
Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году в сборниках по проблемам кибернетики. Тогда же вышла статья
| Вид материала | Статья |
СодержаниеТри основных схемы размножения Половой процесс Чередование поколений и эволюция размножения |
- Вигена Артаваздовича Геодакяна, посвященные проблеме пола, появились в 1965 году, 4857.78kb.
- I вступление, 32.31kb.
- Культура Кольского Севера; Творческие работы посвященные проблемам студенческой молодежи, 67.08kb.
- «Мой учитель» посвященные к Году учителя, 64.26kb.
- Михаил Александрович Шолохов (родился. В 1905 году) Когда в 1925 году появились Донские, 334.1kb.
- Лекция 21. Генетика пола Определение пола, 33.63kb.
- Публичный доклад о результатах деятельности школы моу сош п. Пола в 2008-2009 учебном, 335.49kb.
- Курс «Обзор перспективных технологий Microsoft. Net» Губанов Ю. А., математико-механический, 177.56kb.
- Больше-Кочинский школьный музей этнографии и фольклора, 25.15kb.
- Воспитание и обучение одаренных детей, 267.86kb.
Три основных схемы размножения
Первым важным событием в эволюции размножения было возникновение процесса скрещивания (оплодотворения)—слияния генетической информации двух родителей. С появлением скрещивания огромное многообразие форм размножения, существующее в растительном и животном мире, разделилось на два главных типа—бесполое (нет скрещивания) и половое (есть скрещивание) (Рис. 5.2). По нашей классификации партеногенез следует считать формой бесполого размножения.
Р
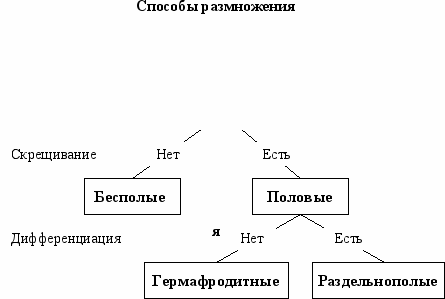
ис. 5.2 Классификация способов размножения (Геодакян, 1983,1989).
В бесполом размножении участвует только одна родительская особь, которая производит себе подобных. В половом размножении участвуют два родителя. Принципиальное значение имеет, однако, не количественная сторона: то что в одном случае получается “из одного—два”, а в другом—“из двух—три”. Гораздо важнее качественная сторона дела, а именно то, что при бесполом варианте не возникает новых свойств, в то время как при половом каждый раз появляется иное качество, отличное от родительского. Это очень существенно. Половое размножение обязательно предполагает скрещивание, благодаря которому возникают новые варианты. Оно необходимо для обеспечения генетического разнообразия.
Вторым важным событием, видимо, следует считать появление половой дифференциации—разделения на мужской и женский пол, с наложением запрета на скрещивания между особями одинакового пола. С возникновением половой дифференциации, половые способы, в свою очередь, разделились на две формы: гермафродитные (нет дифференциации) и раздельнополые (есть дифференциация).
Таким образом, в зависимости от наличия или отсутствия двух фундаментальных признаков размножения—скрещивания и дифференциации полов, существующие способы размножения можно разделить на три основных типа: бесполый, гермафродитный и раздельнополый (Рис. 5.2). В Табл. 5.1 приведены принципиальные схемы и некоторые характеристики этих типов.
Табл. 5.1 Основные типы размножения и некоторые их характеристики
| Тип размножения | Программа | Источник разнообразия* | ||
| репродукции | реком- бинации | эволюции | ||
| | | | | |
| Бесполое | max | mid | min | MUT |
| Гермафродитное | mid | max | mid | MUT+REC |
| Раздельнополое | min | mid | max | MUT+REC+DIF |
* MUT—мутации, REC—рекомбинация, DIF—дифференциация.
Преимущества бесполого размножения связаны, прежде всего с его простотой (не требуется находить партнера) и высокой эффективностью (потомство может оставить любая, отдельно взятая особь в любом месте). Ещё одно преимущество связано с тем, что ценные сочетания генов, появившись в результате мутаций, не рассыпаются, а передаются из поколения в поколение в неизмененном виде.
Недостаток бесполого размножения в том, что источником необходимого для естественного отбоpa разнообразия (изменчивости) являются только мутации. Поэтому, если две благоприятные мутации а → А и в → В, возникают у разных особей бесполой популяции, то способа, посредством которого обе эти мутации могли бы оказаться в генотипе одного из потомков этих особей не существует. Особь с генотипом АВ может появиться лишь в том случае, если вторая мутация В произойдет у одного из потомков исходного мутанта А, или наоборот, если мутация А возникнет у потомка мутанта В (Fisher, 1930; Muller, 1932).
C возникновением полового процесса—скрещивания, у половых форм преимущества бесполого способа утрачиваются, но появляется новый, фактически неисчерпаемый источник разнообразия—комбинаторный потенциал популяции. Эволюционные преимущества рекомбинации не до конца понятны, поскольку замечено, что полезные комбинации, созданные в одном поколении, могут быть разрушены в следующем (Michod, 1987). К преимуществам рекомбинации относят также возможность компенсации повреждений генов.
Половой процесс и половая дифференциация явления разные и, по своей сути, диаметрально противоположные. Первый создает (повышает) разнообразие генотипов, и в этом его эволюционное предназначение, вторая, наоборот, снижает его вдвое, и, в чем её эволюционная роль, никто не может объяснить.
Чередование поколений и эволюция размножения
Многие организмы могут размножаться как бесполым, так и половым путем. При этом говорят о разных поколениях данного вида. Если они закономерно сменяют друг друга, такое явление называется чередованием поколений.
Чередование поколений встречается как у растений так и у животных и имеет много общего. Границей, разделяющей половое и бесполое поколение в цикле развития является процесс оплодотворения (Рис. 5.3 Приложение В). При оплодотворении в результате слияния гаплоидных гамет появляется диплоидная зигота и половое поколение переходит в бесполое. И бесполое и половое поколения могут иметь как одинарный, так и двойной набор хромосом в зависимости от того, на какой стадии жизненного цикла происходит процесс мейоза. При мейозе число хромосом уменьшается вдвое и диплоидный набор переходит в гаплоидный. В процессе эволюции в цикле развития закономерно уменьшается роль (продолжительность существования и размеры) гаплоидной фазы и увеличивается роль диплоидной (Рис. 5.4 Вилли, Детье, 1975).
Р
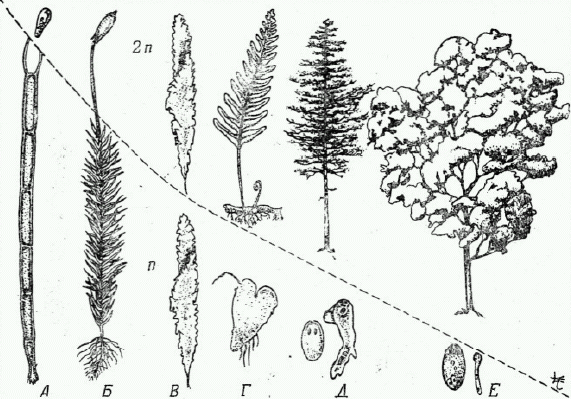
ис. 5.4
Эволюция бесполого (2 n) и полового (n) поколений у растений (из Вилли, Детье, 1975).
А. Водоросли (Oedogonium). Б. Мхи. В. Водоросли (Ulva). Г. Папоротники. Д. Голосеменные. Е. Покрытосеменные.
Чередование поколений зависит от условий среды. В благоприятных условиях размножение происходит, как правило, бесполыми способами (делением, почкованием, вегетативно, партеногенетически). При наступлении неблагоприятных условий или незадолго перед этим, происходит смена бесполого поколения на половое. Например, у многих водяных блох и тлей при благоприятных условиях (обычно летом) имеет место партеногенез. Молодь—исключительно самки—вылупляется из “летних” яиц с мягкой оболочкой. При наступлении менее подходящих условий производится несколько самцов, которые затем оплодотворяют самок обычным путем. Оплодотворенные самки откладывают “зимние” яйца с твердой оболочкой, которые могут сохраняться в течение длительных периодов неблагоприятных условий (Кэйн, 1958). Тля Therioaphis trifolii в Северной Америке выработала устойчивость к инсектицидам и перешла с партеногенеза на половое размножение (Blackman, 1981). Когда клетки дрожжей имеют достаточно питательных веществ, они размножаются бесполым путем, недостаток питания запускает половое размножение (Goddard M., 2005).
Другой пример—описанные М. М. Завадовским (1923) опыты Кана и Таусена которые, перемещая коловратку из прудовой воды в речную или колодезную и обратно, наблюдали на 3-й и 4-й день появление самцов, ранее отсутствовавших. При этом характерно, что направление перемещения безразлично (из прудовой воды в речную или наоборот).
Аналогичная ситуация наблюдается у различных видов и популяций низших ракообразных, у которых есть все переходы от чистого партеногенеза (без самцов) через разные ступени сезонного партеногенеза, когда самцы появляются при ухудшении условий обитания, и до популяций, у которых самцы присутствуют все время. Интересно при этом, что чем стабильнее условия среды, тем меньше в популяции самцов. Поэтому в мелких пересыхающих водоемах встречаются, как правило, популяции с самцами, а в крупных, характеризующихся относительной стабильностью условий обитания, превалируют партеногенетические формы (Мануйлова, 1965). Например, из 25 видов семейства Polyphemidae, населяющих Каспийское море, несмотря на специальные поиски, только у восьми видов обнаружены самцы (Мордухай-Болтовской, 1965а).
Эволюция размножения шла от бесполых форм к половым. Примитивные формы размножаются только бесполым путем, у более сложных бесполое размножение чередуется с половым. Наиболее прогрессивные виды размножаются только половым путем (Рис. 5.3Д Приложение В). Таким образом, чередование поколений можно рассматривать как переходный этап в эволюции размножения.