
Биология под редакцией академика рамн профессора В. Н. Ярыгина Вдвух книгах
| Вид материала | Книга |
- Решение проблем остеоартроза, 401.08kb.
- Под редакцией М. А. Пальцева, В. Н. Ярыгина, Н. Д. Ющука, И. Н. Денисова, Н. Н. Володина,, 6882.41kb.
- Под редакцией М. А. Пальцева, В. Н. Ярыгина, Н. Д. Ющука, И. Н. Денисова, Н. Н. Володина,, 6092.7kb.
- Методические рекомендации для практических занятий дисциплина «Биология с основами, 86.45kb.
- Правовых учений, 4116.46kb.
- Тестовый контроль вопросы для государственных экзаменов по терапии на 5 курсе педиатрического, 1331.38kb.
- Образовательный стандарт послевузовской профессиональной подготовки специалистов Специальность, 2055.19kb.
- Образовательный стандарт послевузовской профессиональной подготовки специалистов Специальность, 2316.49kb.
- Лекции: Современные главы биологии рекомендуемая литература основная: Биология для, 1501.07kb.
- Программа курса для аспирантов и соискателей Составители, 442.16kb.
ГЛАВА 13
ЗАКОНОМЕРНОСТИ МАКРОЭВОЛЮЦИИ
Действие элементарных эволюционных факторов приводит к дивергенции популяций и образованию новых видов. Многообразные виды связаны друг с другом разной степенью родства. Виды, дивергировавшие от общих предков относительно недавно, сохраняют нередко значительное фено- и генотипическое сходство. Дивергенция, произошедшая в отдаленное время, в результате оставляет меньшее количество общих фенотипических признаков и особенностей геномов. Виды, практически не связанные родством, характеризуются серьезными различиями.
Благодаря тому что процесс микроэволюции, завершающийся видообразованием, протекает постепенно на протяжении всего существования жизни на Земле, в живой природе образовались естественные группировки видов разного ранга родства, называемые таксонами. Тип — это наиболее крупный таксой, объединяющий виды, которые имеют сходный общий план строения благодаря отдаленному родству. Типы подразделяются на классы, далее следуют отряды, семейства и роды. Дивергенция видов, входящих в состав одного рода, произошла относительно недавно (рис. 13.1).

Рис. 13.1. Эволюция таксономических групп
Процесс формирования таксонов надвидового ранга называют макроэволюцией. Процесс макроэволюции изучают методами палеонтологии, сравнительной анатомии и сравнительной эмбриологии. Современные методы изучения макроэволюции — иммунологический и молекулярно-биологический — подробнее описаны в § 15.2.
13.1. ЭВОЛЮЦИЯ ГРУПП ОРГАНИЗМОВ
13.1.1. Уровень организации
Говоря о прогрессивной эволюции природы в целом и животного мира в частности, следует определить понятие уровень организации. Это понятие прямо не связано со степенью сложности строения животного.
Действительно, степень сложности не только морфофизиологической организации, но и генома человека и современных человекообразных обезьян практически совпадает, но уровень организации человека несомненно выше.
Предложено несколько вариантов оценки уровня организации животных. Наиболее приемлемым из них является, вероятно, определение уровня организации через характер взаимоотношения организмов данной группы со средой обитания. Примитивные организмы проявляют себя в среде пассивно. Выживание и дальнейшая эволюция их обеспечиваются в первую очередь интенсивным процессом размножения, благодаря которому компенсируются огромные потери, возникающие в процессе естественного отбора. В противоположность им высокоорганизованные организмы более активно проявляют себя в среде обитания и не только противостоят ей, но и способны преобразовывать ее, приспосабливая для своих нужд. Плодовитость их невелика.
Рассмотренный критерий уровня организации может применяться только по отношению к группам организмов, связанных более или менее отдаленным родством. Так, пресмыкающиеся более высоко организованы, чем земноводные, потому, что они активнее ведут себя •в среде обитания (размножение их может происходить в разнообразных условиях), при более низкой плодовитости выживаемость их потомства значительно выше. Уровень организации млекопитающих еще более высок благодаря постоянной температуре тела, сложному поведению, внутриутробному эмбриональному развитию и заботе о потомстве. Уровень организации человека чрезвычайно высок благодаря его социальному способу адаптации к среде и активному ее преобразованию (см. гл. 17).
13.1.2. Типы эволюции групп
В зависимости от того, изменяется ли уровень организации в эволюционирующих группах, выделяют два основных типа эволюции: аллогенез и арогенез.
При аллогенезе у всех представителей данной группы сохраняются без изменения основные черты строения и функционирования систем органов, благодаря чему уровень организации их остается прежним. Аллогенная эволюция происходит в пределах одной адаптивной зоны — совокупности экологических ниш, различающихся в деталях, но сходных по общему направлению действия основных средовых факторов на организм данного типа. Интенсивное заселение конкретной адаптивной зоны достигается благодаря возникновению у организмов идиоадаптаций — локальных морфофизиологических приспособлений к определенным условиям существования. Пример аллогенеза с приобретением идиоадаптаций к разнообразным условиям обитания в отряде насекомоядных млекопитающих см. на рис. 13.2.

Рис. 13.2. Аллогенез в отряде насекомоядных млекопитающих
Наземные формы: а—прыгунчик, б—землеройка, в—еж; земноводные формы: г—кутора, д—выдровая землеройка, е—выхухоль; роющие формы: ж—крот, з—златокрот
Арогенез — такое направление эволюции, при котором у некоторых групп внутри более крупного таксона появляются новые морфофизиологические особенности, приводящие к повышению уровня их организации. Эти новые прогрессивные черты организации называют ароморфозами. Ароморфозы позволяют организмам заселять принципиально новые, более сложные адаптивные зоны. Так, арогенез ранних земноводных был обеспечен появлением у них таких основных ароморфозов, как пятипалые конечности наземного типа, легкие и два круга кровообращения с трехкамерным сердцем. Завоевание адаптивной зоны с более сложными для жизни условиями (наземной по сравнению с водной, воздушной по сравнению с наземной) сопровождается активным расселением в ней организмов с появлением у них локальных идиоадаптаций к различным экологическим нишам.
Таким образом, периоды арогенной эволюции группы могут сменяться периодами аллогенеза, когда в результате возникающих идиоадаптаций новая адаптивная зона заселяется и используется наиболее эффективно. Если в ходе филогенеза организмы осваивают более 49
простую по сравнению с исходной адаптивную зону, например, переходя к прикрепленному неподвижному образу жизни или становись паразитами, то они претерпевают морфофизиологический регресс, или дегенерацию, утрачивая часть прогрессивных черт, которыми обладали их предки, и оказываясь на более низком уровне организации. Нередко и новая, более простая, адаптивная зона дает возможность широких идиоадаптаций к не менее разнообразным условиям обитания в ней, т.е. обеспечивает аллогенез (рис. 13.3).

Рис. 13.3. Направления эволюции групп:
I — арогенез, II — аллогенез, III — дегенерация; а, б, в — адаптивные зоны
Современный органический мир характеризуется огромным многообразием форм, отличающихся как по уровню организации, так и по разнообразию локальных адаптации именно благодаря смене типов эволюции, протекающей своеобразно в каждой из эволюционирующих групп. Наряду с высокоорганизованными формами существуют примитивные, свободноживущие и паразитические организмы, возникшие эволюционно относительно недавно, и реликтовые группы, время бурного адаптационного процесса которых давно прошло.
13.1.3. Формы эволюции групп
Существуют две элементарные формы филогенеза: филетическая и дивергентная эволюция.
Филетическая эволюция — это изменения, происходящие в одном филогенетическом стволе, эволюционирующем во времени как единое целое. Реконструированы филогенетические ряды слона, лошади и других организмов, в которых можно наблюдать постепенное нарастание количества и степени выраженности признаков, характерных для современных форм (рис. 13.4).

Рис. 13.4. Филетическая эволюция конечности в сем. Лошадиные.
I—преобразование скелета конечности; II—схема филетической эволюции:
предковая форма—фенакодус, 1— эогиппус, 2—миогиппус, 3—парагиппус, 4—плиогиппус, 5— лошадь; Ап—А5 — этапы эволюции
Дивергентная эволюция заключается в образовании на основе одной предковой группы двух или нескольких производных (рис. 13.5). Она приводит к дифференциации более крупных таксонов на более мелкие, например классов на отряды, родов на виды.
Как филетическая, так и дивергентная эволюция протекают на общей генетической базе, поэтому между организмами сохраняется более или менее выраженное генотипическое и морфофункциональное сходство.
Сопоставление филогенеза в разных группах позволяет выделить и некоторые общие закономерности соотносительной эволюции. Так, при попадании в одну и ту же среду обитания двух или более филогенетических групп неродственных организмов у них обычно проявляется конвергенция признаков. При этом сходные экологические задачи они решают сходным образом. Конвергентные адаптации возникают в этом случае на разной генетической основе, затрагивают в первую очередь поверхностные признаки, не распространяясь на общий план строения и наиболее существенные черты организации соответствующих групп. Примером конвергентной эволюции являются форма тела и особенности локомоции в воде у акуловых рыб, водных пресмыкающихся — ихтиозавров, костистых рыб, пингвинов, ластоногих и китообразных млекопитающих, внутреннее строение которых полностью соответствует особенностям, характерным для классов, к которым они относятся (рис. 13.6).
Другая форма соотносительной эволюции — параллелизм — реализуется в двух или нескольких группах, связанных более или менее отдаленным родством, которое основано на дивергенции от общего предка. В связи с общностью части генофондов, унаследованных от предков, у них возникают сходные адаптации в условиях действия факторов отбора в одинаковом направлении. Параллельное филетическое развитие двух родственных групп обеспечивается реализацией закона гомологических рядов (см. разд. 13.3.5). Примером параллелизма является эволюция одногорбого и двугорбого верблюдов соответственно в Африке и Центральной Азии, имеющих сходные адаптации к жизни в пустынях
Параллелизм можно наблюдать и в родственных группах организмов, разобщенных временем. Так, во второй половине кайнозойской эры в семействе кошачьих такой признак, как саблезубость (гипертрофия клыков верхней челюсти), возникал по крайней мере четырежды у представителей четырех разных родов (рис. 13.7).
13.1.4. Биологический прогресс и биологический регресс
Эволюция любого типа — арогенная, аллогенная или идущая по типу мофофизиологического регресса в природе — в целом приводит к всесветному расселению живых организмов разного уровня организации.

Рассматривая эволюцию отдельных таксонов, можно убедиться в том, что некоторые из них находятся в состоянии расцвета, в то время как другие вымирают. Успех группы организмов в эволюционном процессе оценивают как состояние биологического прогресса, критериями которого являются: 1) увеличение количества представителей соответствующей группы; 2) расширение ареала распространения;
3) активизация видоообразования в роде, увеличение количества родов в семействе, семейств в отряде и т.д.
Явление, противоположное биологическому прогрессу, — биологический регресс — свидетельствует об угасании филогенетической группы, ведущем к ее вымиранию. Так же как и биологический прогресс, состояние регресса может длиться очень долго. Оставшиеся немногочисленные представители некогда биологически прогрессивной группы, найдя соответствующую их организации экологическую нишу, могут продолжать существовать на протяжении сотен миллионов лет. Они называются реликтами, обладают консервативностью организации и крайне медленно эволюционируют. Примерами реликтов являются современная кистеперая рыба — латимерия, ближайшие родственники которой жили в девонском периоде более 200 млн. лет назад, а также дерево гинкго, непосредственные предки которого существовали на Земле уже в первой половине мезозойской эры.
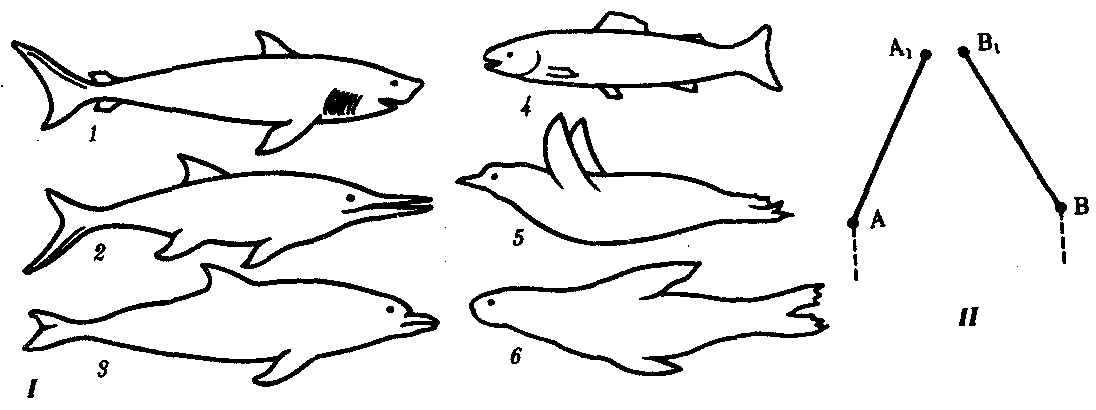
Рис. 13.6. Конвергентная эволюция формы тела позвоночных. I—водные животные разного происхождения; II—схема конвергентной эволюции:
1—акула, 2—ихтиозавр, 3—дельфин, 4—костистая рыба, 5—пингвин, 6— тюлень; A—A1, B—B1 — этапы эволюции
На протяжении исторического развития одна и та же группа может претерпевать периоды бурного расцвета, стабильного состояния, или стагнации, и упадка. Так, в девонском периоде появились первые земноводные, вставшие на путь арогенной эволюции. Заселив наземную среду обитания, они господствовали на ней в течение 75 млн. лет. На границе палеозойской и мезозойской эр (230 млн. лет назад) бурный биологический прогресс претерпевают пресмыкающиеся, быстро вытесняющие земноводных, которые с тех времен и до современного состояния переживают биологический регресс.
Расцвет пресмыкающихся длился до последней трети мезозойской эры, когда (около 65 млн. лет назад) он сменился биологическим регрессом, после чего освобождающиеся вымирающими динозаврами экологические ниши стали занимать бурно эволюционирующие млекопитающие и птицы, биологический прогресс которых продолжается до настоящего времени. В то же время такой класс, как костистые рыбы, пройдя период бурного расцвета к началу мезозойской эры и прочно заняв разнообразные места обитания, до настоящего времени находится в состоянии стабильного господства среди позвоночных в водной среде обитания.
Состояние биологического прогресса может быть обусловлено как аро- и аллогенезом, так и морфофизиологическим регрессом. Действительно, занятие новых экологических ниш как в новых адаптивных зонах, так и в уже освоенных приводит и к активизиции размножения, и к расширению ареала, и к усилению видообразования.

Рис. 13.7. Параллельная эволюция саблезубости у кошачьих. I—черепа саблезубых кошек в разные периоды кайнозойской эры; II—схемы параллельной эволюции:
а—махайрод (олигоцен - 37-25 млн. лет назад), б—смилодон (миоцен - 25-9 млн. лет назад) в—лжесаблезубая кошка (олигоцен - 37-25 млн. лет назад), г— саблезубый тигр (плейстоцен — 1,8 млн.-10 тыс. лет назад); А-А2 — этапы синхронного параллелизма, B-B2 - этапы асинхронного
13.1.5. Эмпирические правила эволюции групп
Из многочисленных обнаруженных эмпирическим путем правил и закономерностей эволюции групп остановимся на важнейших.
Согласно правилу необратимости эволюции, эволюция является необратимым процессом, и организмы не могут вернуться к прежнему состоянию, уже пройденному их предками ранее. Действительно, каждое эволюционно значимое изменение представляет собой комбинацию многих мутаций и появление новых регуляторных взаимодействий, подхваченных естественным отбором, но возникших случайно и независимо друг от друга. Поэтому понятно, что возвращение генофонда данной группы организмов или даже одного генотипа к состоянию, характерному для предков, статистически практически невероятно. Однако в ряде случаев за счет отдельных обратных мутаций или за счет мутаций регуляторных генов возможно повторное возникновение отдельных признаков атавистической природы (разд. 13.3.4).
Правило прогрессивной специализации утверждает, что филогенетическая группа, эволюционирующая по пути приспособления к данным конкретным условиям, и в дальнейшем будет продвигаться по пути углубления специализации. Генетические основы этого правила заключены в том, что в г«роцессе естественного отбора в условиях данной адаптивной зоны отсеиваются те гены генофондов популяций, которые не соответствуют ей. В результате возникает ограниченность способности генофондов к изменениям в разных направлениях.
Примером прогрессивной специализации служат морфологические преобразования конечностей в эволюционной ветви лошадей (см. рис. 13.4). При переходе к жизни на открытых пространствах с плотной почвой у предков лошади уменьшается количество пальцев до одного, что не позволяет современным лошадям населять другие биотопы. Другой пример — эволюция Юго-Восточно-азиатских человекообразных обезьян гиббонов. Специализация их к древесному образу жизни при отсутствии хватательного хвоста, имеющегося, например, у всех американских древесных обезьян, привела к возникновению своеобразного типа движения — брахиации, при котором осуществляются прыжки по веткам в подвешенном состоянии на передних конечностях. Они при этом резко удлиняются, большой палец значительно редуцируется, а рука становится почти неспособной к манипуляциям мелкими предметами. При передвижении по земле руки гиббонов уже не участвуют в локомоции.
Прогрессивная специализация резко уменьшает экологическую пластичность видов и часто является причиной их вымирания при изменившихся условиях. Уменьшение размеров и упрощение видового состава тропических лесов Индонезии являются причиной биологического регресса такого узкоспециализированного вида человекообразных обезьян, как орангутан.
Следствием правила прогрессивной специализации является правило происхождения новых групп организмов от малоспециализированных предков. Примеров, иллюстрирующих это правило, много. Действительно, млекопитающие произошли от пресмыкающихся, сохранивших в своей организации некоторые черты земноводных. Прогрессивная линия эволюции приматов, ведущая к человеку, не связана с рамапитеком (специализированной древесной формой), как полагали раньше, а берет начало от неспециализированных африканских приматов, ведущих полуназемный образ жизни (см. § 15.2 и 15.3). Генетической основой этого правила является отсутствие жесткого одностороннего отбора генотипов и сохранение в связи с этим их большего разнообразия у неспециализированных форм и, следовательно, их высокой экологической пластичности.