
Биология под редакцией академика рамн профессора В. Н. Ярыгина Вдвух книгах
| Вид материала | Книга |
- Решение проблем остеоартроза, 401.08kb.
- Под редакцией М. А. Пальцева, В. Н. Ярыгина, Н. Д. Ющука, И. Н. Денисова, Н. Н. Володина,, 6882.41kb.
- Под редакцией М. А. Пальцева, В. Н. Ярыгина, Н. Д. Ющука, И. Н. Денисова, Н. Н. Володина,, 6092.7kb.
- Методические рекомендации для практических занятий дисциплина «Биология с основами, 86.45kb.
- Правовых учений, 4116.46kb.
- Тестовый контроль вопросы для государственных экзаменов по терапии на 5 курсе педиатрического, 1331.38kb.
- Образовательный стандарт послевузовской профессиональной подготовки специалистов Специальность, 2055.19kb.
- Образовательный стандарт послевузовской профессиональной подготовки специалистов Специальность, 2316.49kb.
- Лекции: Современные главы биологии рекомендуемая литература основная: Биология для, 1501.07kb.
- Программа курса для аспирантов и соискателей Составители, 442.16kb.
14.4. КРОВЕНОСНАЯ СИСТЕМА
Необходимым условием существования высокоорганизованных крупных многоклеточных организмов является наличие жидкой подвижной внутренней среды, которая обеспечивает интеграцию организма в целостную систему, выполняя транспортные функции. Эти функции являются основными для кровеносной системы. Конкретная функция кровеносной системы зависит от того, что она транспортирует: питательные вещества, кислород, углекислый газ, другие продукты диссимиляции или гормоны. Кровеносная система всех хордовых замкнутая и состоит из двух основных артериальных сосудов: брюшной и спинной аорт. По брюшной аорте венозная кровь продвигается кпереди, обогащается кислородом в органах дыхания, а по спинной — кзади. Из спинной аорты кровь через систему капилляров возвращается по венам в брюшную аорту. Брюшная аорта или ее часть, периодически сокращаясь, проталкивает кровь по сосудам (рис. 14.27).
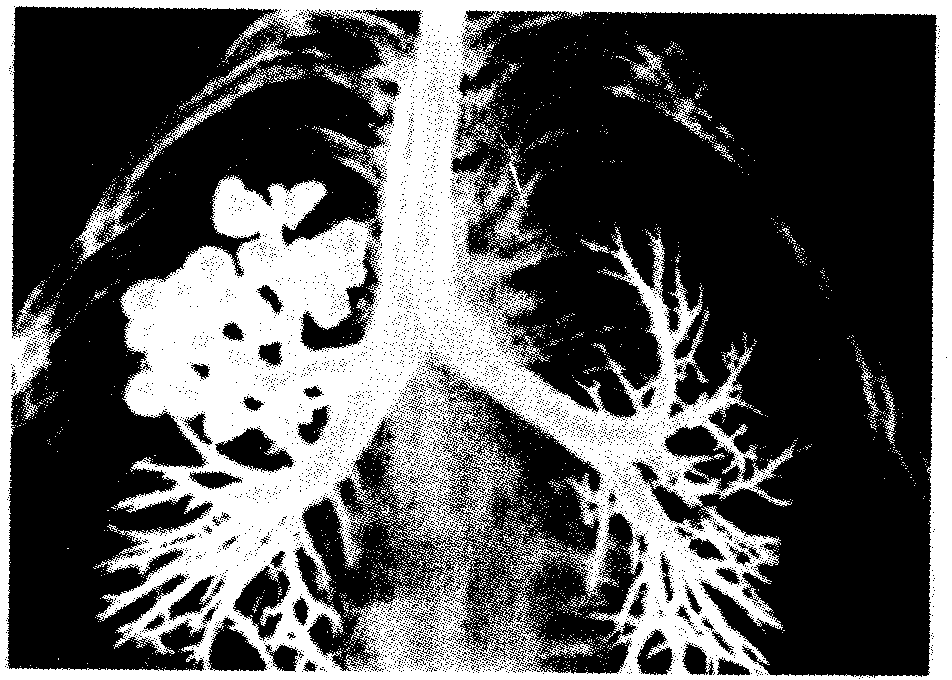
Рис. 14.26. Кистозная гипоплазия легкого
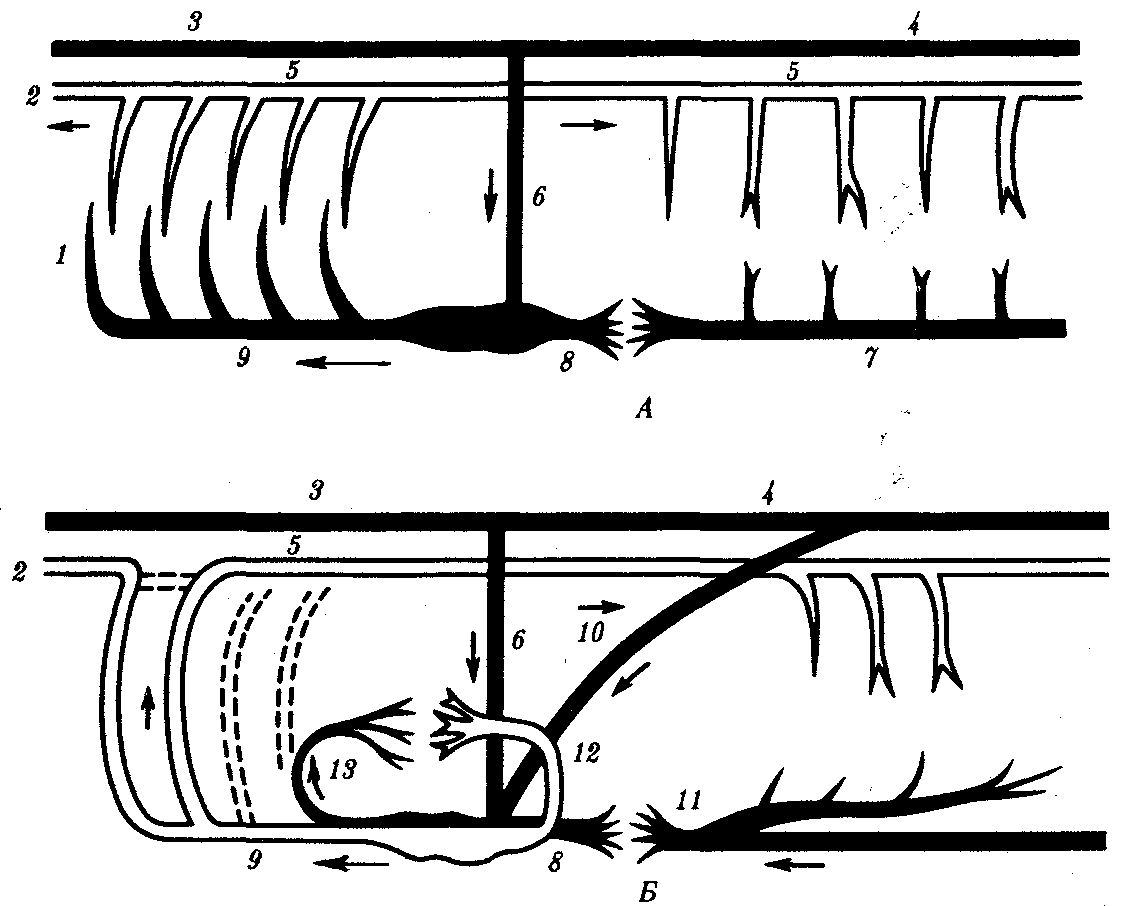
Рис. 14.27. Строение кровеносной системы водных (А) и наземных (Б) позвоночных:
1—жаберные артерии, 2—сонная артериям—передняя кардинальная вена, 4— задняя кардинальная вена, 5—спинная аорта, 6—кювьеров проток, 7—подкишечная вена, 8—печеночная вена, 9—брюшная аорта, 10—задняя (нижняя) полая вена, 11—воротная вена печени, 12—легочная вена, 13—легочная артерия; стрелками указано направление движения крови
14.4.1. Эволюция общего плана строения
кровеносной системы хордовых
У ланцетника кровеносная система наиболее проста (рис. 14.28, А). Круг кровообращения один. По брюшной аорте венозная кровь поступает в приносящие жаберные артерии, которые по количеству соответствуют числу межжаберных перегородок (до 150 пар), где и обогащается кислородом.
По выносящим жаберным артериям кровь поступает в корни спинной аорты, расположенные симметрично с двух сторон тела. Они продолжаются как вперед, неся артериальную кровь к головному мозгу, так и назад. Передние ветви этих двух сосудов являются сонными артериями. На уровне заднего конца глотки задние ветви образуют спинную аорту, которая разветвляется на многочисленные артерии, направляющиеся к органам и распадающиеся на капилляры.
После тканевого газообмена кровь поступает в парные передние или задние кардинальные вены, расположенные симметрично. Передняя и задняя кардинальные вены с каждой стороны впадают в кювьеров проток. Оба кювьеровых протока впадают с двух сторон в брюшную аорту. От стенок пищеварительной системы венозная кровь оттекает по воротной вене печени в печеночный вырост, где формируется система капилляров. Затем капилляры вновь собираются в венозный сосуд — печеночную вену, по которой кровь поступает в брюшную аорту.
Таким образом, несмотря на простоту кровеносной системы в целом, уже у ланцетника имеются основные магистральные артерии, характерные для позвоночных, в том числе для человека: это брюшная аорта, преобразующаяся позже в сердце, восходящую часть дуги аорты и корень легочной артерии; спинная аорта, становящаяся позже собственно аортой, и сонные артерии. Основные вены, имеющиеся у ланцетника, также сохраняются у более высокоорганизованных животных. Так, передние кардинальные вены станут позже яремными венами, правый кювьеров проток преобразуется в верхнюю полую вену, а левый, сильно редуцировавшись, — в коронарный синус сердца. Для того чтобы понять, как это происходит, необходимо сопоставить кровеносные системы всех классов позвоночных животных.
Более активный образ жизни рыб предполагает более интенсивный метаболизм. В связи с этим на фоне олигомеризации их артериальных жаберных дуг в конечном счете до четырех пар в них отмечается высокая степень дифференцировки: жаберные сосуды распадаются на капилляры, пронизывающие жаберные лепестки. В процессе интенсификации сократительной функции брюшной аорты часть ее преобразовалась в двухкамерное сердце, состоящее из предсердия и желудочка и располагающееся под нижней челюстью, рядом с жаберным аппаратом. В остальном кровеносная система рыб соответствует строению ее у ланцетника (рис. 14.28, Б).
В связи с выходом земноводных на сушу и появлением легочного дыхания у них возникает два круга кровообращения. Соответственно этому в строении сердца и артерий появляются приспособления, направленные на разделение артериальной и венозной крови. Перемещение земноводных в основном за счет парных конечностей, а не хвоста обусловливает изменения в венозной системе задней части туловища.
Сердце амфибий расположено каудальнее, чем у рыб, рядом с легкими; оно трехкамерное, но, как и у рыб, от правой половины единственного желудочка начинается единственный сосуд — артериальный конус, разветвляющийся последовательно на три пары сосудов:
кожно-легочные артерии, дуги аорты и сонные артерии. Как и у всех более высокоорганизованных классов, в правое предсердие впадают вены большого круга, несущие венозную кровь, в левое— малого с артериальной кровью. При сокращении предсердий в желудочек, внутренняя стенка которого снабжена большим количеством мышечных перекладин, одновременно попадают обе порции крови. Полного их смешения из-за своеобразного строения стенки желудочка не происходит, поэтому при его сокращении первая порция венозной крови поступает в артериальный конус и с помощью спирального клапана, находящегося там, направляется в кожно-легочные артерии. Кровь из середины желудочка, смешанная, поступает таким же образом в дуги аорты, а оставшееся небольшое количество артериальной крови, последней попадающей в артериальный конус, направляется в сонные артерии.
Две дуги аорты, несущие смешанную кровь, огибают сердце и пищевод сзади, образуя спинную аорту, снабжающую все тело, кроме головы, смешанной кровью. Задние кардинальные вены сильно редуцируются и собирают кровь только с боковых поверхностей туловища. Функционально их замещает возникшая заново задняя полая вена, собирающая кровь в основном из задних конечностей. Она располагается рядом со спинной аортой и, находясь позади печени, вбирает в себя печеночную вену, которая у рыб впадала непосредственно в венозный синус сердца. Передние кардинальные вены, обеспечивая отток крови от головы, называют теперь яремными венами, а кювьеровы потоки, в которые они впадают вместе с подключичными венами, — передними полыми венами (рис. 14.28, В).
В кровеносной системе пресмыкающихся возникают следующие прогрессивные изменения: в желудочке их сердца имеется неполная перегородка, затрудняющая смешение крови, поступающей из правого и левого предсердий; от сердца отходит не один, а три сосуда, образовавшихся в результате разделения артериального ствола. Из левой половины желудочка начинается правая дуга аорты, несущая артериальную кровь, а из правой — легочная артерия с венозной кровью. Из середины желудочка, в области неполной перегородки, начинается левая дуга аорты со смешанной кровью. Обе дуги аорты, как и у предков, срастаются позади сердца, трахеи и пищевода в спинную аорту, кровь в которой смешанная, но более богата кислородом, чем у земноводных, в связи с тем что до слияния сосудов только по левой дуге течет смешанная кровь. Кроме того, сонные и подключичные артерии с обеих сторон берут начало от правой дуги аорты, в результате чего артериальной кровью снабжается не только голова, но и передние конечности. В связи с появлением шеи сердце располагается еще более каудально, чем у земноводных. Венозная система пресмыкающихся принципиально не отличается от системы вен земноводных (рис. 14.28, Г).
Прогрессивные изменения кровеносной системы млекопитающих сводятся к полному разделению венозного и артериального кровотоков. Это достигается, во-первых, завершенной четырехкамерностью сердца и, во-вторых, редукцией правой дуги аорты и сохранением только левой, начинающейся от левого желудочка. В результате все органы млекопитающих снабжаются артериальной кровью. В венах большого круга кровообращения также обнаруживаются прогрессивные изменения: возникла безымянная вена, объединяющая левые яремную и подключичную вены с правыми, в результате чего остается лишь одна передняя полая вена, располагающаяся справа. Левый кювьеров проток в виде рудиментарного сосуда sinus coronarius теперь собирает венозную кровь только от миокарда, а непарная и полунепарная вены — рудименты задних кардинальных вен, имеют существенное значение в основном в случаях формирования обходных путей венозного оттока через кава-кавальные анастомозы, формируемые ими (рис. 14.28, Д).
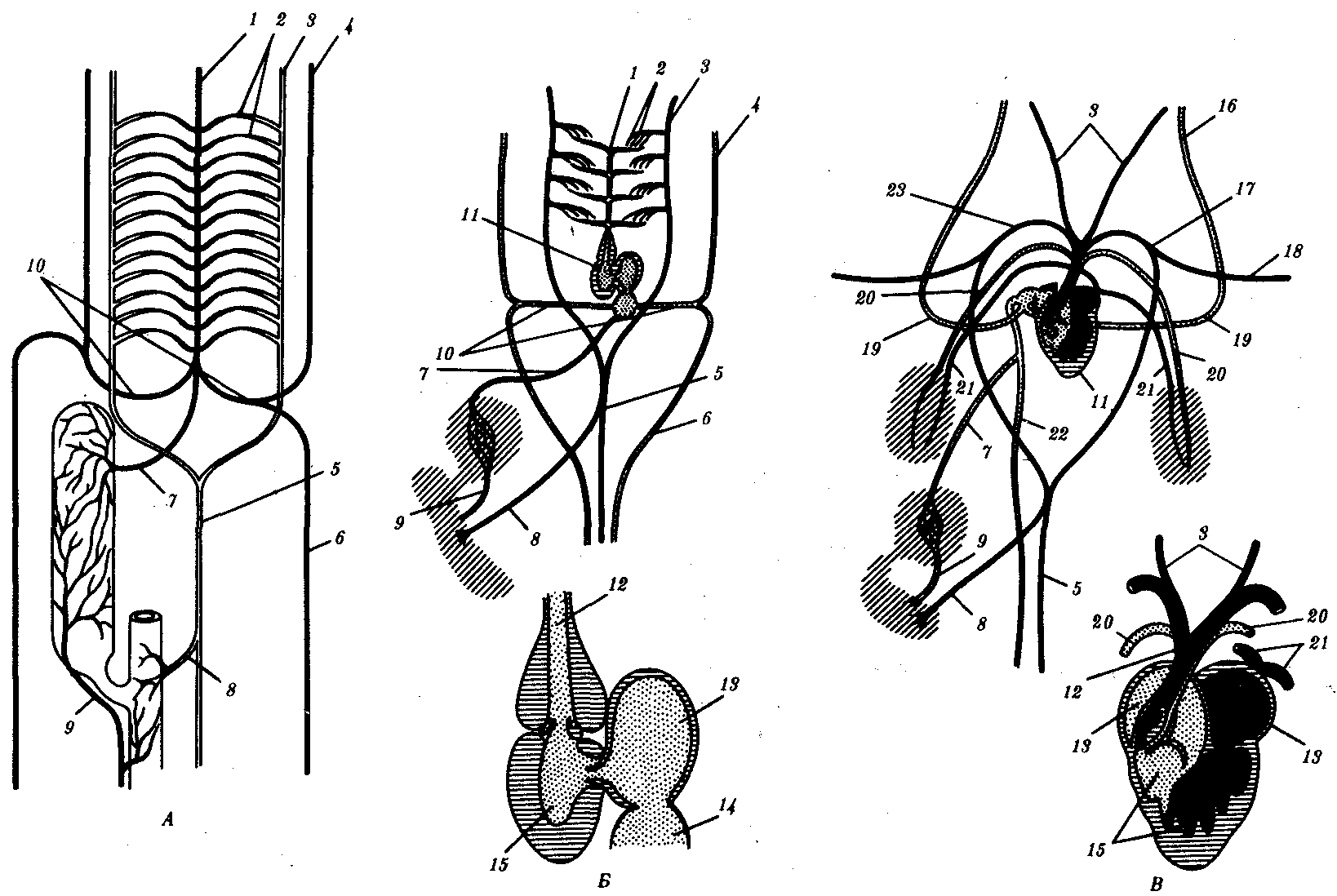
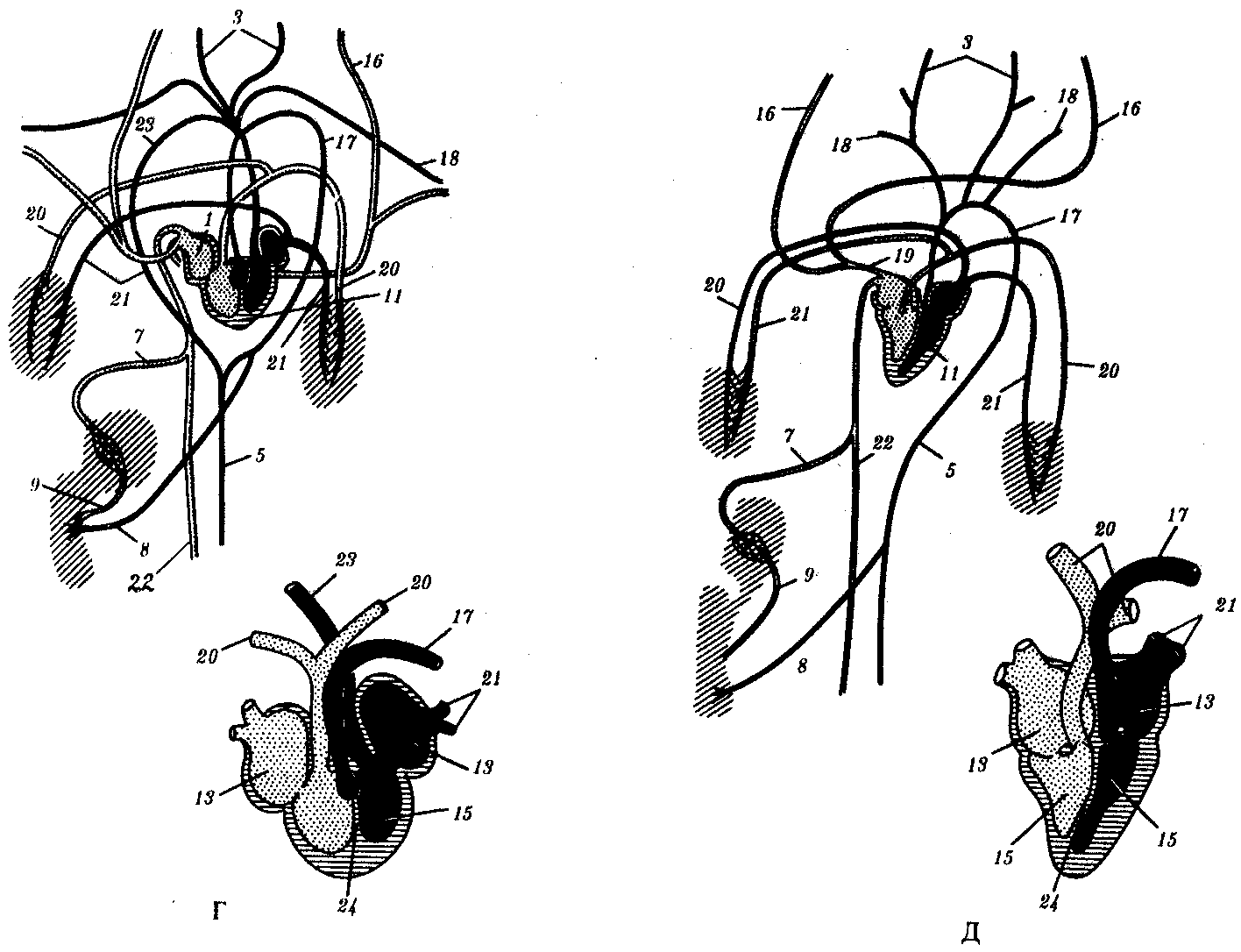
Рис 14.28. Строение кровеносной системы и сердца хордовых животных.
А—ланцетника; Б—рыбы; В—земноводного; Г—пресмыкающегося; Д—млекопитающего:
1—брюшная аорта, 2—жаберные артерии, 3—сонная артерия, 4—передняя кардинальная вена, 5—спинная аорта, 6—задняя кардинальная вена, 7— печеночная вена, 8—чревная артерия, 9—воротная вена печени, 10— кювьеровы протоки, 11—сердце, 12—артериальный конус, 13—предсердие, 14—венозный синус, 15—желудочек, 16—яремная вена, 17—левая дуга аорты, 18—подключичная артерия, 19—передняя полая вена, 20—легочная артерия, 21—легочная вена, 22—задняя полая вена, 23—правая дуга аорты, 24— межжелудочковая перегородка
В эмбриональном развитии млекопитающих и человека рекапитулируют закладки сердца и основных кровеносных сосудов предковых классов.
Сердце закладывается на первых этапах развития в виде недифференцированной брюшной аорты, которая за счет изгибания, появления в просвете перегородок и клапанов, становится последовательно двух-, трех- и четырехкамерным (рис. 14.29). Однако рекапитуляции здесь неполны в связи с тем, что межжелудочковая перегородка млекопитающих формируется иначе и из другого материала по сравнению с рептилиями. Поэтому можно считать, что четырехкамерное сердце млекопитающих формируется на базе трехкамерного сердца, а межжелудочковая перегородка является новообразованием, а не результатом доразвития перегородки пресмыкающихся. Таким образом, в филогенезе сердца позвоночных проявляется девиация: в процессе морфогенеза этого органа у млекопитающих рекапитулируют ранние филогенетические стадии, а затем развитие его идет в ином направлении, характерном лишь для этого класса.
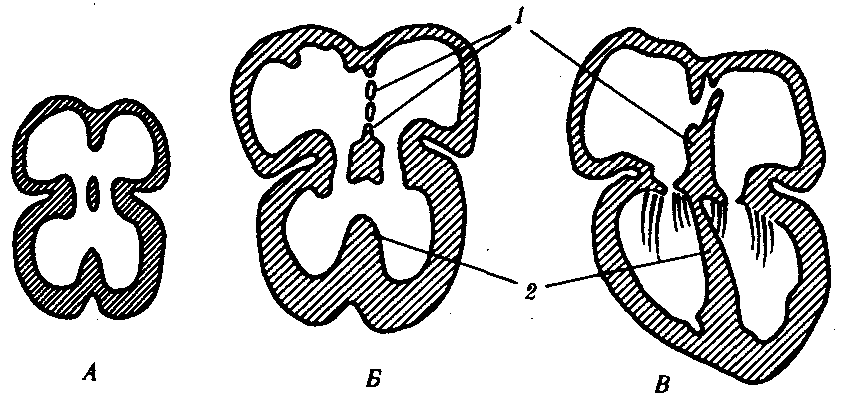
Рис. 14.29. Развитие перегородок сердца у зародыша человека в возрасте 30 сут (А) 35 сут (Б) и 2 мес (Д): 1—межпредсердная, 2—межжелудочковая
Интересно, что место закладки и положение сердца в филогенетическом ряду позвоночных полностью рекапитулируют у млекопитающих и человека. Так, закладка сердца у человека осуществляется на 20-е сутки эмбриогенеза, как у всех позвоночных, позади головы. Позже за счет изменения пропорций тела, появления шейной области, смещения легких в грудную полость осуществляется и перемещение сердца в переднее средостение.
Нарушения развития сердца могут выражаться как в возникновении аномалий строения, так и места его положения. Возможно сохранение к моменту рождения двухкамерного сердца. Этот порок совершенно не совместим с жизнью.
Чаще встречаются дефекты межпредсердной перегородки (1 случай на 1000 рождений), межжелудочковой перегородки (2,5—5 случаев на 1000 рождений), вплоть до трехкамерного сердца с одним общим желудочком (см. рис. 14.31, В). Известен и такой порок, как шейная эктопия сердца, при которой оно находится в шейной области. Этот порок связывают с задержкой сердца в области его первоначальной закладки. При этом ребенок обычно погибает сразу после рождения. Перечисленные пороки сердца наиболее часто встречаются не в изолированном виде, а в комплексе с другими аномалиями сердца, сосудов, а нередко и других органов. Это свидетельствует о том, что в морфо-генезе сердца большое значение имеют онтогенетические корреляции (см. § 13.4). Состояние больных при таких пороках зависит от того, насколько сильно нарушается гемодинамика и осуществляется смешение крови в кровеносном русле.
14.4.2. Филогенез артериальных жаберных дуг
В связи с тем что основные артериальные сосуды у млекопитающих и человека формируются на базе закладок жаберных артерий, проследим их эволюцию в филогенетическом ряду позвоночных (рис. 14.30). В эмбриогенезе абсолютного большинства позвоночных закладывается шесть пар артериальных жаберных дуг, соответствующих шести парам висцеральных дуг черепа. В связи с тем что две первые пары висцеральных дуг включаются в состав лицевого черепа, две первые артериальные жаберные дуги быстро редуцируются. Оставшиеся четыре пары функционируют у рыб как жаберные артерии. У наземных позвоночных 3-я пара жаберных артерий теряет связь с корнями спинной аорты и несет кровь к голове, становясь сонными артериями. Сосуды 4-й пары достигают наибольшего развития и вместе с участком корня спинной аорты во взрослом состоянии становятся дугами аорты — основными сосудами большого круга кровообращения.
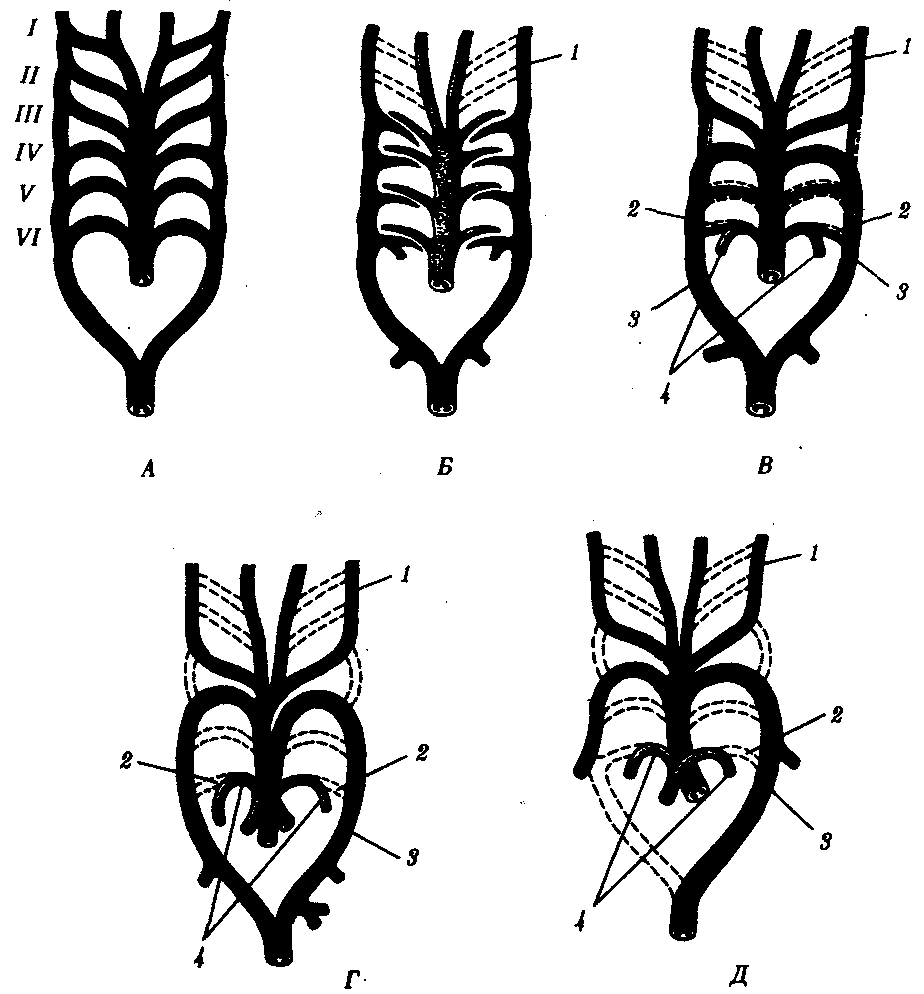
Рис. 14.30. Эволюция артериальных жаберных дуг позвоночных. А—закладка в эмбриогенезе; Б—жаберные дуги рыб; В—жаберные дуги хвостатых земноводных- Г—жаберные дуги пресмыкающихся; Д—жаберные дуги млекопитающих:
I—VI - жаберные дуги; 1—сонные артерии, 2—боталлов проток, 3—артериальные дуги большого круга кровообращения, 4—легочные артерии
У земноводных и пресмыкающихся оба сосуда развиты и принимают участие в кровообращении. У млекопитающих также закладываются оба сосуда 4-й пары, а позже правая дуга аорты редуцируется таким образом, что от нее остается лишь небольшой рудимент— плечеголовной ствол. Пятая пара артериальных дуг в связи с тем, что она функционально дублирует четвертую, редуцируется у всех наземных позвоночных, кроме хвостатых амфибий. Шестая пара, которая снабжает венозной кровью кроме жабр еще и плавательный пузырь, у кистеперых рыб становится легочной артерией.
В эмбриогенезе человека рекапитуляции артериальных жаберных дуг происходят с особенностями: все шесть пар дуг никогда не существуют одновременно. В то время, когда две первые дуги закладываются, а затем перестраиваются, последние пары сосудов еще не начинают формироваться. Кроме того, пятая артериальная дуга уже закладывается в виде рудиментарного сосуда, присоединенного обычно к 4-й паре, и редуцируется очень быстро.
Из атавистических пороков развития сосудов, развивающихся из артериальных жаберных дуг, остановимся на следующих: с частотой 1 случай на 200 вскрытии детей, умерших от врожденных пороков сердца, встречается персистирование обеих дуг аорты 4-й пары. При этом обе дуги, так же как у земноводных или пресмыкающихся, срастаются позади пищевода и трахеи, образуя нисходящую часть спинной аорты. Порок проявляется нарушением глотания и удушьем. Несколько чаще (2,8 случая на 200 вскрытии) встречается нарушение редукции правой дуги аорты с редукцией левой. Эта аномалия часто клинически не проявляется.
Наиболее частый порок (0,5—1,2 случая на 1000 новорожденных) — персистирование артериального, или боталлова, протока (рис. 14.31, А) представляющего собой часть корня спинной аорты между 4-й и'6-и парами артерий слева. Проявляется сбросом артериальной крови из большого круга кровообращения в малый. Очень тяжелый порок развития — персистирование первичного эмбрионального ствола, в результате которого из сердца выходит только один сосуд, располагающийся обычно над дефектом в межжелудочковой перегородке (рис. 14.31, Б). Он обычно заканчивается смертью ребенка. Нарушение дифференцировки первичного эмбрионального ствола может привести к такому пороку развития, как транспозиция сосудов — отхождение аорты от правого желудочка, а легочного ствола — от левого, что встречается в 1 случае на 2500 новорожденных. Этот порок обычно несовместим с жизнью.
Рекапитуляции проявляются и в эмбриональном развитии крупных вен человека. При этом возможно формирование атавистических пороков развития. Среди пороков развития венозного русла укажем на возможность персистирования двух верхних полых вен. Если обе они впадают в правое предсердие, аномалия клинически не проявляется. При впадении левой полой вены в левое предсердие происходит сброс венозной крови в большой круг кровообращения. Иногда обе полые вены впадают в левое предсердие. Такой порок несовместим с жизнью. Данные аномалии встречаются с частотой 1% от всех врожденных пороков сердечно-сосудистой системы.
Очень редкая врожденная аномалия — неразвитие нижней полой вены. Отток крови от нижней части туловища и ног осуществляется в этом случае через коллатерали непарной и полунепарной вен, являющихся рудиментами задних кардинальных вен.
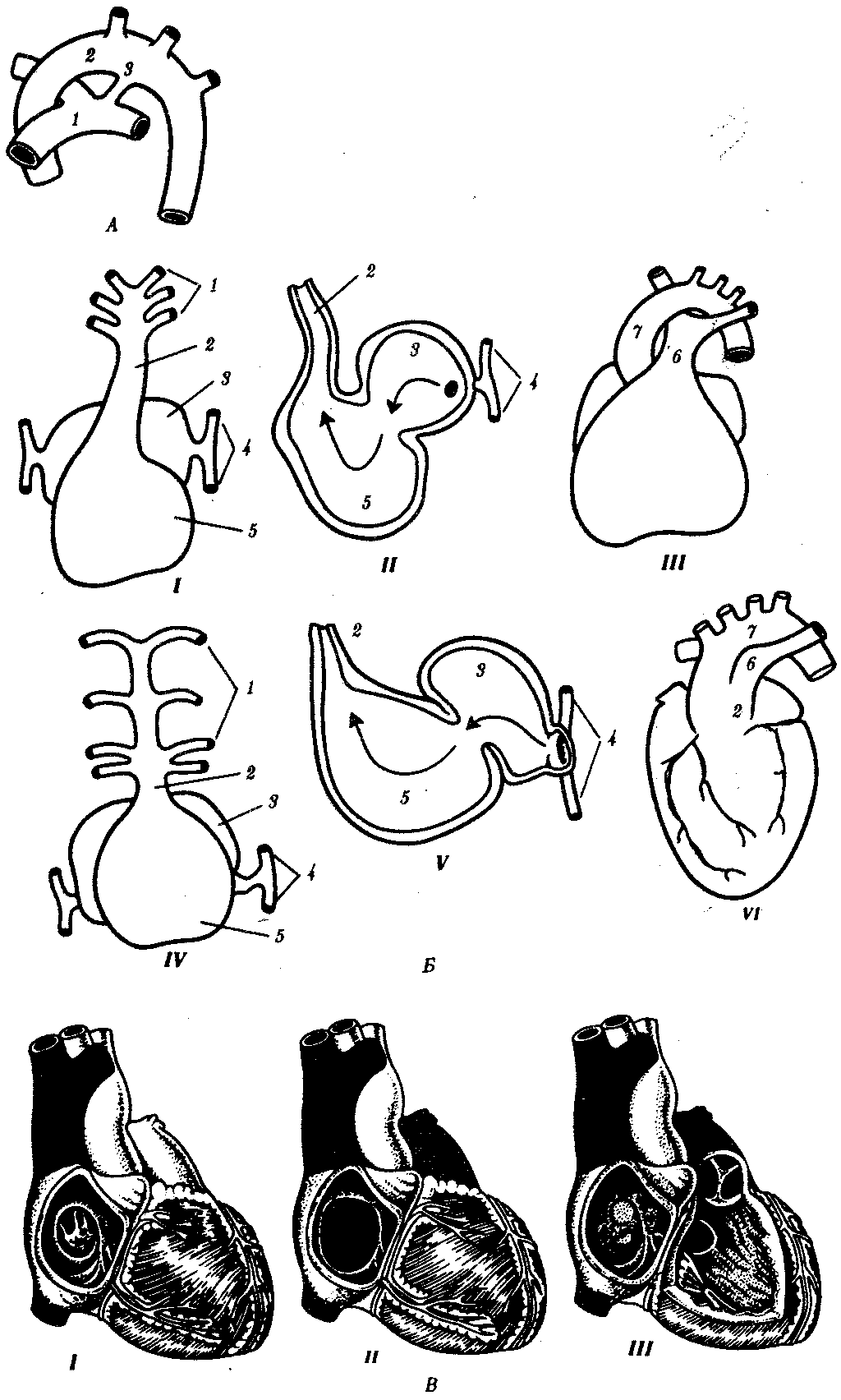
Рис. 14.31. Врожденные пороки развития сердца и сосудов у человека:
А — персистирование артериального (боталлова) протока у человека:
1—легочный ствол, 2—левая дуга аорты, 3—артериальный (боталлов) проток;
Б—онтофилогенетические основы формирования пороков развития сердца и отходящих от него сосудов у человека: I—сердце зародыша человека в возрасте 5 недель; II—то же, в разрезе III—разделение артериального конуса у зародыша человека в возрасте 6 недель, IV—сердце рыбы, V—то же, в разрезе, VI—персистирование неразделенного артериального конуса у человека: 1—артериальные жаберные дуги, 2—артериальный конус, 3—предсердие, 4—кардинальные вены, 5—желудочек, 6—общая легочная артерия, 7—левая дуга аорты;
В — аномалии межпредсердной (I, II) и межжелудочковой (III) перегородок
14.5. МОЧЕПОЛОВАЯ СИСТЕМА
Выделительная и половая системы в целом выполняют разные функции. Однако их рассматривают в едином комплексе в связи с единством эмбрионального развития и первичной функциональной связью со вторичной полостью тела — целбмом. В эмбриогенезе закладка мочеполовой системы — нефрогонотом — формируется в области ножки сомита в непосредственном контакте с целомом. Половые железы у всех хордовых животных располагаются в целоме. Продукты диссимиляции у всех целоми-ческих животных, в том числе и низших хордовых, также поступают в целом. Наиболее простой путь выведения во внешнюю среду как половых клеток, так и продуктов диссимиляции — через общий канал, начинающийся воронкой в целоме и заканчивающийся выделительной порой на покровах. Таковы взаимоотношения половых желез и выделительных каналов у многих беспозвоночных и, вероятно, у древних предков хордовых (рис. 14.32).
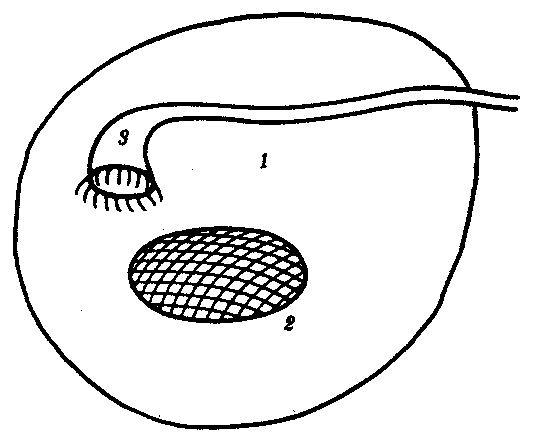
Рис. 14.32. Взаимоотношения целома (1), половой железы (2) и общего выделительного канала (3)
14.5.1. Эволюция почки
Органами выделения позвоночных являются почки — парные компактные органы, структурная единица которых представлена нефроном. В наиболее примитивном виде это — воронка, открывающаяся в целом и соединенная с выделительным канальцем, который впадает в общий выводной проток — мочеточник. В филогенезе позвоночных почка прошла три этапа эволюции: предпочка — головная, или пронефрос; первичная почка—туловищная, или мезонефрос, и вторичная почка - тазовая, или метанефрос.
Предпочка полностью развивается и функционирует как самостоятельный орган у личинок рыб и земноводных. Она находится на переднем конце тела, состоит из 2—12 нефронов, воронки которых открыты в целом, а выводные канальцы впадают в пронефрический канал, который соединен с клоакой. Предпочка имеет сегментарное строение. Продукты диссимиляции фильтруются в целом из кровеносных сосудов, которые поблизости от нефронов формируют клубочки (рис. 14.33, А).
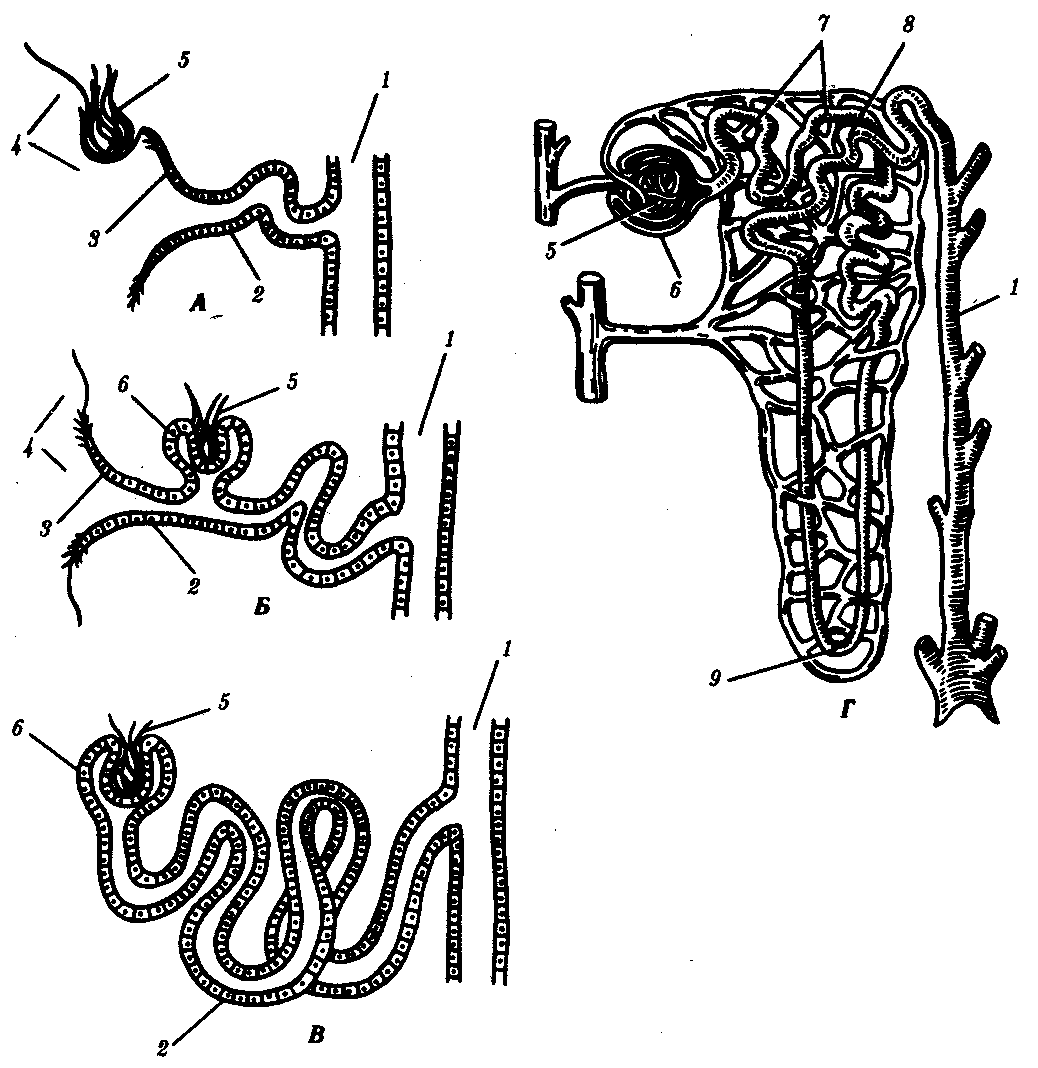
Рис. 14.33. Эволюция тефрона. А—предпочка; Б, В—первичная почка; Г—вторичная почка:
1—собирательная трубочка, 2—выделительный канадец, 3—нефростом, 4—целом, 5—капиллярный клубочек, 6—капсула, 7, 8—извитой канадец, 9—петля нефрона
У взрослых рыб и земноводных кзади от предпочек, в туловищных сегментах тела, формируются первичные почки, содержащие до нескольких сотен нефронов. В ходе онтогенеза нефроны увеличиваются в количестве за счет их почкования друг от друга с последующей дифференцировкой. Они вступают в связь с кровеносной системой, формируя капсулы почечных клубочков. Капсулы имеют вид двустенных чаш, в которых располагаются сосудистые клубочки, благодаря чему продукты диссимиляции могут поступать из крови непосредственно в нефрон. Некоторые нефроны первичной почки сохраняют связь с целомом через воронки, другие — утрачивают её (рис. 14.33, Б, В).
Выделительные канальцы удлиняются и в них осуществляется обратное всасывание в кровь воды, глюкозы и других веществ, в связи с чем концентрация продуктов диссимиляции в моче повышается. Однако воды с мочой теряется много, поэтому животные, обладающие такой почкой, могут обитать только в водной или влажной среде. Первичная почка сохраняет признаки метамерного строения.
У пресмыкающихся и млекопитающих возникают вторичные почки. Они закладываются в тазовом отделе тела и содержат сотни тысяч нефронов наиболее совершенного строения. У новорожденного ребенка в почке их насчитывается около 1 млн. Они образуются за счет многократного ветвления развивающихся нефронов. Нефроны не имеют воронки и, таким образом, теряют полностью связь с целомом. Канадец нефрона удлиняется, теснее контактирует с кровеносной системой, а у млекопитающих дифференцируется на проксимальный и дистальный участки, между которыми появляется еще и так называемая петля Генле (рис. 14.33, Г).
Такое строение нефрона обеспечивает не только полноценную фильтрацию плазмы крови в капсуле, но и, что более важно, эффективное обратное всасывание в кровь воды, глюкозы, гормонов, солей и других необходимых организму веществ. В результате концентрация продуктов диссимиляции в моче, выделяемой вторичными почками, велика, а само ее количество — мало. У человека, например, за сутки в капсулах нефронов обеих почек фильтруется около 150 л плазмы крови, а мочи выделяется около 2 л. Это позволяет животным, обладающим вторичными почками, быть более независимыми от водной среды и заселять засушливые участки земли. У пресмыкающихся вторичные почки на протяжении всей жизни сохраняются на месте их первоначальной закладки — в тазовой области. В них прослеживаются черты первичного метамерного строения.
Почки млекопитающих располагаются в поясничной области, и у большинства из них внешняя сегментация не выражена. В онтогенезе человека обнаруживается выраженная рекапитуляция в развитии почки: закладывание вначале про-, затем мезо-, а позже метанефроса. Последний развивается в тазовой области, а затем за счет различий в скоростях роста позвоночника, таза и органов брюшной полости перемещается в поясничную область. У пятинедельного зародыша можно обнаружить сосуществование предпочки, первичной, а также зачатков вторичной почки (рис. 14.34).
На начальных этапах развития почка человека сегментирована. Позже ее поверхность сглаживается и метамерность сохраняется лишь во внутреннем строении в виде почечных пирамид. Пороки развития почек у человека, основанные на их филогенезе, многообразны. Сохранение мезонефроса и одностороннее отсутствие вторичной почки описаны пока только у мышей, хотя в принципе такая аномалия возможна и у человека. Относительно часто встречается сегментированная вторичная почка, имеющая один или даже несколько мочеточников; возможно и полное ее удвоение. Часто наблюдается тазовое расположение почки, связанное с нарушением ее перемещения на 2—4-м месяцах зародышевого развития (рис. 14.35).
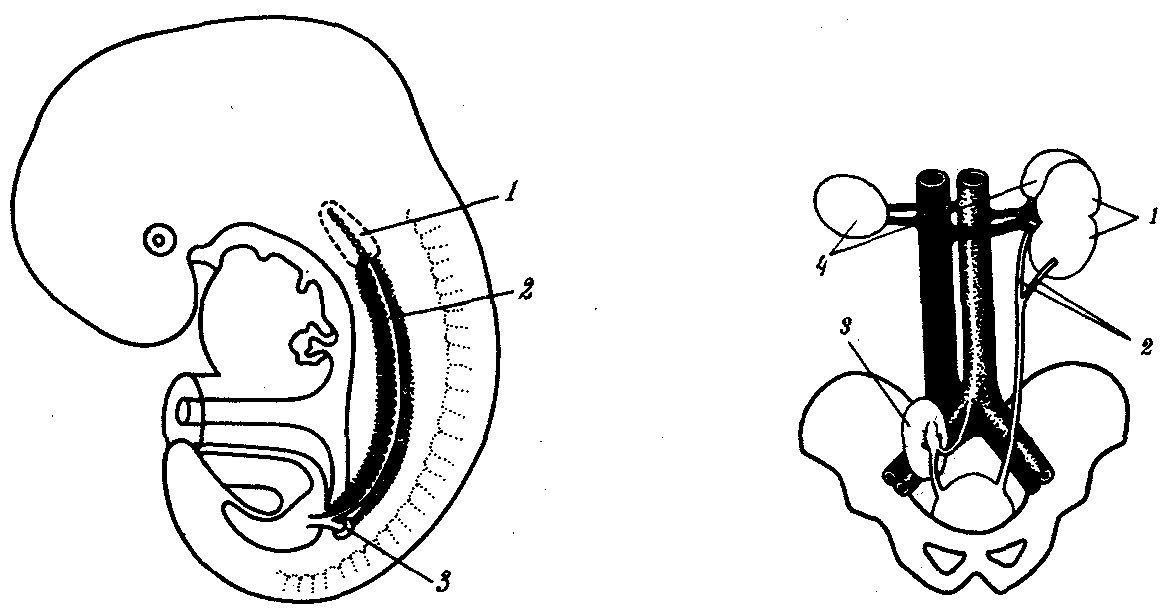
Рис. 14.34. Пятинедельный зародыш человека с тремя поколениями почек:
1—предпочка, 2—первичная почка, 3—вторичная почка
Рис. 14.35. Онтофилогенетически обусловленные пороки развития почек:
1—удвоение почки, 2—удвоенный мочеточник, 3—тазовая эктопия почки, 4—надпочечники
14.5.2. Эволюция половых желез
Половые железы у всех позвоночных развиваются в виде парных складок части нефрогонотома в области ножки сомита. Половые складки вдаются в полость тела и оказываются подвешенными на брыжейке. Первичные половые клетки обособляются у зародышей очень рано — уже на стадии гаструляции. Вначале они обнаруживаются в составе презумптивной эктодермы головного конца эмбриона, затем попадают в энтодерму, откуда активно перемещаются в половые складки. Здесь дифференцирующийся эпителий половой железы, включающий в себя первичные половые клетки, объединяется с соединительнотканной стромой в виде шнуров. Такая гонада индифферентна в половом отношении и может развиваться в дальнейшем как в семенник, так и в яичник в зависимости от генетических и эпигенетических факторов дифференцировки пола.
У хрящевых рыб строение половых желез самок и самцов в значительной степени сходно. У всех остальных позвоночных яичник всегда имеет фолликулярное строение, т.е. содержит пузырьки — фолликулы, в каждом из которых находится одна будущая яйцеклетка. При созревании яйцеклетки стенка фолликула разрывается, и она поступает вначале в брюшную полость, а затем в яйцевод. Семенники содержат семенные трубочки, которые соединены с семявыносящими каналами, по которым зрелые сперматозоиды выходят во внешнюю среду.
Индифферентность развивающейся половой железы позвоночных называют первичным гермафродитизмом. Он эволюционно связан, вероятно, с гермафродитизмом древних предков позвоночных. Доказательством этого может служить наличие у наиболее примитивных современных позвоночных — круглоротых — таких половых желез, передняя часть которых является яичником, а задняя — семенником. У самок некоторых земноводных на протяжении всей жизни сохраняется рудиментарный участок индифферентной половой железы. Зародыш человека до определенного возраста имеет недифференцированные половые железы, которые в зависимости от различных факторов (см. разд. 6.1.2) становятся либо семенниками, либо яичниками. Нарушение их дифференцировки может привести к возникновению ovotestis, в котором сочетаются элементы семенника и яичника. У детей, имеющих ovotestis, обнаруживаются признаки гермафродитизма и в наружных половых органах.
У всех позвоночных с непостоянной температурой тела половые железы находятся в брюшной полости. У большинства млекопитающих мужские гонады перемещаются через паховой канал в мошонку, где температура всегда несколько ниже. Вероятно, пониженная температура более благоприятна для сперматогенеза, обеспечивая более низкий уровень спонтанных мутаций в созревающих сперматозоидах.
У человека семенники, закладываясь в брюшной полости, перемещаются через паховой канал и к 8-му месяцу внутриутробного развития оказываются в мошонке. У 2,2 % мальчиков пубертатного возраста обнаруживаются различные формы крипторхизма — неопущения яичек, которые при этом обычно недоразвиты, а часть семявыносящих канальцев заменена соединительной тканью. Для предотвращения бесплодия таким мальчикам необходимо хирургическое низведение яичек в мошонку в раннем возрасте.
14.5.3. Эволюция мочеполовых протоков
В эмбриогенезе всех позвоночных при развитии предпочки вдоль тела, от головного конца к клоаке, закладывается канал, по которому продукты диссимиляции из нефронов поступают во внешнюю среду. Это пронефрический канал. При развитии первичной почки этот канал либо расщепляется на два канала, идущих параллельно, либо второй канал образуется в продольном утолщении стенки первого. Один из них — вольфов — вступает в связь с нефронами первичной почки. Другой — мюллеров — срастается передним концом с одним из нефронов предпочки и образует яйцевод, открывающийся передним концом в целом широкой воронкой, а задним — впадающий в клоаку.
Вне зависимости от пола у всех позвоночных обязательно формируются как вольфов, так и мюллеров каналы, однако судьба их различна как у разных полов, так и у представителей разных классов. У самок рыб и земноводных вольфов канал всегда выполняет функцию мочеточника, а мюллеров — яйцевода. У самцов мюллеров канал редуцируется и обе функции — половую и выделительную — выполняет вольфов канал. Семенные канальцы при этом впадают в почку, а сперматозоиды при оплодотворении поступают в воду вместе с мочой.
У пресмыкающихся и млекопитающих большая часть вольфова канала не принимает участия в выведении мочи и только его наиболее каудальная часть в области впадения в клоаку образует выпячивание, становящееся мочеточником вторичной почки. Сам же вольфов канал у самцов выполняет функцию семяизвергательного канала. Мюллеров канал у них подвергается редукции. У самок вольфов канал редуцируется (за исключением его каудальной части, формирующей мочеточник), а мюллеров — становится яйцеводом (рис. 14.36). У плацентарных млекопитающих мюллеров канал дифференцируется на собственно яйцевод, матку и влагалище. Будучи парным образованием, как и все элементы половой системы, мюллеров канал сохраняет парность строения у яйцекладущих и частично у сумчатых млекопитающих, у которых имеется два влагалища, две матки и два яйцевода. В дальнейшей эволюции происходит срастание мюллеровых каналов с образованием одного влагалища и матки, которая может быть либо двойной, как у многих грызунов, либо двураздельной, как у хищных; либо двурогой, как у насекомоядных и китообразных, либо простой, как у приматов и человека.
Соответственно дифференцировкам мюллерова канала самок у самцов пресмыкающихся и млекопитающих развиваются копулятивные органы. У большинства пресмыкающихся, а также у сумчатых млекопитающих они парные. У плацентарных с одним влагалищем копулятивный орган непарный, но в его развитии обнаруживается срастание парных зачатков.
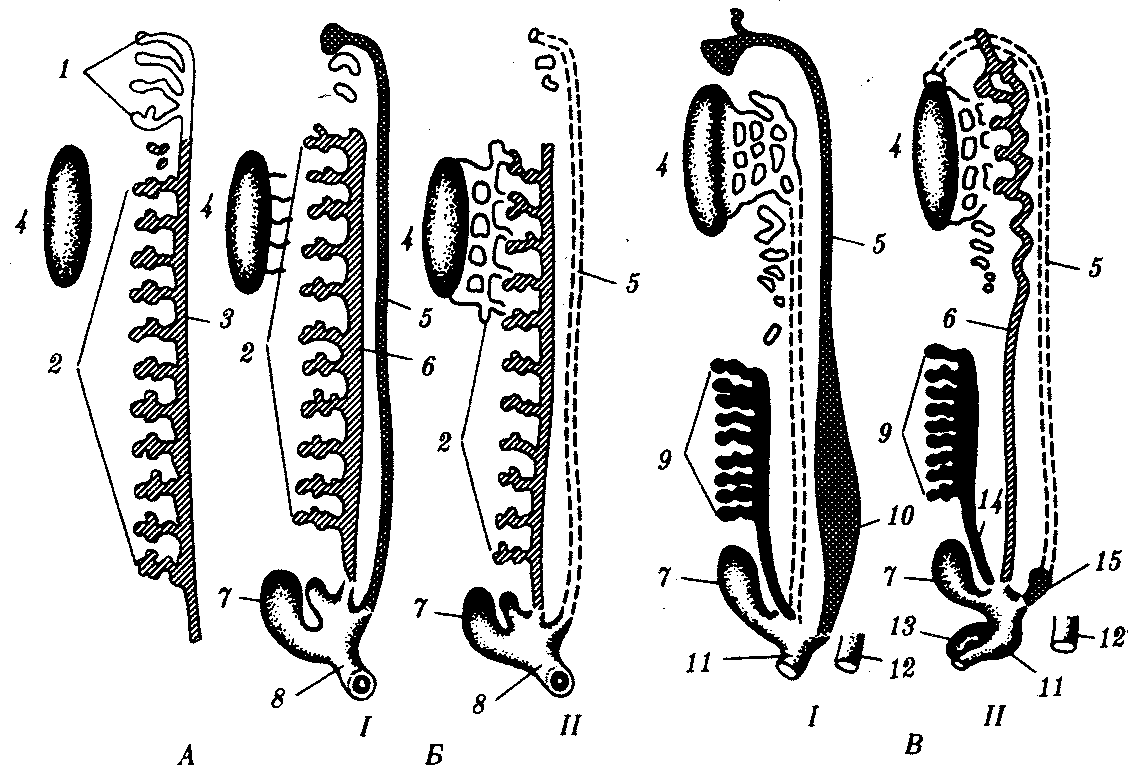
Рис. 14.36. Эволюция почки и мочеполовых каналов.
А — нейтральное зародышевое состояние; Б — анамнии; В — амниоты:
I—самки, II—самцы; 1—предпочка, 2—первичная почка, 3—канал предпочки, 4—половая железа, 5—мюллеров канал, 6—вольфов канал, 7—мочевой пузырь, 8—клоака, 9—вторичная почка, 10—матка, 11—мочеполовой синус, 12—задняя кишка, 13—половой член, 14—мочеточник вторичной почки, 15—мужская «маточка»
В эмбриогенезе человека закладываются парные вольфовы и мюллеровы каналы. Позже в зависимости от пола происходит их редукция. Рудимент мюллерова канала у мужчин располагается в предстательной железе и называется мужской маточкой — utriculus masculinus. Канальцы передней части первичной почки у них вступают в связь с семенниками и преобразуются в придаток семенника — эпидидимис. У плодов женского пола возможно нарушение редукции вольфовых каналов, которые располагаются по бокам от влагалища. Эта аномалия опасна возможностью образования кист и злокачественного перерождения. Распространенными пороками развития являются также различные формы удвоения матки (1 случай на 1000 перинатальных вскрытии). Они развиваются как результат нарушения срастания мюллеровых каналов. Нарушение срастания парных зачатков полового члена в эмбриогенезе человека может привести к формированию такого порока развития, как его удвоение (рис. 14.37).
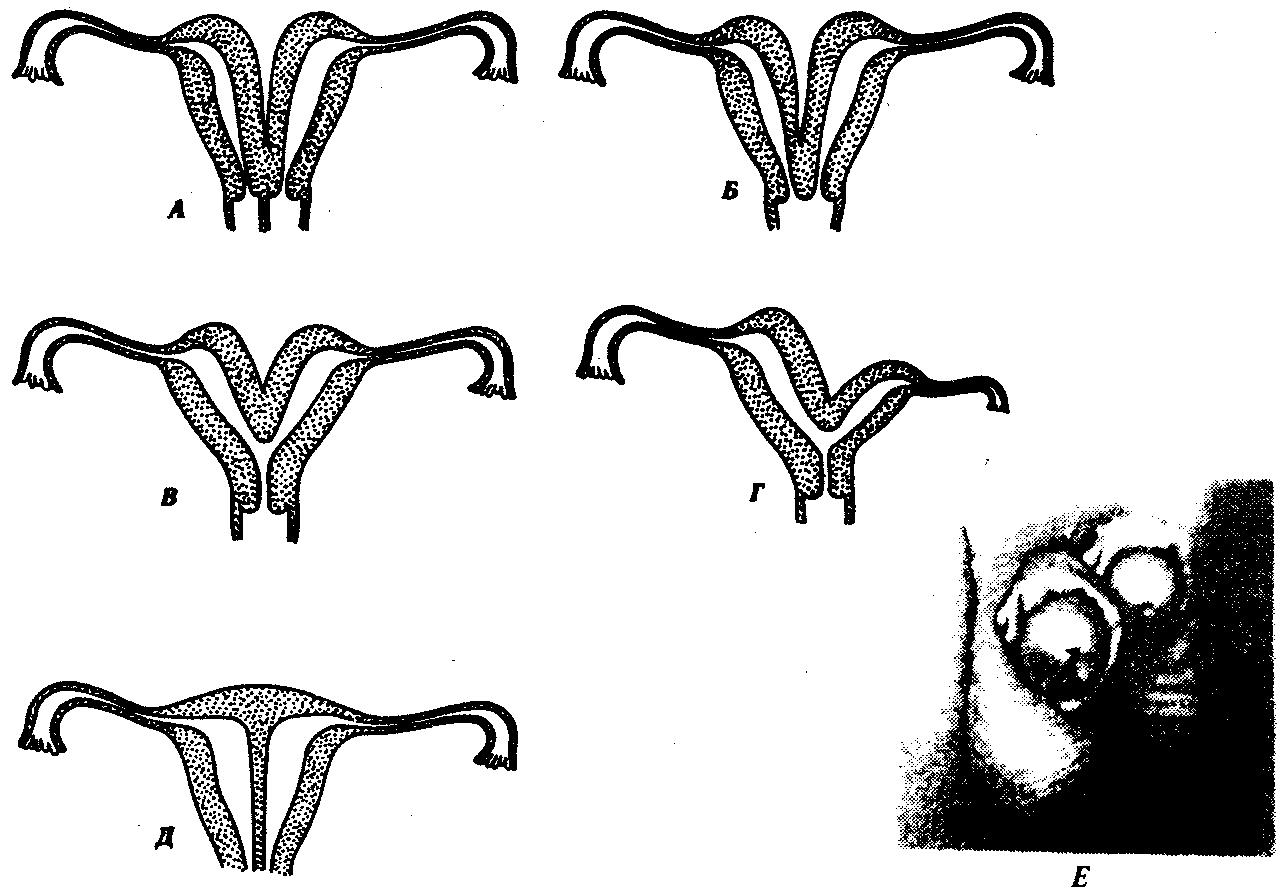
Рис. 14.37. Аномалии матки и полового члена у человека. А—Д—различные формы удвоения матки и влагалища; Е—удвоение полового члена
14.6. ИНТЕГРИРУЮЩИЕ СИСТЕМЫ
У большинства многоклеточных животных, в том числе и у позвоночных, существует две системы регуляции функций, интегрирующие организм в целостную систему, — нервная и эндокринная. Нервная система осуществляет быстрое реагирование организма на изменения условий среды посредством рефлексов. Эндокринная система с помощью гормонов обеспечивает более медленные, но и более стойкие приспособительные реакции. В связи с тесным взаимодействием обеих регуляторных систем в их филогенезе проявляются тесные динамические координации, а в онтогенезе — эргонтические корреляции.
14.6.1. Центральная нервная система
Нервная система хордовых животных, как и у всех многоклеточных, развивается из эктодермы. Она возникла за счет погружения чувствительных клеток, первоначально лежавших на поверхности тела, под его покровы. Это доказывается и сравнительно-анатомическими, и эмбриологическими данными.
Действительно, у наиболее примитивного представителя хордовых — ланцетника — центральная нервная система, состоящая из нервной трубки, сохранила функции органа чувств: среди клеток, лежащих внутри нее, имеются отдельные светочувствительные образования — глазки Гессе. Кроме того, основные дистантные органы чувств — зрения, обоняния и слуха — образуются у всех позвоночных первоначально как выпячивания передней части нервной трубки.
В эмбриогенезе нервная система формируется вначале всегда в виде полосы утолщенной эктодермы на спинной стороне зародыша, которая впячивается под покровы и замыкается в трубку с полостью внутри — невроцелем. У ланцетника это замыкание еще не полное, поэтому нервная трубка выглядит как желобок (рис. 14.38). Передний конец ее расширен. Он гомологичен головному мозгу позвоночных. Большинство клеток нервной трубки ланцетника не являются нервными, они выполняют опорные или рецепторные функции.

Рис 14.38. Нервная трубка ланцетника: 1 — невроцель, 2 — глазки Гессе
У всех позвоночных центральная нервная система является производной нервной трубки, передний конец которой становится головным мозгом, а задний — спинным. Образование головного мозга называют кефализацией. Она связана с усилением двигательной активности позвоночных и необходимостью постоянного анализа раздражении, приходящих из внешней среды, в первую очередь с переднего конца тела. Этот процесс сопровождается также дифференциацией органов чувств, особенно дистантных — обоняния, зрения и слуха. Совместная эволюция органов чувств и головного мозга приводит к возникновению динамических координации между обонятельными рецепторами и передним мозгом, зрительными — и средним, слуховыми — и задним. Видимо, поэтому головной мозг всех современных позвоночных животных в эмбриогенезе закладывается вначале из трех мозговых пузырей — переднего, среднего и заднего — и только позже дифференцируется на пять отделов. Вероятно, предки позвоночных имели более простой головной мозг, развивающийся на основе трех мозговых пузырей.
Головной мозг современных взрослых позвоночных всегда состоит из пяти отделов: переднего, промежуточного, среднего, заднего и продолговатого. Внутри головного и спинного мозга расположена общая полость, соответствующая невроцелю. В спинном мозге это спинномозговой канал, а в головном — желудочки мозга. Ткань мозга состоит из серого вещества (скопления нервных клеток) и белого (отростков нервных клеток).
Во всех отделах головного мозга различают мантию, располагающуюся над желудочками, и основание, лежащее под ними. В прогрессивной эволюции головного мозга проявляется постепенное усиление роли его передних отделов и мантии по сравнению с задними и основанием.
У рыб головной мозг в целом невелик. Слабо развит его передний отдел. Передний мозг не разделен на полушария. Крыша его тонкая, состоит только из эпителиальных клеток и не содержит нервной ткани. Основание переднего мозга включает полосатые тела, от него отходят обонятельные доли. Функционально передний мозг является высшим обонятельным центром.
В промежуточном мозге, с которым связаны эпифиз и гипофиз, расположен гипоталамус, являющийся центральным органом эндокринной системы. Средний мозг рыб наиболее развит. Он состоит из двух полушарий и служит высшим зрительным центром. Кроме того, он представляет собой высший интегрирующий отдел головного мозга. Задний мозг содержит мозжечок, осуществляющий регуляцию координации движений. Он развит очень хорошо в связи с перемещением рыб в трехмерном пространстве. Продолговатый мозг обеспечивает связь высших отделов головного мозга со спинным и содержит центры дыхания и кровообращения. Головной мозг такого типа, в котором высшим центром интеграции функций является средний мозг, называют ихтиопсидным.
У земноводных головной мозг также ихтиопсидный. Однако передний мозг их имеет большие размеры и разделен на полушария. Крыша его состоит из нервных клеток, отростки которых располагаются на поверхности. Как и у рыб, больших размеров достигает средний мозг, также представляющий собой высший интегрирующий центр и центр зрения. Мозжечок несколько редуцирован в связи с примитивным характером движений.
Условия наземного существования пресмыкающихся требуют более сложной морфофункциональной организации мозга. Передний мозг — наиболее крупный отдел по сравнению с остальными. В нем особенно развиты полосатые тела. К ним переходят функции высшего интегративного центра. На поверхности крыши впервые появляются островки коры очень примитивного строения, ее называют древней — archicortex. Средний мозг теряет значение ведущего отдела, и относительные размеры его сокращаются. Мозжечок сильно развит благодаря сложности и многообразию движений пресмыкающихся. Головной мозг такого типа, в котором ведущий отдел представлен полосатыми телами переднего мозга, называют зауропсидным.
У млекопитающих — маммалийный тип мозга. Для него характерно сильное развитие переднего мозга за счет коры, которая развивается на основе небольшого островка коры пресмыкающихся и становится интегрирующим центром мозга. В ней располагаются высшие центры зрительного, слухового, осязательного, двигательного анализаторов, а также центры высшей нервной деятельности. Кора имеет очень сложное строение и называется новой корой — neocortex. В ней располагаются не только тела нейронов, но и ассоциативные волокна, соединяющие разные ее участки. Характерным является также наличие комиссуры между обоими полушариями, в которой располагаются волокна, связывающие их воедино. Промежуточный мозг, как и у других классов, включает гипоталамус, гипофиз и эпифиз. В среднем мозге располагается четверохолмие в виде четырех бугров. Два передних связаны со зрительным анализатором, два задних—со слуховым. Очень хорошо развит мозжечок (рис. 14.39).
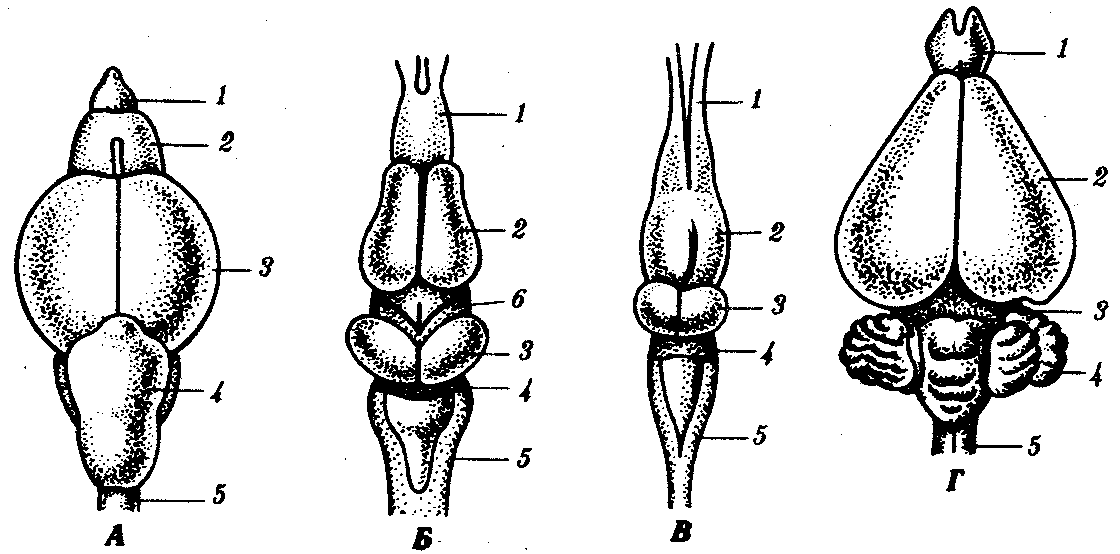
Рис. 14.39. Эволюция головного мозга позвоночных. А—рыба; Б—земноводное; В—пресмыкающееся; Г—млекопитающее:
1—обонятельные доли, 2—передний мозг, 3—средний мозг, 4—мозжечок, 5—продолговатый мозг, 6—промежуточный мозг
По мере усиления функций передних отделов головного мозга в филогенезе спинного мозга наблюдается его продольная дифференцировка с образованием утолщений в области отхождения крупных нервов к конечностям и редукция его заднего конца.
Так, у рыб спинной мозг равномерно тянется вдоль всего тела. Начиная от земноводных происходит его укорочение сзади. У млекопитающих на заднем конце спинного мозга остается рудимент в виде конечной нити — filum terminale. Нервы, идущие к заднему концу тела, проходят по позвоночному каналу самостоятельно, образуя так называемый конский хвост — cauda equina.
Основные этапы эволюции центральной нервной системы отражаются и в онтогенезе человека. На стадии нейруляции закладывается нервная пластинка, превращающаяся в желобок и затем в трубку. Передний конец трубки образует сначала три мозговых пузыря (рис. 14.40): передний (I), средний (II) и задний (III). Вслед за этим передний пузырь подразделяется на два, дифференцирующихся на передний (7) и промежуточный (2) мозг — telencephalon, diencephalon. Средний мозговой пузырь развивается в средний (3) мозг — mesencephalon, a задний — в задний (4) мозг — metencephalon — и продолговатый (5) мозг — medulla oblongata.
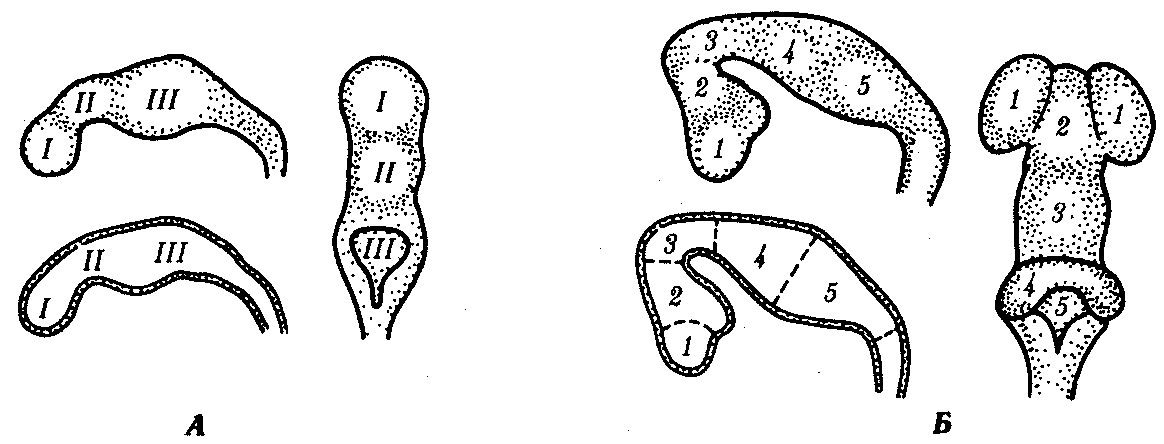
Рис. 14.40. Мозговые пузыри в эмбриогенезе мозга человека. А — стадия трех мозговых пузырей; Б — стадия пяти мозговых пузырей (остальные пояснения см. в тексте)
Задний конец спинного мозга редуцируется, превращаясь в терминальную нить. Позже скорости роста спинного мозга и позвоночника оказываются разными, и к моменту рождения конец спинного мозга оказывается на уровне третьего, а у взрослого человека — уже на уровне первого поясничного позвонка.
Головной мозг рано начинает развиваться по пути, характерному для млекопитающих и человека. Первично почти прямая нервная трубка резко изгибается в области будущего продолговатого и среднего мозга. На этом фоне большие полушария переднего мозга растут с особенно большой скоростью. В результате головной мозг оказывается расположенным над лицевым черепом. Дифференцировка коры приводит к развитию извилин, борозд и формированию высших сенсорных и двигательных центров, в том числе центров письменной и устной речи и др., характерных только для человека (см. § 15.3).
Центральная нервная система столь важна для интеграции индивидуального развития человека, что большинство ее врожденных пороков несовместимы с жизнью. Среди пороков спинного мозга, онтогенетические механизмы которых известны, отметим рахисхиз,нли платиневрию,— отсутствие замыкания нервной трубки (рис. 14.41). Эта аномалия связана с нарушением клеточных перемещений и адгезии в зоне формирования нервной трубки в процессе нейруляции. Аномалия переднего мозга — прозэнцефалия — выражается в нарушении морфо-генеза мозга, при котором полушария оказываются неразделенными, а кора — недоразвита. Этот порок формируется на 4-й неделе эмбриогенеза, в момент закладки переднего мозга. Как и предыдущий, он несовместим с жизнью. Часто встречается у мертворожденных при различных хромосомных и генных синдромах.
Нарушения дифференцировки коры — агирия (отсутствие извилин) и олигогирия с пахигирией (малое количество утолщенных извилин) — сопровождаются упрощением гистологического строения коры. У детей с такими пороками выявляются грубая олигофрения и нарушение многих рефлексов. Большинство детей умирают в течение первого года жизни.

Рис. 14.41. Рахисхиз (платиневрия)
14.6.2. Эндокринная система
Эндокринная система наряду с нервной является ведущим аппаратом интеграции многоклеточного организма, обеспечивая гуморальную регуляцию функций органов. Эта регуляция осуществляется гормонами — биологически активными веществами разной химической природы, выделяемыми железами внутренней секреции. Действие гормонов строго специфично: разные гормоны действуют на разные органы, вызывая определенные изменения их функционирования. Железы внутренней секреции не имеют протоков и выделяют гормоны непосредственно в кровь, что облегчает их транспорт к органам-мишеням. Клетки органов-мишеней на своих мембранах имеют специфические рецепторы, с которыми связываются гормоны, вызывая определенные изменения их метаболизма. Гуморальная регуляция эволюционно возникла значительно раньше нервной потому, что она более проста и не требует развития таких сложных структур, как нервная система.
Обсуждая вопрос о филогенезе эндокринной системы хордовых, необходимо рассмотреть происхождение и эволюцию как гормонов, так и самих желез внутренней секреции.
14.6.2.1. Гормоны
Химическая структура гормонов многообразна. Это могут быть белки (инсулин, пептидные нейрогормоны), стероиды (половые гормоны), продукты метаболизма отдельных аминокислот (тироксин, адреналин). Они могут иметь и другое строение. Это свидетельствует о том, что гормоны разнообразны и по происхождению. Главное, что их объединяет, — способность специфически изменять клеточный метаболизм при контакте с цитоплазматической мембраной.
Предполагают, что уже древние одноклеточные организмы использовали биологически активные вещества для межклеточных коммуникаций. Некоторые вещества такого рода, обладающие регуляторными функциями, могли действовать как на отдельные клетки простейших, так позже и на клетки многоклеточных организмов. Впоследствии они и стали выполнять функции гормонов. Интересно, что в прогрессивной эволюции гуморальной регуляции структура самих гормонов может и не меняться. Доказательством этого является обнаружение таких известных гормонов, как адреналин, норадреналин и некоторые другие, в клетках простейших и низших растений, где они выполняют функции регуляторов клеточного деления, движения ресничек и вакуолей.
В эмбриогенезе многоклеточных ряд гормонов выявляется уже в первые часы и дни развития. В процессе дробления они регулируют течение клеточного цикла. Позже — перемещения клеток и образование межклеточных контактов, действуя либо внутри клеток, их продуцирующих, либо на близлежащие клетки. Гормоны приобретают свойства дистантных регуляторов в филогенезе только у трехслойных животных, а в онтогенезе многоклеточных — соответственно на стадии первичного органогенеза.
При неизменности химической структуры функции гормонов могут нередко изменяться. Так, гормон пролактин, выделяющийся у млекопитающих и человека гипофизом и регулирующий секрецию молока млечными железами, обнаружен также у рыб, земноводных и птиц. У первых он регулирует выделение кожными железами слизи, которой питаются мальки, у вторых — образование оболочек икринок в яйцеводах, у третьих — некоторые элементы брачного поведения, а также выделение у кормящих родителей «зобного молочка». Как видно из этого примера, эволюция действия гормонов может быть канализированной выполнением одной и той же функции в самом широком смысле. Действительно, все перечисленные функции пролактина у животных, находящихся на разных уровнях организации, имеют отношение к обеспечению успешности размножения.
Функции других гормонов, например адреналина, могут в филогенетическом ряду тех же позвоночных практически не меняться, обеспечивая у всех регуляцию в первую очередь энергетического обмена.
При этом часто гормоны продолжают выполнять те функции, которые первично проявлялись еще у одноклеточных и низших многоклеточных животных. Так, адреналин в организме млекопитающих и человека замедляет прохождение клеток по клеточному циклу и вступление их в митоз. Из главной эта функция у высокоорганизованных животных превратилась во второстепенную. Таким образом, на филогенез гормонов, так же как и на эволюцию органов, распространяются основные закономерности макроэволюции (см. гл. 13).
14.6.2.2. Железы внутренней секреции
Железы внутренней секреции, как и гормоны, выделяемые ими, имеют разное происхождение, что важно для изучения их эволюции.
Некоторые эндокринные железы связаны по происхождению с эпителиальной выстилкой глотки. К ним относятся щитовидная и паращитовидная железы. Эпифиз развивается как вырост мозга; гипофиз, надпочечники и поджелудочная железа имеют сложное происхождение.
Среди хордовых только у бесчерепных эндокринная система существует в виде отдельных клеток и клеточных комплексов, которые находятся в разных отделах тела, объединенных друг с другом за счет гуморального взаимодействия. У позвоночных в основании промежуточного мозга развивается гипоталамус — нейросекреторное образование, осуществляющее связь между двумя системами интеграции организма в единое целое: нервной и эндокринной. Вместе с гипофизом гипоталамус образует единую гипоталамо-гипофизарную систему.
Эволюционным предшественником гипоталамуса является так называемый инфундибулярный вырост ланцетника, состоящий из нейросекреторных клеток и находящийся на вентральной стороне переднего конца нервной трубки.
Начиная с рыб гипоталамус дифференцируется на многочисленные ядра, клетки которых с помощью отростков контактируют как с нейронами мозга, так и с клетками гипофиза. Нейросекреторные клетки гипоталамуса выделяют две основные группы гормонов: пептидные и моноаминовые.
Первые — гормоны, влияющие на функции внутренних органов — вазопрессин, регулирующий артериальное давление, окситоцин, действующий на мускулатуру матки, и др. Вторая группа гормонов (дофамин, норадреналин, серотонин) регулирует деятельность передней доли гипофиза. Под их действием стимулируется или подавляется секреция гормонов соответствующими гипофизарными клетками.
Подробнее остановимся на эволюции гипофиза. Эта железа состоит из трех долей: передней (аденогипофиза), средней (промежуточной) и задней (нейрогипофиза). Гипофиз соединен с гипоталамусом его выростом — воронкой, через которую проходят отростки нейронов гипоталамуса и кровеносные сосуды, обеспечивающие их гуморальное взаимодействие.
Доли гипофиза имеют разное происхождение. Передняя доля развивается из выпячивания эктодермального эпителия крыши ротовой полости, так называемого кармана Ратке, который растет в сторону промежуточного мозга. Задняя доля развивается из задней части воронки. Клетки, входящие в ее состав, по происхождению являются глиальными. Промежуточная доля — производная от передней (рис. 14.42).
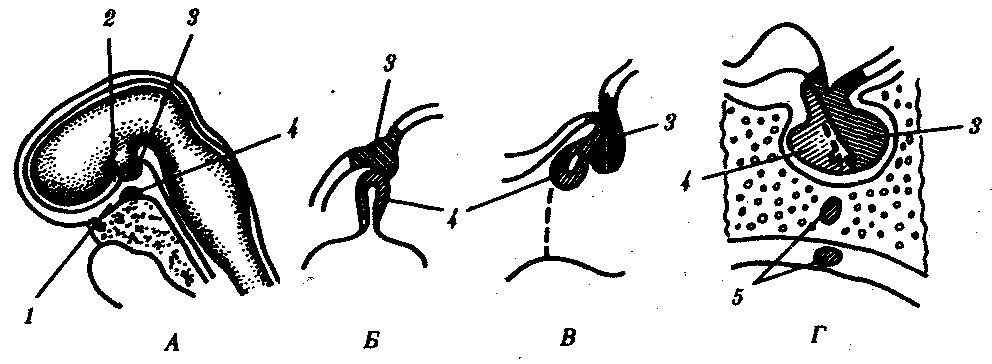
Рис. 14.42. Развитие гипофиза у человека. А—сагиттальный срез 4-недельного зародыша; Б—объединение зачатков гипофиза из стенки глотки и основания мозга; В—формирование трех долей гипофиза на 8-й неделе развития; Г—сформированный гипофиз:
1—крыша ротовой полости, 2—основание мозга, 3—выпячивание основания мозга (задняя доля гипофиза), 4—карман Ратке (промежуточная и передняя доли гипофиза), 5—аномалия развития (добавочные дольки передней доли в стенке глотки и внутри клиновидной кости, оставшиеся в области выпячивания кармана Ратке)
У хрящевых рыб во взрослом состоянии сохраняется первоначальная связь передней доли гипофиза с эпителием ротовой полости. За счет ее задней части формируется также средняя доля. Обе доли вырабатывают гонадотропные гормоны. У костных рыб и личинок земноводных имеются передняя и промежуточная доли, а у взрослых амфибий, переходящих к наземному существованию, появляется также задняя, регулирующая водный обмен. Средняя доля у них перестает выделять гонадотропный гормон, но вырабатывает пролактин.
В связи с наземным образом жизни пресмыкающихся и млекопитающих у них наиболее прогрессивно развивается задняя доля гипофиза, что связано с интенсификацией водного обмена.
Передняя доля вырабатывает соматотропный гормон (гормон роста) и ряд гормонов, регулирующих функции других желез внутренней секреции, а средняя — пролактин и некоторые другие. При этом усиливается дифференцировка и интенсифицируются функции ядер гипоталамуса, находящихся в функциональной связи со всеми долями гипофиза.
У человека в эмбриогенезе развитие гипофиза соответствует основным этапам его эволюции. Очень часто, в 30—40%, у нормальных людей под слизистой оболочкой крыши глотки, в основании клиновидной кости, обнаруживается группа клеток длиной 5—6 мм и шириной 0,5—1 мм, по структуре и функциям соответствующая передней доле гипофиза. Это результат нарушения перемещения клеток при закладке гипофиза в эктодерме ротовой полости в области турецкого седла. Эту 139 аномалию называют эктопией аденогипофиза, она не сопровождается патологическими проявлениями. Более опасно сохранение полости в области кармана Ратке — киста кармана Ратке. Она располагается между передней и промежуточной долями, содержит слизь и в ряде случаев имеет тенденцию к росту и даже к переходу в злокачественное новообразование. Растущие кисты сдавливают гипофиз и вызывают его гипофункцию. Больные нуждаются в оперативном вмешательстве.
Щитовидная железа, гормон которой — тироксин — регулирует энергетический обмен, среди хордовых как компактный орган впервые появляется у рыб. Однако уже у ланцетника отдельные тироксин-синтезирующие клетки обнаруживаются в желобке на вентральной стороне глотки. Щитовидная железа рыб закладывается также в виде желобка на вентральной стороне глотки между 1-й и 2-й жаберными щелями в области зачатка основания языка. Позже этот клеточный материал погружается под слизистую оболочку и формирует фолликулы, характерные для щитовидной железы. У других позвоночных железа закладывается так же, как у рыб, но затем она перемещается в область подъязычной кости (у земноводных) или в шейную область (у пресмыкающихся и млекопитающих).
У человека в эмбриогенезе щитовидной железы происходит рекапитуляция предковых состояний. Гетеротопия ее осуществляется посредством миграции клеток в виде тяжа, полого внутри, называющегося щитоязычным протоком — canalis thyreoglossus. При нормальном развитии этот проток полностью редуцируется, рудиментом его является слепое отверстие foramen caecum в корне языка.
Свидетельством эмбрионального перемещения железы является и расположение верхней щитовидной артерии a. thyreoidea superior, которая, начинаясь от общей сонной артерии, резко поворачивает вниз и входит в ее ткань. На ранних этапах эмбрионального развития эта артерия направляется вверх к зачатку щитовидной железы, а затем меняет свое направление вместе с его перемещением книзу.
Персистирование участков подъязычного протока в постнатальном периоде сопровождается накоплением в них жидкости и образованием срединных кист шеи, которые могут располагаться в любом месте от корня языка до верхней границы щитовидного хряща. Иногда, кисты загнаиваются и прорываются с образованием срединных свищей шеи. Не менее известны такие пороки развития, как эктопия щитовидной железы, которая может развиться в результате нарушения клеточной миграции (рис. 14.43).
Из эпителия глотки в области III-V жаберных карманов у позвоночных развиваются мелкие эндокринные образования, связанные по месту окончательного положения с щитовидной железой. Это паращитовидные железы и ультимобранхиальные тельца. Первые, выделяя гормон паратиреоидин, повышающий содержание ионов кальция в крови и уменьшающий их количество в костях, развиваются как самостоятельные железы только у наземных позвоночных, а у земноводных — лишь после метаморфоза. Клетки вторых выделяют кальцитонин, являющийся антагонистом паратиреоидина. Таким образом, паращитовидные железы и ультимобранхиальные тельца являются регуляторами кальциевого обмена. Значение их наиболее велико у наземных позвоночных. В филогенетическом ряду позвоночных они постепенно перемещаются из глоточной области в сторону щитовидной железы, а у млекопитающих даже срастаются с ней. Паращитовидные железы еще сохраняют самостоятельность, а клетки ультимобранхиальных телец мигрируют между фолликулами щитовидной железы и сохраняются под названием парафолликулярных клеток. Таким образом, на примере щитовидной, паращитовидных желез и ультимобранхиальных телец видна интеграция железистых структур в сложное надерганное образование, выполняющее целый комплекс взаимосвязанных функций.

Рис. 14.43. Аномалии щитовидной железы:
1—нормальное расположение. железы, 2—место эмбриональной закладки железы, 3—варианты аномального расположения железы; стрелками показано направление перемещения зачатка щитовидной железы в эмбриогенезе
У человека известны аномалии расположения паращитовидных желез, связанные с нарушением их гетеротопий.
Надпочечники позвоночных имеют двойственное происхождение. У рыб и земноводных ткани, соответствующие мозговому и корковому веществам этих желез, расположены отдельно друг от друга. Зачатки мозгового вещества связаны по происхождению с симпатическими нервными узлами и расположены метамерно. Зачатки коркового вещества развиваются из утолщений эпителия брюшины. У наземных позвоночных мозговое и корковое вещества объединяются в компактные эндокринные железы, имеющие сложное гистологическое строение. У млекопитающих они прилежат к переднему концу почек. Мозговое вещество выделяет в основном адреналин — регулятор кровообращения и энергетического обмена, а корковое вещество — разнообразные стероидные гормоны, влияющие на минеральный, углеводный обмены и функции почек.