
Задачи темы: 1 Дать понятие клетки как основной структурной и функциональной единицы растительного организма. 2 выявить взаимосвязь между структурой химических веществ и их физиологической ролью
| Вид материала | Документы |
Содержание2. 5. Поступление воды в растительную клетку 2.5.1. Диффузия и осмос 2. 6. Клетка как осмотическая система |
- План лекции: Разделение химических элементов по количественному содержанию. Вода,, 130.01kb.
- Вить взаимосвязь между понятиями "искусство", "литература" и "журналистика", выявить, 389.32kb.
- Задачи: Дать понятие контраста в литературе как художественного приема, 137kb.
- Генная инженерия, 313.05kb.
- Известно, что между пулом свободных аминокислот крови и усвоением питательных веществ, 65.14kb.
- Темы рефератов по курсу «Основы инженерного творчества», 8.74kb.
- Тема: «Роль минеральных веществ в жизнедеятельности организма Значение молока в питании, 55.24kb.
- Тема урока: Общие сведения о пище, 16.16kb.
- Основой жизнедеятельности клетки и организма являются обмен веществ и превращение, 36.92kb.
- Рабочая программа дисциплины «физиология и биохимия бактерий» Код дисциплины по учебному, 112.26kb.
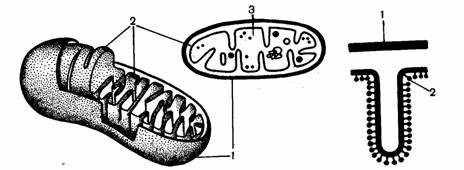
Рис.6. Схема строения митохондрии: 1 – наружная мембрана, 2 – внутренняя мембрана, 3 – матрикс.
Ядро. В растительной клетке ядро имеет диаметр порядка 10 мкм. Оно может быть сферическим, удлиненным или лопастным. Внутреннее содержимое ядра (нуклеоплазма) ограничено ядерной оболочкой, состоящей из двух элементарных мембран — внутренней и наружной. Ядерная оболочка пронизана порами диаметром 10 — 20 нм, через которые
транспортируются нуклеиновые кислоты и белки. С наружной мембраной ядерной оболочки могут быть непосредственно связаны элементы эндоплазматического ретикулума. В нуклеоплазме локализованы участки хроматина, состоящего из комплекса ДНК, РНК и белков. В делящихся клетках хроматин организуется в хромосомы, число которых специфично для каждого вида растений. Количество ДНК в одном ядре постоянно для каждого вида (кроме случаев мейоза и полиплоидии). Геном растений отличается от генома животных большим количеством ДНК. В нуклеоплазме ядра содержатся также ферменты и кофакторы, необходимые для обеспечения процессов репликации и транскрипции ДНК, молекулы различных РНК, ферменты, фосфорилирующие и ацетилирующие ядерные белки, ферменты гликолиза и др.
Ядрышко отчетливо видно в ядре под световым и электронным микроскопами. Оно формируется на определенных участках ДНК, называемых ядрышковым организатором. В хроматине ядрышка находятся участки ДНК, ответственные за синтез рибосомальных РНК (рРНК).
В целом ядро является местом хранения генетической информации клетки и репликации ДНК. В нем происходит процесс транскрипции ДНК в РНК различных типов. В тесном взаимодействии с цитоплазмой ядро участвует в обеспечении экспрессии генетической информации и контролирует процессы жизнедеятельности клетки.
Рибосомы. Рибосомы осуществляют синтез белков - трансляцию матричной, или информационной, РНК (мРНК). На электронных фотографиях они выглядят округлыми частицами диаметром 20-30 нм. Каждая рибосома состоит из двух нуклеопротеиновых субъединиц. В цитоплазме растительных клеток находятся 80 S рибосомы, состоящие из 40 и 60 S субъединиц, в хлоропластах - 70 S рибосомы, а в митохондриях — 78 - 80 S рибосомы, отличные от цитоплазматических и хлоропластных. Субъединицы рибосом, образованные в ядрышке, поступают в цитоплазму, где происходит сборка рибосом на молекуле мРНК.
Рибосомы в цитоплазме могут быть свободными, прикрепленными к мембранам эндоплазматического ретикулума, к наружной мембране ядерной оболочки либо образуют полирибосомные (полисомные) комплексы. Полирибосомы возникают потому, что одну молекулу мРНК могут одновременно транслировать несколько рибосом. Полирибосомы разрушаются при воздействии на растения неблагоприятных факторов внешней среды (например, засухи, недостатка кислорода). В процессе синтеза белка, осуществляемого рибосомами, участвуют компоненты, образованные в ядре, ядрышке, в цитоплазме, митохондриях и хлоропластах.
Пластиды. Для клеток растений типична система органоидов, называемая пластидной. Пластиды - это ограниченныедвойной мембраной округлые или овальные органоиды, содержащие внутреннюю систему мембран. Пластиды могут быть бесцветными (пропластиды, лейкопласты, этиопласты) или окрашенными (хлоропласты, хромопласты)
В меристемах присутствуют пропластиды, у которых внутренняя мембрана имеет лишь небольшие инвагинации внутренняя мембрана имеет лишь небольшие инвагинации (впячивания). Если структура пропластид сохраняется у органоидов зрелых клеток, их называют лейкопластами. В лейкопластах откладываются запасные вещества и названия они получают в зависимости от этих соединений: если запасается крахмал — амилопласты, жиры — элайопласты, белки — протеинопласты и т. д. Этиопласты формируются при выращивании зеленых растений в темноте.
Важнейший для всего живого процесс преобразования энергии поглощенного света в химическую энергию органических веществ, синтезируемых из СО2 и Н2О (фотосинтез), осуществляется в зеленых пластидах — хлоропластах. Это овальные тела 5 — 10 мкм длиной и 2 — 3 мкм в диаметре. В одной клетке листа могут находиться 15-20 и более хлоропластов, а у некоторых водорослей — лишь 1 — 2 гигантских хлоропласта различной формы. Как и другие пластиды, хлоропласты обладают наружной и внутренней мембранами (рис.7). Внутренняя мембрана, ограничивающая внутреннюю гомогенную среду (строму) хлоропласта, образует уплощенные инвагинации — тилакоиды, которые могут иметь форму дисков и в этом случае называются тилакоидами гран (гранальными). Несколько таких тилакоидов, лежащих друг над другом, формируют стопку - грану. Другие тилакоиды, связывающие между собой граны или не контактирующие с ними, называются тилакоидами стромы.
В мембранах тилакоидов локализованы зеленые (хлорофиллы), желтые и красные (каротиноиды) пигменты, компоненты редокс-цепей и запасания энергии, участвующие в погло щении и использовании энергии света. Биохимические системы синтеза и превращения углеводов функционируют в строме хлоропластов. В ней же может откладываться крахмал.
Хромопласты содержат в везикулах (пузырьках) стромы каротиноиды. Их присутствием объясняется окраска плодов томатов, рябины и др. В строме всех пластид обнаружены кольцевые молекулы ДНК.
При росте клетки количество хлоропластов увеличивается путем деления. При делении ламеллярная система пересекается перемычкой поперек органоида, иногда наблюдается почкование хлоропластов. Затем размер дочерних хлоропластов увеличивается. Деление хлоропластов происходит через 6 - 20 ч и не обязательно совпадает с делением ядра. Оно может регулироваться красным светом (660 нм) и устраняется облучением дальним красным светом (730 нм). Деление останавливается также низкой температурой.
2.4. Обмен веществ. Ферменты.
В основе специфических свойств каждой клетки, каждого организма, которые передаются по наследству, лежит специфика обмена веществ. Обмен веществ — это совокупность всех происходящих в организме химических процессов.
Химические реакции, составляющие обмен веществ, тесно взаимосвязаны и согласованы друг с другом. Обмен веществ внутри клетки тесно взаимосвязан со средой. Из внешней среды поступают вещества, необходимые для жизнедеятельности организма, и определенные вещества выделяются организмом в среду. Условия среды (температура, влажность, освещение), в которых осуществляется жизнедеятельность организма, оказывают глубокое влияние на скорость и направленность обмена. Организм обладает способностью регулировать обмен веществ.
Обмен веществ (метаболизм) имеет 2 стороны:
1) ассимиляция (анаболизм), сущность которой заключается в усвоении углерода и биосинтезе первичных углеводов (т.е. осуществление процесса фотосинтеза).
2) диссимиляция (катаболизм), сущность этого процесса заключается в осуществлении процессов дыхания и брожения, которые связаны с высвобождением энергии, необходимой для нужд клетки.
Даже поверхностное рассмотрение особенностей химических процессов в живой клетке позволяет заметить, что они протекают с огромной скоростью. И все они являются каталитическими и осуществляются благодаря присутствию биологических катализаторов — ферментов, ничтожное количество которых осуществляет колоссальный объем превращений.
Для осуществления химических реакций необходимо, чтобы молекулы были в активном состоянии. В самом общем виде катализаторы, повышая активность реагирующих молекул, как бы снижают силы химического сопротивления. Вместе с тем катализаторы не могут вызвать реакцию, которая в их отсутствие не идет, они ускоряют лишь реакции, которые термодинамически осуществимы. Многие ферменты ускоряют реакции в 109— 1011 раз. В отсутствие ферментов биохимические реакции происходили бы настолько медленно, что жизнь была бы невозможной.
По химической природе все ферменты — простые или сложные белки. Небелковый компонент, прочно связанный, называют простетической группой, слабо связанный, обслуживающий несколько ферментов,— коферментом. Часто кофермент соединяется с соответствующим белком только в период реакции. Состав простетических групп или коферментов разнообразен. Во многих случаях это витамины или их производные, в частности производные витаминов B1, B2, В6 никотиновой кислоты и др. В простетическую группу ряда ферментов входят металлы (железо, медь, цинк). Металлы могут входить в состав простетических групп в виде особых соединений, например железо в соединении с 4 пиррольными группировками (геминовое железо). Многие коферменты принимают участие в окислительно-восстановительных реакциях (НАД, НАДФ, и др.).
Важнейшим свойством ферментов является их специфичность. Еще в 1911 г. крупный немецкий химик Эмиль Фишер выдвинул положение, что фермент должен подходить к субстрату, как «ключ к замку». Однако во многих случаях пространственное соответствие молекулы фермента и субстрата возникает лишь в процессе их взаимодействия (Кошланд). Ферменты — это не только катализаторы, но и регуляторы процессов обмена. В клетке содержатся сотни соединений и должны бы происходить бесчисленные реакции. Однако число реакций ограничивается, поскольку специфичность ферментов позволяет различать определенные молекулы. Каждый организм имеет свой набор ферментов, обусловленный его наследственной основой.
Действие фермента проходит в несколько стадий. Начальной стадией является образование комплекса фермента с субстратом. При этом между ферментом и субстратом возникают связи разного характера (водородные, ван-дер-ваальсовы и др.). Именно образование фермент-субстратного комплекса требует высокой специфичности фермента. Как правило, молекула субстрата очень мала по сравнению с молекулой фермента. Поэтому при образований фермент-субстратного комплекса участвует лишь незначительная часть молекулы фермента, его активный центр.
Активный центр — это совокупность функциональных групп, принимающих непосредственное участие в ферментативной реакции.
Образование фермент-субстратного комплекса вызывает переход субстрата в более реактивное состояние, его активацию. Известно, что при любом химическом взаимодействии вступают в реакцию только те молекулы, которые обладают избытком энергии. Число столкновений между молекулами, приводящее к химическому взаимодействию (число эффективных столкновений), составляет лишь некоторую долю (иногда очень малую) общего числа столкновений. Эффективными оказываются лишь столкновения между молекулами, которые в этот момент обладают некоторым избытком внутренней энергии по сравнению со средней (для данной температуры) величиной. Энергия, которую необходимо придать молекулам вещества А для превращения их в В, сверх той средней, которую молекулы А уже содержат, называют энергией активации. В самом общем виде фермент благодаря созданию фермент-субстратного комплекса проводит реакцию обходным путем и тем самым снижает энергию активаций или энергетический барьер.
Выделяют три фазы действия фермента: 1) образование фермент-субстратного комплекса; 2) преобразование промежуточного соединения в один или несколько активных комплексов; 3) выделение продуктов реакции и регенерация молекулы фермента.
Ферменты проявляют свою активность при выделении из клетки. Однако в клетке их действие может отличаться от действия вне клетки. Ферменты могут быть локализованы в определенных частях клетки, вплетены в мембраны, пространственно разделены или, наоборот, объединены с субстратом. Все это накладывает большой отпечаток на их работу.
Разнообразие ферментов в клетке чрезвычайно велико, однако все их можно разделить на шесть классов: 1) оксидоредуктазы - катализирующие окислительно-восстановительные реакции; 2) трансферазы — катализирующие перенос целых атомных группировок от одного соединения к другому; 3) гидролазы — осуществляющие распад различных органических соединений с участием воды (гидролиз); 4) лиазы — катализирующие присоединения какой-либо атомной группировки к органическим соединениям или отцепляющие от субстратов определенную группу без участия воды; 5) изомеразы — катализирующие превращение одних изомеров в другие; 6) лигазы, или синтетазы,— катализирующие синтез органических соединений, происходящий при участии АТФ (с использованием энергии этого соединения).
Белки-ферменты, катализирующие одну и ту же реакцию, встречающиеся у одного вида организмов, но различающиеся по ряду физико-химических свойств (внутренней локализации, электрофоретической подвижности), называют изоферментами. Они различаются по реакции на внешние условия; их максимальная активность проявляется в различных условиях температуры и рН. По-видимому, наличие изоферментов позволяет организмам лучше приспосабливаться к меняющимся условиям среды. Скорость и направленность ферментативных реакций клетки зависят от следующих причин: 1) условий, в которых протекает ферментативная реакция; 2) активности действующего фермента; 3) количества белка-фермента.
2. 5. ПОСТУПЛЕНИЕ ВОДЫ В РАСТИТЕЛЬНУЮ КЛЕТКУ
Для осуществления всех процессов жизнедеятельности в клетку из внешней среды должны поступать вода и питательные вещества. Вода прямо или косвенно участвует во всех реакциях обмена и является важнейшей составной частью растительной клетки.
2.5.1. ДИФФУЗИЯ И ОСМОС
Прежде чем перейти к изучению поступления воды, остановимся на рассмотрении некоторых общих закономерностей.
Как известно, при температуре выше абсолютного О °С все молекулы находятся в постоянном беспорядочном движении. Это показывает, что они обладают определенной кинетической энергией. Благодаря постоянному движению при смешении двух жидкостей или двух газов их молекулы равномерно распределяются по всему доступному объему.
Диффузия — это процесс, ведущий к равномерному распределению молекул растворенного вещества и растворителя. Как всякое движение, диффузия требует энергии. Диффузия всегда направлена от большей концентрации данного вещества к меньшей, от системы, обладающей большей свободной энергией, к системе с меньшей свободной энергией. Свободной энергией называется часть внутренней энергии системы, которая может быть превращена в работу. Свободная энергия, отнесенная к 1 молю вещества, носит название химического потенциала. Таким образом, химический потенциал — это мера энергии, которую данное вещество использует на реакции или движение. Химический потенциал — функция концентрации. Скорость диффузии зависит от температуры, природы вещества и разности концентраций. Чем выше концентрация данного вещества, тем выше его активность и его химический потенциал. Диффузионное передвижение вещества всегда идет от большего к меньшему химическому потенциалу. Наибольший химический потенциал у чистой воды. Добавление к воде молекул растворенного вещества приводит к возникновению связи между молекулами воды и растворенного вещества, что уменьшает ее активность, ее свободную энергию, ее химический потенциал. В том случае, если диффундирующие вещества встречают на своем пути мембрану, движение замедляется, а в некоторых случаях прекращается.
Диффузия воды по направлению от своего большего к меньшему химическому потенциалу через мембрану носит название осмоса.
Иначе говоря, осмос — это диффузия воды или другого растворителя через полупроницаемую перепонку, вызванная разностью концентраций или разностью химических потенциалов. Осмос — результат неравенства химических потенциалов воды по разные стороны мембраны.
Идеальная полупроницаемая мембрана пропускает молекулы воды и не пропускает молекулы растворенного вещества. В 1877 г. немецкий физиолог Пфеффер приготовил искусственную полупроницаемую мембрану. Для этого в пористый фарфоровый сосуд наливали раствор медного купороса и помещали в другой сосуд, в свою очередь заполненный раствором ферроцианида калия. В порах первого фарфорового сосуда растворы соприкасались и реагировали друг с другом. В результате в порах образовалась пленка из ферроцианида меди [Сu2Fe(СN)6], которая обладала полупроницаемостью. Таким образом, была создана как бы модель клетки: полупроницаемая пленка имитировала цитоплазму, а стенки сосуда —пектоцеллюлозную оболочку. Сосуд, в порах которого образовалась полупроницаемая мембрана, заполненный раствором сахарозы, помещали в воду. Такой прибор получил название осмометра.
Химический потенциал воды во внутреннем сосуде будет тем меньше, чем выше концентрация сахарозы. Таким образом, поступление воды в раствор через полупроницаемую перегородку обусловливается разностью между свободной энергией чистой воды и раствора и происходит самопроизвольно по градиенту свободной энергии воды. В осмометре при наличии полупроницаемой мембраны вода будет поступать в раствор, который при этом будет становиться все более разбавленным и движение воды будет замедляться. Однако равновесия не наступит, поскольку возникнет гидростатическое давление. Если к осмометру присоединить трубку, вода будет подниматься по ней. Наконец давление столба воды уравновесит силу, с которой молекулы воды поступают в осмометр. Таким образом, энергия молекул воды, которая уменьшилась благодаря введению растворенного вещества, восполнится давлением столба воды либо грузом, т. е. давлением, которое мы приложим извне. Это давление повышает химический потенциал раствора (Iр), делая его равным химическому потенциалу чистой воды (IВ).
Согласно закону термодинамики самопроизвольно идут только процессы, сопровождающиеся выделением энергии. В большинстве случаев измерить свободную энергию в абсолютном выражении не представляется возможным. Однако можно измерить разность между конечным (g2) и начальным энергетическим состоянием (g1). Поскольку g1 больше g2, то изменение Δg будет иметь отрицательное значение. В нашем случае g1— это химический потенциал чистой воды, a g2—это химический потенциал воды после добавления растворенного вещества. Δg будет равно, но противоположно по знаку тому давлению, которое надо приложить к системе, чтобы предотвратить поступление в него воды через полупроницаемую мембрану. Δg и будет осмотический потенциал (Ψосм) раствора.
Таким образом, осмотический потенциал равен разности между химическим потенциалом раствора и химическим потенциалом чистой воды и всегда отрицателен. Осмотический потенциал показывает недостаток энергии в растворе по сравнению с чистой водой, вызванный взаимодействием вода — растворенное вещество. Иначе говоря, осмотический потенциал показывает, насколько прибавление растворенного вещества снижает активность воды.
Осмотический потенциал относится к так называемым коллигативным свойствам раствора, таким, как понижение точки замерзания или повышение точки кипения. Все эти показатели зависят от молярной концентрации. 1 моль раствора любого недиссоциированного вещества имеет осмотический потенциал 22,7 бара. Поскольку уменьшение химического потенциала, или активности воды, пропорционально числу частиц, то при растворении диссоциированных веществ абсолютное значение осмотического потенциала будет больше, для чего вводится соответствующий (изотонический) коэффициент. Надо учесть, что осмотический потенциал любого раствора проявляется только в условиях системы раствор — полупроницаемая мембрана — растворитель.
Сказанное дает возможность измерить осмотический потенциал раствора (Ψосм). Присоединив манометр, можно измерить давление, которое надо приложить к системе, чтобы предотвратить поступление воды в раствор. Оно будет по абсолютной величине равно, но противоположно по знаку осмотическому потенциалу раствора.
2. 6. КЛЕТКА КАК ОСМОТИЧЕСКАЯ СИСТЕМА
Растительная клетка представляет собой осмотическую систему. Пектоцеллюлозная оболочка хорошо проницаема как для воды, так и для растворенных веществ. Однако плазмалемма и тонопласт обладают избирательной проницаемостью, легко пропускают воду и менее проницаемы, а в некоторых случаях непроницаемы для растворенных веществ. В этом можно убедиться, рассмотрев явления плазмолиза и тургора. Если поместить клетку в раствор более высокой концентрации, чем в клетке, то под микроскопом видно, что протоплазма отстает от клеточной оболочки. Это особенно хорошо проявляется на клетке с окрашенным клеточным соком. Клеточный сок остается внутри вакуоли, а между протоплазмой и оболочкой образуется пространство, заполненное внешним раствором.
Явление отставания протоплазмы от клеточной оболочки получило название плазмолиза. Плазмолиз происходит в результате того, что под влиянием более концентрированного внешнего раствора вода выходит из клетки (от своего большего химического потенциала к меньшему), тогда как растворенные вещества остаются в клетке. При помещении клеток в чистую воду или в слабо концентрированный раствор вода поступает в клетку. Количество воды в клетке увеличивается, объем вакуоли возрастает, клеточный сок давит на цитоплазму и прижимает ее к клеточной оболочке. Под влиянием внутреннего давления клеточная оболочка растягивается, в результате клетка переходит в напряженное состояние (тургор).
Избирательная проницаемость мембран обеспечивает прохождение через них молекул воды, препятствует проникновению растворенных в воде веществ и обусловливает явление плазмолиза при действии на клетку гипертонического раствора. Если же молекулы растворенного вещества через мембрану проходят, но медленнее, чем молекулы воды, то начавшийся плазмолиз потом исчезает. Деплазмолиз происходит в результате постепенного проникновения растворенного вещества в клетку, изменения водного потенциала снаружи и внутри, а также поступления воды в клетку из наружного раствора по градиенту водного потенциала.
2.6.1. Формы плазмолиза
В ходе плазмолиза форма плазмолизированного протопласта меняется. Вначале протопласт отстает от клеточной стенки лишь в отдельных местах, чаще всего в уголках. Плазмолиз такой формы называют уголковым. Затем протопласт продолжает отставать от клеточных стенок, сохраняя связь с ними в отдельных местах, поверхность протопласта между этими точками имеет вогнутую форму. На этом этапе плазмолиз называется вогнутым. Постепенно протопласт отрывается от клеточных стенок по всей поверхности и принимает округлую форму. Такой плазмолиз носит название выпуклого. А если у протопласта связь с клеточной стенкой в отдельных местах сохраняется, то при дальнейшем уменьшении объема в ходе плазмолиза протопласт приобретает неправильную форму. Такой плазмолиз носит название судорожного (рис. 10). Время, в течение которого вогнутый плазмолиз переходит в выпуклый, позволяет оценивать степень вязкости цитоплазмы.