
Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году в сборниках по проблемам кибернетики. Тогда же вышла статья
| Вид материала | Статья |
- Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году, 4857.78kb.
- I вступление, 32.31kb.
- Культура Кольского Севера; Творческие работы посвященные проблемам студенческой молодежи, 67.08kb.
- «Мой учитель» посвященные к Году учителя, 64.26kb.
- Михаил Александрович Шолохов (родился. В 1905 году) Когда в 1925 году появились Донские, 334.1kb.
- Лекция 21. Генетика пола Определение пола, 33.63kb.
- Публичный доклад о результатах деятельности школы моу сош п. Пола в 2008-2009 учебном, 335.49kb.
- Курс «Обзор перспективных технологий Microsoft. Net» Губанов Ю. А., математико-механический, 177.56kb.
- Больше-Кочинский школьный музей этнографии и фольклора, 25.15kb.
- Воспитание и обучение одаренных детей, 267.86kb.
Взаимоотношения системы со средой
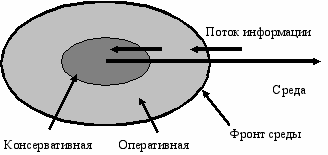
Дифференциация системы на постоянную и оперативную память создает структуру “стабильного ядра” и “лабильной оболочки” в информационных взаимоотношениях системы со средой (Рис. Б.3). Если рассматривать потоки, передающие генетическую информацию от поколения к поколению, то емкости постоянной памяти образуют осевую (генетическую) линию, тогда как емкости оперативной памяти составляют боковую (экологическую) линию, “вынос” части информации навстречу факторам среды.
Р
 ис. Б.3
ис. Б.3По оси “система-среда” система
разделяется на “стабильное ядро”
и “лабильную оболочку”
Центростремительные потоки информации, поступающие от среды, попадают сначала в оперативную память системы, преобразуются там, подвергаются отбору, и только после этого часть информации попадает в постоянную память (Рис. Б.3). Такая структура систем предполагает существование каких-то барьеров между подсистемами, препятствующих смешению всей информации. Описанный способ хранения генетической информации, в двух объемах, сообщающихся между собой каналом связи контролируемого сечения, повышает устойчивость системы.
Для обеспечения информационного контакта системы со средой в основном через оперативную память, необходимо, чтобы элементы оперативной памяти обладали большей дисперсией признаков по сравнению с элементами постоянной памяти, то есть первые должны быть разнообразнее вторых.
Осуществление преимущественного контакта среды с оперативной памятью за счет сдвига средних значений признаков, а не за счет разной дисперсии признаков не решает задачу в случае альтернативных признаков. Например, одна и та же особь может быть одновременно и жаростойкой и морозостойкой (неальтернативный признак), но не может быть одновременно и крупной и мелкой (альтернативный признак).
Отставание консервативной подсистемы от оперативной
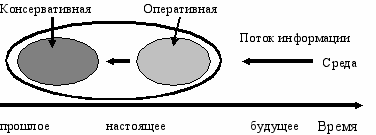
Необходимость переработки новой информации в оперативной памяти, до того как она сможет попасть в постоянную, делает постоянную память системы инерционной. Инерционность постоянной памяти, её отставание от оперативной в получении новой информации от среды придает ей и её элементам черты совершенства. Напротив, элементы оперативной памяти приобретают черты прогрессивности. Рассматривая эволюцию системы во временнóм аспекте, можно сказать, что оперативная подсистема содержит новые признаки, которые в будущем перейдут в консервативную. Поэтому оперативную подсистему можно считать эволюционным “авангардом” системы (Рис. Б.4).
Такое разделение и специализация подсистем по альтернативным задачам сохранения и изменения обеспечивает оптимальные условия для реализации основного метода эволюции живых систем—метода отбора, то есть в известном смысле метода проб и ошибок. С сосредоточением проб в оперативной памяти там же локализуются и ошибки и находки. Это дает возможность системе пробовать различные варианты решения эволюционных задач без риска закрепления неудачных решений.
Р
 ис. Б.4
ис. Б.4По оси времени оперативную
подсистему можно рассматривать
как “авангард” по сравнению с
консервативной.
Примеры бинарно-сопряженных подсистем
ДНК — белки. Рассматривая нуклеопротеид как систему, с подсистемами ДНК и белок, можно видеть, что ДНК более стабильна. О большей лабильности белков по сравнению с ДНК говорят температуры их денатурации, которые у белков значительно ниже, чем у ДНК. Смысл дифференциации—окружить более устойчивые к факторам среды молекулы ДНК менее устойчивыми белками. Например, если температура денатурации ДНК примерно 65оС, а белков—около 45оС, то вирус, представляющий собой ДНК в белковой оболочке, денатурирует при 45оС, то есть благодаря сопряженному с ней белку нечувствительная к 45оС ДНК становится чувствительной. Значит, белок является информационным посредником между ДНК и средой, способным “предупредить” заблаговременно о наступлении жары. Другие белки позволяют ДНК “ощущать” холод, третьи—“видеть”, “слышать” и таким образом “узнавать” об опасностях среды.
Морфологическое строение известных нуклеопротеидов вирусов также говорит о том, что ДНК (или РНК у некоторых вирусов) в них представляет “ядро”, а белки—“оболочку”. Поэтому со средой прежде всего взаимодействуют белки и поток информации должен быть среда ↔ белок ↔ ДНК. Центробежный поток информации, ДНК → белок, хорошо изучен. Это поток информации, осуществляющий синтез белков и определяющий поведение системы. Больший интерес представляет обратный, центростремительный поток информации: среда → белок → ДНК, в особенности звено белок → ДНК. Долгое время вообще отрицалось существование такого потока. Конечно, можно представить, что в процессе эволюции, когда образовались более высокие уровни организации (клетки, организмы) и появились соответствующие обратные связи, то обратные связи на нижележащих уровнях (в данном случае на молекулярном) утратили свое значение и исчезли. Однако существуют факты, которые очень трудно объяснить, не допустив наличия центростремительных потоков белок → ДНК. К ним можно отнести прежде всего явления, связанные с образованием адаптивных ферментов и антител.
Аутосомы—половые хромосомы. Дифференциация на аутосомы и половые хромосомы существует не у всех видов. Кроме того, между хромосомами нет видимых барьеров. Однако в пользу взгляда на аутосомы и половые хромосомы как на постоянную и оперативную память ядра свидетельствуют многие факты. Эволюционно половые хромосомы значительно “моложе” аутосом и произошли из них, а Y-хромосома на 20–80 тыс. лет моложе Х-хромосомы. Отмечена большая лабильность и изменчивость половых хромосом по сравнению с аутосомами. Среди ядер с ненормальным числом хромосом подавляющее большинство составляют отклонения, связанные с половыми хромосомами (у человека известен даже набор ХХХХY (Штерн, 1965)). При действии ультразвука прежде всего разрушаются половые хромосомы. Радиоактивные вещества метят половые хромосомы интенсивнее аутосом (в особенности Y-хромосому). По дисперсии размеров половые хромосомы (прежде всего Y-хромосома) также превосходят аутосомы. Есть указания на преимущественно периферическое расположение половых хромосом в ядре у человека (Barton et al., 1964).
Ядро—цитоплазма. На клеточном уровне организации бросается в глаза прежде всего дифференциация на ядро и цитоплазму. Морфология клеток делает довольно очевидными соотношения между ядром, цитоплазмой и средой, как в смысле последовательности попадания информации (среда → цитоплазма → ядро), так и в смысле большего разнообразия цитоплазм (клетки разных тканей) и однообразия ядер. Показано, что ядра в любой клетке организма (по крайней мере у определенных видов) содержат всю генетическую информацию (постоянная память), в то время как цитоплазма клетки определяет специализацию данного сорта клеток (оперативная память), определяет, какую часть генетической информации необходимо извлечь из ядра в каждом отдельном случае.
Гаметы—соматические клетки. На организменном уровне организации элементами являются клетки. Клетки организма делятся на гаплоидные (гаметы) и диплоидные (соматические). Прежде всего обращает на себя внимание большое разнообразие (морфологическое и физиологическое) соматических клеток по сравнению с однообразием гамет. Очевидна также специализация этих подсистем в осуществлении задач наследственности и изменчивости. Гаметы представляют консервативную тенденцию, а соматические клетки, наоборот, тенденцию изменчивости. Потоки информации от среды в течение всей жизни воспринимают соматические клетки (организмы растут, развиваются, стареют и пр.), и только после переработки в оперативной памяти суммарная за жизнь, итоговая информация может попасть в постоянную память, в гаметы (в форме элиминации, дискриминации или привилегии данной особи или в форме передачи каких-либо мутагенных влияний).
Общество (правши—левши). Дифференциация общества на правшей и левшей создает два поведенческих режима (психики): консервативный режим правшества—психология сохранения (аналог женского пола), и оперативный режим левшества—реформаторская психология изменения, поиска нового, (аналог мужского пола). Первые максимально приспособлены к оптимальной (стабильной) среде, вторые—к экстремальной.
* * *
Сопряженные пары подсистем не исчерпываются только приведенными структурами. Уже сейчас можно привести (или возможно, что будут открыты в будущем) и другие самовоспроизводящиеся системы, построенные аналогичным образом. Между парами бактерия—фаг, ген—признак и другими существуют, видимо, во многих отношениях такие же взаимоотношения.
Дифференциация на сопряженные подсистемы имеет место не только в самовоспроизводящихся структурах, но также во многих биологических рабочих системах и вообще во всех эволюционирующих, следящих системах. Например, такие биологические системы, как кофермент—апофермент, антиген—антитело, правое—левое полушария и подкорка—кора головного мозга, симпатическая—парасимпатическая вегетативная нервная система и другие обладают чертами, характерными для постоянной и оперативной памяти.
Принцип сопряженных подсистем лежит также в основе дифференциации некоторых социальных институтов: государственный аппарат—правительство, производство—наука, больница—поликлиника, школа—вуз. Отмечались, например, аналогичные соотношения между наукой и производством, с одной стороны, и взаимоотношениями подсистем в популяции—с другой (Гуревич, 1966). Такие черты науки, как большее разнообразие направлений, относительная лабильность, право на поиск, с вытекающими отсюда надеждой находок и риском ошибок, придают ей свойства прогрессивности (оперативная память). Напротив, меньшая дисперсия направлений, относительная стабильность и инерционность, требование рентабельности, придают производству черты совершенства (консервативная память). Можно думать, что для всех эволюционирующих систем, тесно связанных со средой, такая дифференциация повышает устойчивость и способствует их дальнейшей эволюции.
Приложение В: Рисунки и Таблицы
Р


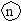
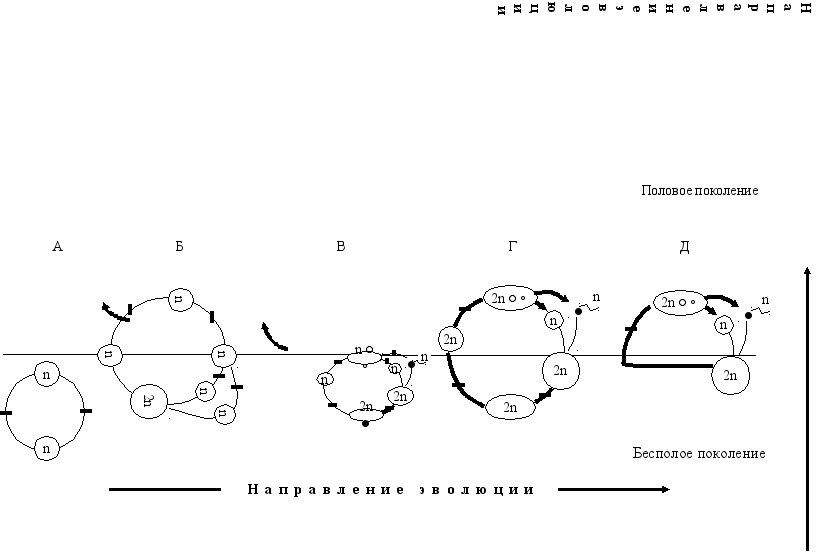
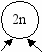 ис. 6.3 Эволюция размножения (Геодакян, Геодакян, 1986). А—только бесполое размножение; Б, В, Г—чередование бесполого и полового поколений;
ис. 6.3 Эволюция размножения (Геодакян, Геодакян, 1986). А—только бесполое размножение; Б, В, Г—чередование бесполого и полового поколений;Д—– только половое размножение. ▬ - 2n—Диплоидная фаза цикла, ─ - n—Гаплоидная фаза цикла, ▬► —Мейоз, ─▌─ —Митоз, ● —Спора,
— одноклеточный или многоклеточный организм, — зигота
● n — гаметы мужские, женские; — процесс оплодотворения
Табл. 2.1 Половой диморфизм по основным причинам смерти (США, 1967 г.)*
| Причина смерти | Смертность на 100 000 | Соотношение полов, % ♂♂ | |
| мужчин | женщин | ||
| Опухоли дыхательной системы | 50.1 | 8.5 | 85.5 |
| Другие болезни дыхательной системы (71% эмфизема) | 24.4 | 5.0 | 83.0 |
| Цирроз печени | 18.5 | 9.1 | 67.0 |
| Атеросклероз, включая болезни сердца | 357.0 | 175.6 | 67.0 |
| Симптомы старости и болезней | 14.9 | 8.3 | 64.2 |
| Пневмония исключая новорожденных | 32.3 | 19.5 | 62.4 |
| Другие болезни сердца | 17.9 | 11.1 | 61.7 |
| Другие болезни сосудистой системы | 18.2 | 11.1 | 62.1 |
| Опухоли органов пищеварения и брюшины | 53.0 | 36.2 | 59.4 |
| Все остальные болезни | 32.4 | 22.4 | 59.1 |
| Опухоли других и неопределенных мест | 20.5 | 14.7 | 58.2 |
| Родовые повреждения, постнатальная асфиксия и ателектаз | 11.9 | 8.4 | 58.6 |
| Некоторые болезни раннего детства | 29.2 | 21.6 | 57.5 |
| Другие болезни, особенно раннего детства и юношества | 15.3 | 11.7 | 56.7 |
| Неревматический хронический эндокардит и повреждения миокарда | 26.8 | 20.5 | 56.7 |
| Общий атеросклероз | 17.2 | 14.8 | 53.8 |
| Повреждения сосудов, влияющие на центральную нервную систему | 96.3 | 83.3 | 53.6 |
| Гипертоническая болезнь | 22.3 | 22.2 | 50.1 |
| Опухоли гениталий | 17.9 | 20.1 | 47.1 |
| Сахарный диабет | 14.9 | 16.8 | 47.0 |
| Опухоли груди | 0.2 | 24.6 | 0.8 |
| Все случаи: | 891.2 | 565.5 | 61.2 |
*—Включены все причины смерти, дающие вклад более 1%.
Табл. 4.3 Соотношение полов при злокачественных новообразованиях и врожденных пороках развития
(сводные данные мировой статистики) (из Раевский, Шерман, 1976 с изменениями).
| Системы и органы | Филогенетическая “новизна” органа и системы | Локализация органа в пределах системы | Соотношение полов, ♂♂ : ♀♀ | Врожденный порок | |
| По раку | По порокам** | ||||
| | | | | | |
| Нервная система | Относительно древняя | | | | |
| Головной мозг | | | 1.21* 1.28*** | 0.54 0.57 | Анэнцефалия Черепно-мозговые грыжи |
| Спинной мозг | Новый | | 1.27* | 0.73 | Спинно-мозговые грыжи |
| Периферические нервы | Древние | | 1.04* | | |
| | | | | | |
| Система дыхания | Новая | | 0.68* | | |
| Гортань | Новый | Вход системы | 11.66* 9.1*** | | |
| Легкие | Новый | Внутреннее звено | 6.09* 6.66*** | 0.73 | Агенезия Аплазия Гипоплазия легкого |
| в том числе: | | | | | |
| центральный рак | | | 11.05* | | |
| периферический рак | | | 7.7* | | |
| бронхиолярный рак | | | 0.75* | | |
| | | | | | |
| Пищеварительная система | Древняя | | 1.3* | 1.5 | Все органы пищеварительной системы |
| Язык | Относительно новые | Вход системы | 3.09* 2.3*** | | |
| | | | | | |
| | | | | Продолжение на следующей странице | |