволюцию как закономерный процесс исторического развития организма. Иногда термин "эволюция" используют в более узком смысле, понимая ее как одну из форм развития, которая противопоставляется революции. Эволюция и революция рассматриваются как взаимообусловленные стороны развития, выступая против абсолютизации какой-либо из них. В любых процессах развития естественно наличие чередующихся участков: эволюционных и революционных.
Термин “революция” в большой советской энциклопедии трактуется следующим образом. Революция - глубокое качественное изменение в развитии каких-либо явлений природы, общества или познания. Наиболее широко понятие революция применяется для характеристики общественного развития. Понятие революция — неотъемлемая сторона диалектической концепции развития. Оно раскрывает внутренний механизм закона перехода количественных изменений в качественные. Революция означает перерыв постепенности, качественный скачок в развитии. Революция отличается от эволюции — постепенного развития какого-либо процесса, а также от реформы, находясь с ней в сложном соотношении, характер которого определяется конкретно-историческим содержанием самой революции и реформы.
Эволюция в широком смысле - представление об изменениях в природе и в обществе, их направленности, порядке, закономерностях; определенное состояние какой-либо системы рассматривается как результат более или менее длительных изменений ее предшествовавшего состояния; в более узком смысле - представление о медленном постепенном количественном изменении.
Эволюция в биологии - это необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием видов, преобразованием биогеоценозов и биосферы в целом.
Кратко рассмотрим историю эволюционного учения. Эволюционная идея зародилась и развилась в XIX в. в качестве оппозиции представлению о неизменности мира, но своего апогея она достигла в нашем столетии, и ее принятие можно считать достижением XX в. В прошлом веке идея неизменчивости органического мира нашла свое яркое выражение в лице Ж.Кювье. Кювье исходил из своей теории постоянства и неизменности видов и ее двух основных принципов - принципа корреляций и принципа условий существования. Неизменность вида входила, согласно Кювье, в организованность, упорядоченность природы. Его теорию катастроф, или смену фаун и флор, в данной органической области можно назвать теорией эволюции при неизменности видов, теорией нарушения гармонии природы только в результате катастрофических событий общеземного масштаба. Теория типов, теория гармонии природы и теория неизменности видов прекрасно согласовались друг с другом и составляли фундамент естествознания первой половины XIX в. Познавательная ценность этих представлений об устойчивости органического мира была огромна. Представления о неизменности видов легли в основу их классификации. Теория типов позволяла делать прогнозы. Гениальная эволюционная идея Ламарка, на полстолетия опередившего свое время, не нашла отклика отчасти потому, что, ополчившись на постоянство вида, он направил свою полемику и против его реальности.
Ч.Дарвин впервые обосновал эволюцию и убедил своих современников именно потому, что он сочетал признание реальности вида с научной теорией его изменяемости.
В XX в. идею гармонии природы сменила идея эволюции. Принцип гармонии природы, теория типов и представление об устойчивости вида отодвинулись в сознании людей на задний план, а многим казались опровергнутыми. С течением времени, однако, полное обоснование эволюционной идеи породило свою противоположность. В науке XX в. вновь возродилась идея устойчивости. И с тем же благородным рвением, с каким человеческая мысль разрушала теорию типов и теорию неизменности видов, она устремилась на поиски механизмов поддержания устойчивости.
В.И.Вернадский сумел раскрыть на уровне биосферы в целом взаимодействие эволюционного процесса и идеи устойчивости живой природы. В 1928 г. В.И.Вернадский писал: "В геохимическом аспекте, входя как часть в мало изменяющуюся, колеблющуюся около неизменного среднего состояния биосферу, жизнь, взятая как целое, представляется устойчивой и неизменной в геологическом времени. В сложной организованности биосферы происходили в пределах живого вещества только перегруппировки химических элементов, а не коренные изменения их состава и количества - перегруппировки, не отражавшиеся на постоянстве и неизменности геологических - в данном случае геохимических процессов, в которых эти живые вещества принимали участие.
Устойчивость видовых форм в течение миллионов лет, миллионов поколений, может, даже составляет самую характерную черту живых форм".
По сложившемуся общему мнению, вершиной творчества Вернадского является учение о биосфере и об эволюционном переходе ее под влиянием человеческого разума в новое состояние - ноосферу: "Масса живого вещества, его энергия и степень организованности в геологической истории Земли непрерывно эволюционировали, никогда не возвращаясь в прежнее состояние. Преобразования в поверхностной оболочке планеты под влиянием деятельности человека стали естественным этапом этой эволюции. Вся биосфера, изменившись коренным образом, должна перейти в новое качественное состояние, сферу действия человеческого разума".
Переводя теорию Дарвина на язык кибернетики, И.И.Шмальгаузен показал, что само преобразование органических форм закономерно осуществляется в рамках относительно стабильного механизма, лежащего на биогеоценотическом уровне организации жизни и действующего по статистическому принципу. Это и есть высший синтез идеи эволюции органических форм с идеей устойчивости и идеей постоянства геохимической функции жизни в биосфере. Так воедино оказались слитыми и вместе с тем поднятыми на новый современный уровень концепции Кювье, Дарвина, Вернадского.
Снова вернемся к термину “эволюция”. И кратко рассмотрим эволюцию искусственных систем. Эволюция - процесс многомерный, и замедление ее в одном измерении (направлении) отнюдь не обязательно влечет за собой снижение темпов в других измерениях. Наоборот, замедление эволюции в одном измерении довольно часто ускоряет рост ее потенциала в другом. Во многих случаях то, что представлялось недосягаемо высоким пределом роста, в действительности - только стартовая площадка для следующей стадии развития. Эволюция искусственных систем имеет, вообще говоря, несколько пределов. Множественность пределов становится почти очевидной, если учесть, сколь широк и разнообразен спектр препятствий, возникающих на пути эволюции. Одни пределы обусловлены тем, что материал конструкции не обладает безграничной изменчивостью, в других случаях пределы возникают из-за неизменности формы. Если в одном направлении обнаруживается "узкое место", то в других, как правило, открываются какие-то новые возможности. Симбиоз можно было бы определить как технологическую находку систем под влиянием внешней среды и под ее ограничениями. Краткосрочная эволюция системы (рождение, рост, зрелось, старость) происходит под влиянием, главным образом, динамики самой системы, в то время как долгосрочная эволюция (развитие популяции, класса систем) зависит от существования более широкой интегрированной системы (т.е. надсистемы) и определяется ею. Эволюция подсистемы может развиваться в направлении возрастающей сложности, в то время как эволюция всей системы в целом развивается в направлении все большей простоты (например, внедрение микропроцессоров в технические системы).
Теперь, рассмотрев основные термины моделирования и эволюции, рассмотрим понятие “эволюционное моделирование”. Эволюционное моделирование - направление в математическом моделировании, объединяющее компьютерные методы моделирования эволюции, а также близкородственные по источнику заимствования идей другие направления в эвристическом программировании. Включает в себя как разделы генетические алгоритмы, эволюционные стратегии, эволюционное программирование, искусственные нейронные сети, нечеткую логику. Эволюционное моделирование является важной частью эволюционной биокибернетики. Рис.1 иллюстрирует области исследований, которые могут быть отнесены к эволюционной биокибернетики.
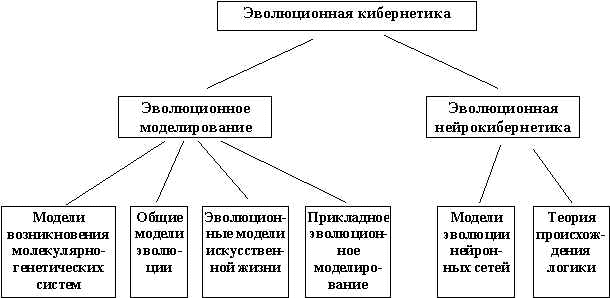
Рис.1. Области исследования эволюционной кибернетики.
В эволюционном моделировании можно выделить 4 части:
- Модели возникновения молекулярно-генетических систем.
- Моделирование общих закономерностей эволюции.
- Эволюционные модели искусственной жизни.
- Прикладное эволюционное моделирование.
Разговор о моделях эволюции пойдет в следующих главах данного реферата.
1.2 Эволюционное моделирование.
1.2.1 Модели возникновения молекулярно-генетических систем.
Интенсивные исследования по моделированию возникновения молекулярно-генетических информационных систем начались в 1960-70-х годах. Интерес к этим исследованиям связан с интригующими проблемами: как могла возникнуть жизнь на Земле? Каковы могли бы быть первые кибернетические схемы функционирования первобытных организмов? Проиллюстрируем эти работы на примерах моделей квазивидов и сайзеров.
Модель квазивидов была предложена лауреатом Нобелевской премии из ФРГ М.Эйгеном. В этой модели анализируется эволюция популяции последовательностей символов (информационных аналогов цепочек ДНК или РНК). В простейшем случае последовательность представляет собой двоичный код, т.е. последовательность нулей и единиц. Последовательности обладают определенными селективными ценностями. Эволюция популяции представляет собой размножение и отбор последовательностей в соответствии с их селективными ценностями, а также мутации, т.е. случайные замены символов в последовательностях. В результате эволюции формируется квазивид: популяция, в которой наряду с оптимальной последовательностью есть множество сходных с ней мутантов. Эйген с сотрудниками провели ряд имитационных расчетов на ЭВМ и продемонстрировали достаточно быструю скорость поиска оптимальных последовательностей в модели квазивидов. Автор настоящей статьи развил модель квазивидов и получил грубую количественную оценку числа особей, участвующих в эволюционном поиске оптимальных последовательностей. Согласно этим оценкам, при эволюционном поиске оптимума число участвующих особей (растущее как небольшая степень длины последовательностей) может быть значительно меньше по сравнению со случайным перебором, при котором число участников экспоненциально растет с ростом длины последовательностей.
Модель сайзеров (syser - сокращение от system of self reproduction - самовоспроизводящаяся система) была предложена новосибирскими учеными В.А.Ратнером и В.В.Шаминым в 1980 г. Эта модель описывает систему макромолекул, в которую входят: полинуклеотидная матрица, ферменты репликации и ферменты трансляции; полинуклеотидная матрица хранит информацию, кодирующую синтезируемые в сайзере ферменты, ферменты репликации обеспечивают копирование полинуклеотидных цепочек, а ферменты трансляции - синтез ферментов в соответствии с закодированной в матрице информацией. Сайзеры - вполне естественная модель простейшей самовоспроизводящейся системы, поэтому неудивительно, что она под разными названиями и в несколько различных формах была предложена независимо от Ратнера и Шамина и другими авторами: Д.Уайтом (Калифорния, 1980 г.) и Р.Файстелем (Берлин, 1983 г.). По общей структуре модель сайзеров сходна со схемой самовоспроизводящихся автоматов, исследованных на заре развития вычислительной техники Дж. фон Нейманом. Самовоспроизводящиеся автоматы по фон Нейману содержат хранящую информацию ленту (аналог полинуклеотидной матрицы), автомат для копирования ленты (аналог ферментов репликации) и автомат для синтеза произвольного автомата по закодированной в ленте информации (аналог ферментов трансляции). Концепция биологических информационных систем, основанная на аналогиях с самовоспроизводящимися автоматами фон Неймана, разработана в книге В.И.Корогодина "Информация и феномен жизни".
1.2.2 Общие модели эволюции.
Общие модели эволюции разрабатывались сравнительно давно. В 1910-1930-х годах классическими работами Р. Фишера ( R.A.Fisher), Дж. Холдейна (J.B.S. Haldane), и С. Райта (S.Wright) были заложены основы популяционной генетики. Классическая теоретическая популяционная генетика была основана на синтетической теории эволюции, т.е. на синтезе Дарвиновской концепции естественного отбора и Меделевской дискретной генетики. Согласно синтетической теории эволюции, главный механизм прогрессивного развития - естественный отбор тех организмов, которые смогли получить выгодные мутации.
Теоретическая популяционная генетика характеризует популяцию как динамический процесс эволюции распределения частот генов.
Математическая популяционная генетика на базе синтетической теории эволюции интенсивно развивалась до 60-х годов, когда возникли определенные трудности, связанные с экспериментальными достижениями молекулярной генетики (оценки скорости эволюционной замены аминокислот в белках и полиморфизма белков). Чтобы проинтерпретировать экспериментальные результаты, М.Кимура предложил так называемую теорию нейтральной эволюции. Основное предположение теории М.Кимуры: мутации на молекулярном уровне (замены нуклеотидов в ДНК и аминокислот в белках) преимущественно нейтральны, или слабо невыгодны. Используя математические методы популяционной генетики, М.Кимура вывел ряд следствий теории нейтральности, которые находятся в довольно хорошем согласии с экспериментальными данными. Образно говоря, теория нейтральности – это популяционная генетика на базе достижений молекулярной биологии. Процитируем пять принципов этой теории. Первые четыре из них – эмпирические, а пятый установлен теоретическим путем.
- Скорость эволюции любого белка, выраженная через число аминокислотных замен на сайт в год, приблизительно постоянна и одинакова в разных филогенетических линиях, если только функция и третичная структура этого белка остаются в основном неизменными.
- Функционально менее важные молекулы и их части эволюционируют (накапливая мутационные замены) быстрее, чем более важные.
- Мутационные замены, приводящие к меньшим нарушениям структуры и функции молекулы (консервативные замены), в ходе эволюции происходят чаще тех, которые вызывают более существенное нарушение структуры и функции этой молекулы.
- Появлению нового в функциональном отношении гена всегда должна предшествовать дупликация гена.
- Селективная элиминация вредных мутаций и случайная фиксация селективно нейтральных или очень слабо вредных мутаций происходят в ходе эволюции гораздо чаще, чем положительный дарвиновский отбор благоприятных мутаций.
Среди работ, посвященных моделированию общих кибернетических закономерностей биологической эволюции, отметим также интересный цикл исследований С.Кауффмана, посвященный анализу автоматов, состоящих из множества случайно связанных логических элементов. Отдельный автомат можно рассматривать как модель молекулярно-генетической системы управления живой клетки, при этом каждый логический элемент интерпретируется как регулятор синтеза определенного фермента. Примечательно, что при исследовании автоматов Кауффмана применяются высокоэффективные математические методы статистической физики. Модели Кауффмана позволяют сделать ряд предсказаний относительно "программ" жизнедеятельности клеток. В частности, продемонстрировано, что для одновременного обеспечения устойчивости и гибкости программы число входов логических элементов должно быть ограничено определенным интервалом, а именно составлять величину примерно равную 2-3.
К работам по моделированию общих аспектов биологической эволюции можно отнести и исследования вопросов эволюции генетических текстов. В работах Д.С.Чернавского (Москва) исследовалась проблема возникновения в процессе эволюции новой биологически значимой информации, а также была сделана оценка вероятности случайного формирования в геноме участка ДНК, кодирующего новый функционально важный для организма белок.
1.2.3 Эволюционные модели искусственной жизни.
В конце 80-х годов сформировалось очень интересное направление кибернетических исследований – искусственная жизнь (английское название Artificial Life или ALife). Многочисленные Интернет-ссылки на работы этого направления можно найти на Интернет-сайте интересного междисциплинарного Института Санта Фе: ссылка скрыта . Основной мотивацией исследований искусственной жизни служит желание понять и промоделировать формальные принципы организации биологической жизни. Как сказал руководитель первой международной конференции по искусственной жизни К.Лангтон (C.Langton) “основное предположение Искусственной Жизни состоит в том, что “логическая форма” организма может быть отделена от материальной основы его конструкции”. “Организмы” в искусственной жизни – придуманные людьми объекты, живущие в мире компьютерных программ.
Приведем некоторые примеры характерных исследований Искусственной жизни:
- исследование динамики жизнеподобных структур в клеточных автоматах (К.Лангтон);
- эволюция двух конкурирующих популяций, одна из которых есть популяция программ, решающих определенную прикладную проблему (проблему сортировки), а вторая – популяция задач, эволюционирующих в направлении усложнения проблемы (Д.Хиллис (D.Hillis));
- эволюция и формирование “биоразнообразия” самовоспроизводящихся программ, живущих в виртуальных компьютерах (Т.Рей (T.Ray));
- анализ "биоразнообразия" самовоспроизводящихся программ на базе теории самоорганизованной критичности (К.Адами (C.Adami));
- компьютерная модель ПолиМир (PolyWorld), разработанная Л.Ягером (L.Yaeger), в которой эволюция популяции искусственных организмов (агентов) происходит вполне естественным образом: агенты питаются растущей на лужайках травой, могут бороться друг с другом (в результате борьбы агент может погибнуть, тогда он превращается в пищу, которую может съесть победитель), могут скрещиваться, давая потомков - новых агентов; агенты имеют нейронную сеть, которая управляет поведением агентов.
Отметим, хотя лозунг “Искусственная Жизнь” провозглашен в конце 80-х, в действительности идейно близкие модели разрабатывались в 50-70-е годы. Приведем два примера из истории отечественной науки.
В 60-х годах блестящий кибернетик и математик М.Л.Цетлин предложил и исследовал модели автоматов, способных адаптивно приспосабливаться к окружающей среде. Работы М.Л.Цетлина инициировали целое научное направление, получившее название “коллективное поведение автоматов”.
В 60-70-х годах под руководством талантливого кибернетика М.М.Бонгарда была построена весьма нетривиальная модель “Животное”, характеризующая адаптивное поведение искусственных организмов, живущих на разбитой на клетки плоскости и обладающих рядом конкурирующих между собой потребностей.
Эволюция кибернетических сущностей – одно из главных направлений исследований Искусственной жизни.
1.2.4 Прикладное эволюционное моделирование.
Согласованность и эффективность работы элементов биологических организмов наводит на мысль – можно ли использовать принципы биологической эволюции для оптимизации практически важных для человека систем?
В нескольких модификациях подобные идеи возникали у ряда авторов. В 1966 году Л. Фогель, А. Оуэнс, М. Уолш предложили схему эволюции логических автоматов, решающих задачи прогноза. В 1975 г. вышла основополагающая книга Дж. Холланда “Адаптация в естественных и искусственных системах”, в которой был предложен генетический алгоритм, исследованный в дальнейшем учениками и коллегами Дж. Холланда в Мичиганском университете. Примерно в это же время группа немецких ученых (И. Рехенберг, Г.-П. Швефель и др.) начала разработку так называемой эволюционной стратегии. Эти работы заложили основы прикладного эволюционного моделирования или эволюционных алгоритмов.
В нашей стране исследования по прикладному эволюционному моделированию, идейно близкие к работам Л. Фогеля с сотрудниками, были разносторонне развиты в работах И.Л. Букатовой.
В общем виде эволюционный алгоритм – это оптимизационный метод, базирующийся на эволюции популяции “особей”. Каждая особь характеризуется приспособленностью – многомерной функцией ее генов. Задача оптимизации состоит в максимизации функции приспособленности. В процессе эволюции в результате отбора, рекомбинаций и мутаций геномов особей происходит поиск особей с высокими приспособленностями.
Основные эволюционные алгоритмы:
- генетический алгоритм, предназначенный для оптимизации функций дискретных переменных и акцентирующий внимание на рекомбинациях геномов;
- эволюционное программирование, ориентированное на оптимизацию непрерывных функций без использования рекомбинаций;
- эволюционная стратегия, ориентированная на оптимизацию непрерывных функций с использованием рекомбинаций;
- генетическое программирование, использующее эволюционный метод для оптимизации компьютерных программ.
По сравнению с обычными оптимизационными методами эволюционные алгоритмы имеют следующие особенности: параллельный поиск, случайные мутации и рекомбинации уже найденных хороших решений. Они хорошо подходят как простой эвристический метод оптимизации многомерных, плохо определенных функций.
Наибольшее распространение получил генетический алгоритм. На его основе осуществляются: оптимизация профилей балок в строительстве, распределение инструментов в металлообрабатывающих цехах, обработка рентгеновских изображений в медицине, оптимизация работы нефтяных трубопроводов и т.д. Одна из основных областей применения генетического алгоритма – решение задач комбинаторной оптимизации (например, задача о коммивояжере).
Для иллюстрации принципов работы эволюционных алгоритмов рассмотрим подробнее генетический алгоритм во второй части данного реферата.
Далее рассмотрим эволюцию моделей Вселенной.
1.3 Модели Вселенной.
Существует много моделей эволюции Метагалактики и Вселенной в целом, но точный сценарий пока неясен. Общепризнанной, пожалуй, является картина Большого взрыва, когда Вселенная начала расширяться из состояния сверхплотного вещества. Однако интерпретация сути этого взрыва даже в среде ученых разная. Одни, например, полагают, что произошел "направленный наружу взрыв". Для других "эта космологическая теория представляет собой верх абсурда", или "типичный пример научной фантастики". Третьи отрицают идею "взрыва бомбы в евклидовом пространстве", признавая картину расширения в целом научной. Столь большой разброс мнений позволяет или согласиться с одной из точек зрения, или выработать свою собственную. Но прежде, чем это сделать, рассмотрим некоторые модели эволюции Вселенной (согласующиеся с наблюдательными данными и являющиеся при определенных условиях следствием общей теории относительности).
1.3.1 Модель Фридмана.
Расширение Вселенной началось из сверхплотного состояния вещества, занимающего нулевой объем, и продолжается вот уже в течение 1010 лет. У этой модели есть две проблемы.
Во-первых, проблема начальной сингулярности, то есть бесконечной плотности вещества и нулевых размеров Вселенной в начальный момент времени. Решение этой проблемы видится ученым в разработке последовательной квантовой теории гравитации в области очень малых масштабов пространства-времени. Во-вторых, проблема горизонта. Суть этой проблемы сводится к следующему. В настоящее время некоторые галактики обнаруживаются на расстоянии в 1010 световых лет от Земли. В процессе расширения Вселенной за 1010 лет (возраст Вселенной) на такое расстояние мог переместиться только свет, но никак не галактики. Проблема эта не находит своего решения в рамках фридмановской модели.
1.3.2 Модель де Ситтера.
Вселенная в течение очень длительного времени практически не меняла свои крайне малые (но не нулевые!) размеры. Затем, начиная с некоторого начального момента времени, в течение очень короткого времени Вселенная расширилась до невообразимо больших размеров, сохраняя в среднем плотность вещества постоянной. В этой модели устраняется сингулярность, но добавляются проблемы сохранения средней плотности материи в процессе ее расширения и сверхсветовые скорости расширения вещества. Поэтому пределы применимости ранней версии модели де Ситтера ограничивались пустым пространством, которое расширяется, не меняя свою плотность, и в котором вещество не перемещается.
1.3.3 Модель раздувающейся Вселенной.
Эта модель является синтезом двух предыдущих. Вначале, в течение времени от 10-43с до 10-35с Вселенная расширялась "по де Ситтеру" и увеличилась до огромных размеров. Далее Вселенная начала развиваться "по Фридману". Синтез обеих моделей оказался возможным благодаря открытию нового вида материи - физического вакуума. Проблемы горизонта в этой модели больше нет: во время первой фазы расширения Вселенная "распадается на множество малых областей, которые являются зародышами метагалактик, развивающихся в дальнейшем по Фридману". Поэтому расстояние от Земли до далеких галактик никак не связано с возрастом нашей Метагалактики. Нет и проблемы начальной сингулярности с ее бесконечно большой плотностью вещества. Однако хоть и не бесконечное, но все же сверхплотное состояние вещества, существовавшее в первые мгновения расширения "по Фридману", вынуждает исследователей искать более правдоподобные модели эволюции Вселенной.
1.3.4 Модель квазистатической Вселенной.
Первая такая модель была предложена Эйнштейном. В ней силы притяжения между телами компенсируются космическими силами отталкивания. "Но после того, как из результатов Хэббла стало ясно, что внегалактические туманности разбегаются", Эйнштейн отказался от искусственно введенных в теорию сил отталкивания и, тем самым, от статической модели Вселенной.
Еще одна модель статической Вселенной появилась в результате неудовлетворенности идеей Большого взрыва с его начальным сверхплотным состоянием вещества. Эта модель основана на новой теории - масштабно-инвариантной теории тяготения, в которой массы всех объектов увеличиваются с течением времени. Красное смещение в спектрах галактик является не следствием их разбегания, а следствием роста массы. Естественное решение в рамках этой гипотезы получают также "происхождение микроволнового фонового излучения, связь между черными и белыми дырами, загадочные красные смещения в спектрах квазаров. Момент времени, соответствующий в модели Фридмана сингулярности, в этой модели представляет собой время, когда масса Метагалактики равнялась нулю.
Теперь небольшой комментарий по поводу этих моделей. Допустим, что модель расширяющейся Вселенной в настоящее время заслуживает большего доверия, чем статическая модель (большинство ученых считает именно так). Тогда Вселенная когда-то занимала ограниченный объем и за конечное время не могла расшириться до бесконечно больших размеров. Тем не менее некоторые авторы говорят о протяженности пространства в случае открытых моделей Фридмана "во все стороны до бесконечности" или о наличии в таких пространствах "бесконечного количества небесных тел, элементарных частиц и галактик". Однако трудно согласиться с тем, что за конечный промежуток времени Вселенная расширилась до бесконечности. Поэтому представляется, что в случае открытых моделей Вселенной речь может идти о их бесконечности только в потенциальном, а не в актуальном смысле, то есть о том, что в любой момент времени "количество небесных тел, элементарных частиц и галактик" во Вселенной ограничено.