
Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году в сборниках по проблемам кибернетики. Тогда же вышла статья
| Вид материала | Статья |
СодержаниеДисперсия полов Повышенный уровень мутаций у мужских особей Наследование родительских признаков Более широкая норма реакции женского пола |
- Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году, 4857.78kb.
- I вступление, 32.31kb.
- Культура Кольского Севера; Творческие работы посвященные проблемам студенческой молодежи, 67.08kb.
- «Мой учитель» посвященные к Году учителя, 64.26kb.
- Михаил Александрович Шолохов (родился. В 1905 году) Когда в 1925 году появились Донские, 334.1kb.
- Лекция 21. Генетика пола Определение пола, 33.63kb.
- Публичный доклад о результатах деятельности школы моу сош п. Пола в 2008-2009 учебном, 335.49kb.
- Курс «Обзор перспективных технологий Microsoft. Net» Губанов Ю. А., математико-механический, 177.56kb.
- Больше-Кочинский школьный музей этнографии и фольклора, 25.15kb.
- Воспитание и обучение одаренных детей, 267.86kb.
Дисперсия полов
Чтобы популяция “чувствовала” приближение фронта вредного фактора среды, необходим контакт кривой смертности популяции с этим фронтом. То есть за получаемую информацию популяции приходится все время платить определенную жертву в виде элиминации самых чувствительных к данному фактору особей. При этом плата пропорциональна получаемой информации и тесно связана с фенотипической дисперсией популяции. Если фенотипическая дисперсия мала, то в стабильной среде нет элиминации и нет информационного контакта популяции со средой. В этом случае очередное внезапное изменение среды может застать популяцию врасплох и погубить ее целиком. Наоборот, если фенотипическая дисперсия очень велика, то чрезмерно растет плата за новую информацию, а это расточительно. Следовательно, существует некая оптимальная для данной популяции в данной среде величина фенотипической дисперсии, которая обеспечивает заблаговременное получение необходимой информации при минимальной плате за нее.
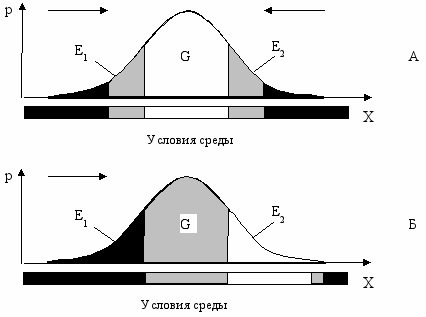
Если схематично разделить популяцию на три части, как показано на Рис. 7.2А, то в привычных старых условиях среды, центральная часть популяции пребывает в комфортных условиях по данному фактору среды, а периферийные части—в условиях дискомфорта. При этом периферийные части E1 и E2 страдают от диаметрально противоположных (альтернативных) факторов, например, если часть E1 страдает от холода, то часть E2—от жары. Это означает, что даже в популяциях животных, обитающих в тропиках, обезьян к примеру, какие-то экземпляры погибают не только от жары, но и от холода, тогда как в популяциях пингвинов или белых медведей, какие-то особи погибают от тепла.
Р
ис. 7.2
Взаимоотношение популяции со средой в условиях стабилизирующего (А) и движущего отбора (Б).
Зоны □—комфорта; ░—дискомфорта; █—элиминации. →—направление давления отбора;
X—обобщенный признак (устойчивость к фактору среды), p—вероятность (или концентрация) его
в популяции, G—генетическая часть популяции, E1, E2—экологические части.
При изменении условий среды, часть E1 попадает в зону элиминации, часть G—в зону дискомфорта, а часть E2— в зону комфорта (Рис. 7.2Б). Особи класса E2, которые в обычных условиях страдали от дискомфорта, попав наконец, в комфортные условия, бурно размножаются и “уводят” популяцию от наступающего вредного фактора среды.
Информативность особей на концах кривой генотипического распределения (области E1 и E2) максимальна, так как именно они могут вызвать максимальный сдвиг генотипов в следующем поколении (Рис. 7.3). Чем дальше генотип от моды, чем он оригинальнее, тем выше его информативность. Однако самых оригинальных особей мало и их вклад в генофонд потомства ничтожен. Поэтому реальный вклад будет определяться произведением степени оригинальности на концентрацию особей с данным значением признака, а также на сечение их канала связи с потомством. Поскольку сечение канала связи самцов с потомством значительно больше чем самок, их вклад также будет выше.
То есть, информационная ценность особей разных классов разная. Центральный класс, максимально приспособленный к старым условиям и поэтому ориентированный как бы на прошлое, “работает” больше на сохранение имеющейся генетической информации, реализует больше наследственную тенденцию—сохранения. Периферийные классы E1 и E2 оба плохо приспособленные к старым условиям среды, новые условия встречают по разному: особи класса E1 гибнут, а класса E2 становятся самыми приспособленными. Поэтому можно сказать, что классы E1 и E2 “работают” больше на изменение, реализуют больше экологическую тенденцию, связь со средой и ориентированы больше на будущее. Иными словами, в популяции существует специализация, своего рода “разделение труда” по связи со средой (изменчивость) и по связи с предшествующими поколениями (наследственность). Одинаково ли представлены мужской и женский пол в этих специализированных классах популяции (E и G) или соотношение полов в них отклоняется от пропорции 1 : 1?
Р
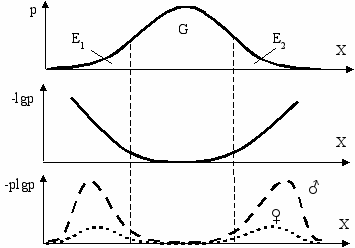
ис. 7.3 Распределение фенотипов (p), их информативность (-lgp) и
информационная эффективность (-plgp) в популяции (Геодакян, 1983б).
X—обобщенный признак (или фенотип), p—вероятность его нахождения в популяции,
G—генетическая часть популяции, E1, E2—экологические части.
Из верхней кривой видно, что в популяции больше всего особей со средним значением признака. Кривая в центре показывает, что у самых редких вариантов информативность максимальна, а у нормы популяции—минимальна. Результирующая (нижняя) кривая учитывает как информативность фенотипов, так и их концентрацию.
Уже отмечалось, что от всех вредных факторов среды, как правило, прежде всего гибнут мужские особи. Мужской пол является биологически более слабым полом. Это значит, что если нарисовать отдельно кривые смертности для каждого пола, то с фронтом вредного фактора среды должна контактировать в основном кривая мужского пола. Следовательно, либо должен иметь место определенный сдвиг кривых таким образом, чтобы мужская кривая оказалась между фронтом вредного фактора и женской кривой, либо это может быть достигнуто за счет большей дисперсии мужской кривой по сравнению с женской. Первое решение не годится, поскольку сдвиг кривых по средним значениям признака, спасая женский пол от данного вредного фактора среды (скажем, холода), подставляет его под удар альтернативного фактора (жары). Остается второе решение—кривая фенотипического распределения мужского пола должна иметь бóльшую дисперсию, чем женского (Рис. 7.4). Это одно из основных положений новой теории.
мужской пол ДОЛЖЕН иметь бóльшую фенотипическую
дисперсию признаков по сравнению с женским
Идея, что мужчины более часто отклоняются в хорошую или плохую сторону (больше гениев и больше идиотов), была введена в начале 20-го века чтобы объяснить, почему мужчины превосходят женщин во многих областях деятельности несмотря на то, что по средним значениям психологических тестов различие между полами не очень велико.
Shields, 1982
Ещё со времен Дарвина биологи отмечали, что по многим признакам у многих видов самцы более изменчивы (гипотеза Большей изменчивости мужского пола) (Edge, 2005).Р
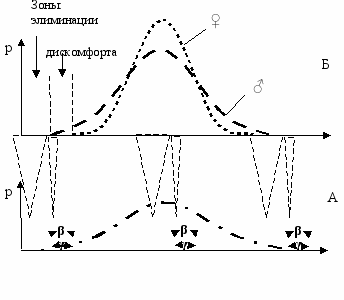
ис. 7.4 Распределение мужских (- - -) и женских (. . .) генотипов (А) и фенотипов (Б) в популяции.
β и γ—нормы реакции мужского и женского пола соответственно (Геодакян, 1983б).
Широкая норма реакции позволяет женским особям уходить из зон элиминации и дискомфорта.
Чтобы первым получать информацию от среды, мужской пол должен иметь повышенную чувствительность (смертность) ко всем факторам среды. Это достигается за счет большей дисперсии признаков, то есть по всем признакам мужской пол должен быть разнообразнее женского. Более широкая фенотипическая дисперсия позволяет за новую информацию “платить” в первую очередь жертвой мужских особей. В то же время мужская особь может оставить несравненно большее количество потомков по сравнению с женской (передача экологической информации потомству). Значит экологическую информацию о происшедших в среде изменениях мужской пол получает и передает потомству эффективнее, чем женский. С другой стороны картину генотипического распределения в популяции полнее передает потомству женский пол.
Следовательно, генетический поток информации (от поколения к поколению) больше реализует женский пол, а экологический (от среды к потомству)—мужской. Более широкая фенотипическая дисперсия мужского пола может быть следствием широкой генотипической дисперсии. Она может быть также следствием более широкой наследственной нормы реакции женских особей, которая позволяет им покинуть зоны элиминации и дискомфорта. Более широкая генотипическая дисперсия мужского пола может быть результатом более высокого уровня мутаций у мужских особей, а также того, что женские потомки наследуют родительские признаки более аддитивно (промежуточное, средне-арифметическое наследование), чем мужские.
Повышенный уровень мутаций у мужских особей
Поскольку число клеточных делений в сперматогенезе намного больше числа делений в овогенезе и ошибки при репликации и репарации ДНК являются главным источником мутаций, был сделан вывод, что все это может привести к большей частоте мутаций в половых хромосомах по сравнению с аутосомами и было предположено, что самцы служат генератором мутаций, по крайней мере, в эволюции млекопитающих. Тот факт, что уровень, как спонтанного так и индуцированного мутагенеза, у гетерогаметных, и у гомогаметных самцов выше чем у самок для дрозофилы, шелкопряда, млекопитающих, в том числе и человека был установлен давно и неоднократно (Анисимов, Соловьев, 1999; Kerkis, 1975). Более высокий уровень точковых мутаций отмечен также у самцов мышей по сравнению с самками (Searle, 1972).
Сравнительным исследованием замещений нуклеотидной последовательности А-, Х- и Y-генов человека и мыши (или крысы) было показано, что самцы являются главным источником мутаций для молекулярной эволюции. Кроме того, было установлено, что соотношение скоростей эволюции генов— Y : А : Х = 2.2 : 1 : 0.6 (хорошо согласуется с теоретическим ожиданием— 2 : 1 : 2/3) (Miyata et al., 1987). В другой работе аналогичной методикой сравнивали Y/X отношения скоростей замещения нуклеотидных последовательностей в генах человека, орангутана, бабуина и беличьей обезьяны. Было показано, что Y-гены дивергируют быстрее и “дальше” друг от друга, чем Х-гены, то есть и у высших приматов самцы предваряют молекулярную эволюцию (Shimmin et al., 1993).
Наследование родительских признаков
Бадр и Спикет (Badr, Spickett, 1965) обнаружили различия между самцами и самками мышей в относительной роли аддитивных и неаддитивных наследственных факторов в определении веса надпочечников. Доля аддитивного генетического разнообразия в фенотипической изменчивости веса надпочечников составляла 40% для самцов и 60% для самок.
По данным Л. Шюлера и др. (1976) у самок мышей гибридов значения относительного веса надпочечников, тимуса, половых желез и гипофиза практически совпадали с полусуммой значений этого признака у родителей. У самцов гибридов наблюдалось отклонение среднего в направлении меньшего значения для надпочечников, тимуса и половых желез, и большего—для гипофиза. У самок мышей гибридов было отмечено также более аддитивное наследование генов, ответственных за двигательную активность (Бородин и др. 1976).
Более широкая норма реакции женского пола
Мужчина женится на женщине и надеется, что она не изменится.
Женщина выходит замуж и надеется, что ее муж изменится.
И они оба неправы.
Где Марья гнется, Иван ломается.
Русская пословица
Разная фенотипическая дисперсия полов может быть связана с их разной нормой реакции. Норма реакции, или модификационная изменчивость, особей женского пола должна быть больше соответствующей нормы реакции особей мужского пола. Иными словами, связь фенотипа с генотипом у мужского пола должна быть более “жесткая”, чем у женского. Поэтому влияние среды в онтогенезе при реализации генотипа в фенотип больше у женского пола (Рис. 7.2, Рис. 7.5).
В “пространстве возможностей” существует некая зона “онтогенетического дискомфорта”, находясь в которой особи не гибнут, но испытывают трудности и неудобства, страдают от неприспособленности (мерзнут, голодают). Широкая норма реакции позволяет особям женского пола покинуть эту зону и выжить, в то время как мужские особи остаются в ней и гибнут. У женских особей за счет широкой нормы реакции онтогенетическое “пространство способностей” больше, чем у мужских. Поэтому половую дифференциацию можно трактовать еще как специализацию по взаимно дополнительным качествам совершенства (женский пол) и новаторства, прогрессивности (мужской) во взаимоотношениях со средой, так как адаптация женских особей к существующим условиям среды совершеннее, чем у мужских особей. А у последних наличие разнообразных отклонений от нормы (моды) популяции (несовершенств) обеспечивает им большую прогрессивность в приспособлении к изменившимся условиям среды в будущем.
Р
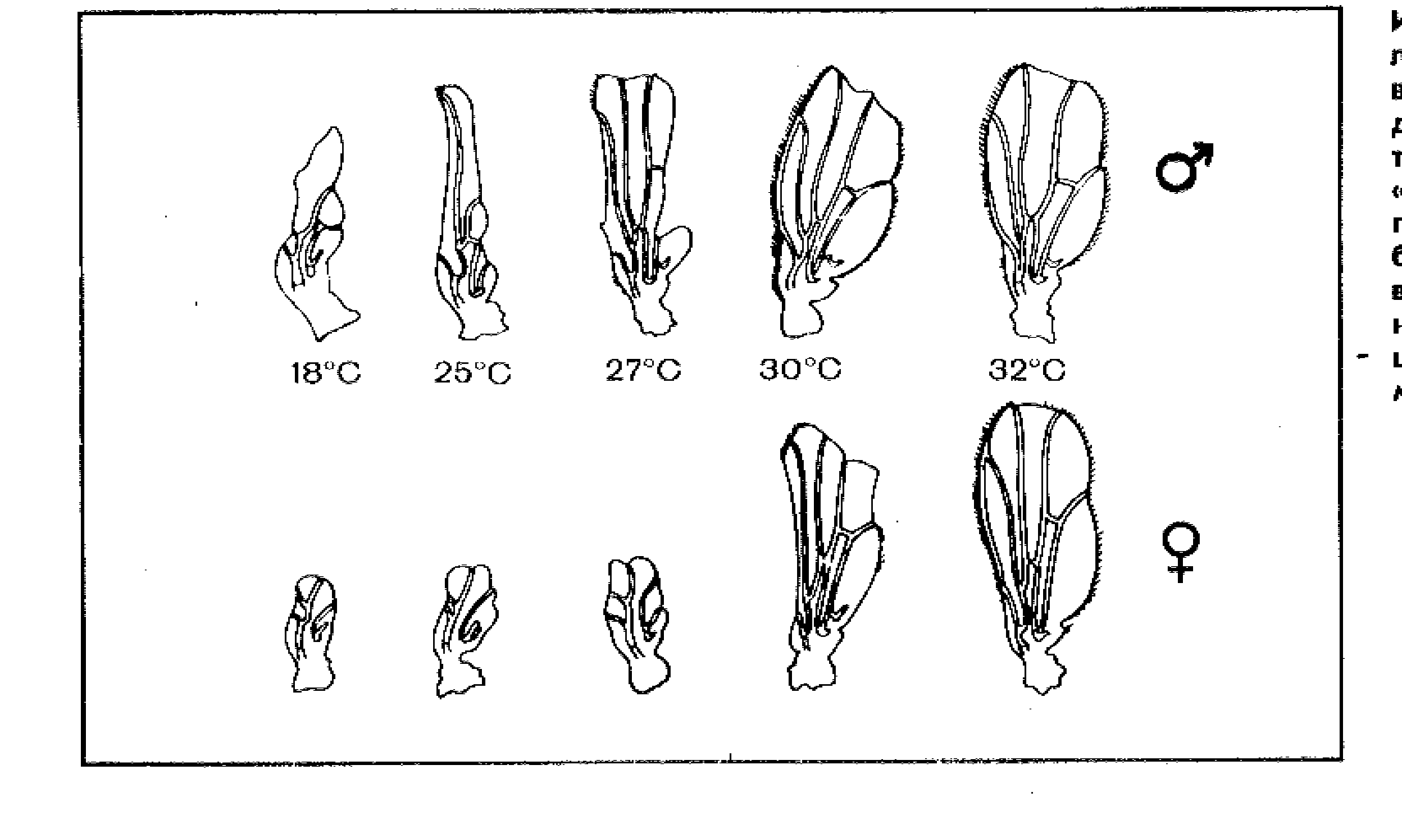
ис. 7.5 Изменение размера и формы крыла у
самцов и самок дрозофилы в зависимости от
температуры среды (Шмальгаузен, 1969).
В онтогенезе из-за более широкой нормы реакции влияние среды на проявление признака у самок больше, чем у самцов.
Узкая норма реакции обеспечивает широкую фенотипическую дисперсию мужского пола. Она дает возможность подвергнуть элиминации и дискриминации в основном мужской пол. При этом, бóльшее сечение канала связи мужского пола с потомством позволяет максимально тиражировать отобранные, самые подходящие, генотипы.
“ ...если объединить всех самцов популяции в мужскую команду, а всех самок—в женскую и устроить “личные и командные соревнования”, то чемпионами в “личном первенстве” по всем программам были бы самцы, а в “командном зачёте” победителями оказывались бы самки”
Геодакян, 1966.
И наоборот, широкая норма реакции женского пола сужает его фенотипическую дисперсию и позволяет сохранить его численность и существующее распределение генотипов. На первый взгляд кажется, что чем шире норма реакции, тем шире должна быть и фенотипическая дисперсия. Однако это не так. На самом деле все наоборот: в популяции фенотипическая дисперсия мужского пола, состоящего из особей с узкой нормой реакции, будет шире, так как она сохранит генотипическую дисперсию, а фенотипическая дисперсия женского пола, состоящего из особей с широкой нормой реакции, будет уже, так как широкая норма реакции даст возможность в условиях стабилизирующего отбора, крайним генотипам “уйти” из зон “дискомфорта”, “подтянуться” к фенотипической норме (моде) популяции и таким образом сгладит генотипическую дисперсию (Рис. 7.2). Именно высокая онтогенетическая пластичность женского пола обеспечивает ему высокую стабильность в филогенезе. Следовательно, в филогенетическом плане более изменчив, пластичен мужской пол, а в онтогенетическом, наоборот, более пластичен и изменчив женский. Такое, на первый взгляд парадоксальное, распределение ролей в филогенезе и онтогенезе на самом деле последовательно и непротиворечиво: оно реализует единую идею специализации полов по консервативной и оперативной задачам эволюции. Несколько утрируя, можно сказать, что информационные взаимоотношения популяции со средой строятся на элиминации мужских особей и “воспитании” (онтогенетическом сдвиге) женских.Гипотеза более широкой нормы реакции женского пола позволяет делать довольно четкие предсказания. Например, у особей мужского пола доля “наследственной компоненты” должна быть больше, а “средовой”—меньше, чем у особей женского пола. Если сравнить внутрипарную и межпарную дисперсию у однояйцевых (монозиготных) близнецов мужского и женского пола, то внутрипарная должна быть больше у женских пар, а межпарная—у мужских, то есть степень конкордантности у однояйцевых близнецов мужского пола должна быть больше, чем у таких же близнецов женского пола. Далее, фенотипическая дисперсия в чистой линии может быть относительно шире у женского пола, а в полиморфной (дикой) популяции она шире у мужского.
Ванденберг и др. (Vandenberg et al., 1962) исследовали внутрипарные различия у 23 пар мужских однояйцевых близнецов и 21 пары женских по различным измерениям: антропологическим, сердечно-сосудистым, гематологическим, биохимическим, психологическим и др. Возраст исследованных пар от 12 до 18 лет. По 185 переменным внутрипарный разброс оказался статистически значимо больше у женских пар, по 41—у мужских, а по 353 переменным не было обнаружено значимой разницы. Аналогичные результаты были получены и другими авторами (Никитюк, 1977; Chovanova, 1980).
Исследование степени разнообразия по 9 параметрам (социальные, поведенческие и умственные способности в возрасте 2, 3 и 4 года) у 2000 пар однояйцевых близнецов показало большее разнообразие в парах девочек Интересно, что в полном соответствии с представлениями теории у дизиготных близнецов наблюдалась обратная корреляция по тем же признакам. (Loat et al., 2004).
Шюлер Л. и др. (1976) обнаружили различия между самцами и самками мышей в определении веса эндокринных органов. Доля генотипической компоненты в фенотипической изменчивости у самцов по всем изученным признакам была больше, чем у самок, что также подтверждает гипотезу нормы реакции.
Более широкая фенотипическая дисперсия мужского пола выявляется в различных тестах по определению коэффициента интеллектуальности (IQ). У женщин показатели IQ группируются вокруг некоторых средних величин, а у мужчин—наблюдается значительный разброс по этому показателю (Hurt, 1978).
“Мужчины либо очень умны, либо уж очень глупы”:
...среди умных двух процентов населения мужчин в два раза больше, чем женщин.
... среди наименее интеллектуальных двух процентов населения мужчин в два раза больше, чем женщин.
Deary et al., 2007.
Психологи из Эдинбургского университета проверили способности у более чем 2500 братьев и сестер, подвергнув их тестам по информатике, математике и английскому, а также оценив их способность к выполнению механических операций. Мужчины набрали максимальное и минимальное количество очков по всем тестам, они также лучше справились с заданиями по информатике и арифметике. Женщины получили средние оценки и выглядели лучше в языковых тестах. В два раза больше мужчин чем женщин оказалось на периферии распределения—как среди двух процентов умных, так и среди двух процентов наименее интеллектуальных испытуемых (Deary et al., 2007).Американские исследователи проверяли математические способности (SAT тест) учеников 7-го класса на крайней правой границе распределения (верхний 0.01 % популяции). В начале 80-х годов на каждую девочку приходилось 13 мальчиков. К 1991 г. разница уменьшилась до 4 : 1 и оставалась на этом же уровне в последующие годы. По тесту на математическое и научное рассуждение (ACT тест) соотношение было 3 : 1, при этом из 19 учеников, набравших наилучший результат, было 18 мальчиков (Wai et al., 2010).
Гипотеза широкой нормы реакции женского пола распространяет эволюционную концепцию дифференциации полов на онтогенез, связывая в единое целое филогенетические и онтогенетические закономерности раздельнополости и взаимоотношения популяции со средой. Она позволяет объяснить феномен дифференциальной смертности полов, и многие другие непонятные онтогенетические явления, затрагивающие в разной степени мужской и женский пол, например полимерию, пенетрантность, экспрессивность, а также психологические особенности полов.
Приведенные рассуждения позволяют построить логическую цепочку явлений, приводящих от разной нормы реакции полов к популяционному половому диморфизму: чем шире норма реакции, тем больше онтогенетическая пластичность, тем меньше фенотипическая дисперсия и меньше смертность; чем меньше смертность, тем лучше сохраняется генотипическая дисперсия и тем меньше филогенетическая пластичность, что и приводит к возникновению полового диморфизма.