
Адаптации ксерофитов Предкавказья в связи с эволюцией экологической группы 03. 00. 16 экология
| Вид материала | Автореферат |
Содержание5.1.Анализ анатомо-морфологических и физиологических адаптаций |
- Региональный конкурс учебно-исследовательских экологических проектов школьников 2011, 103.07kb.
- Главы Администрации Ростовской области от 17. 04. 1997 n 170 "О проведении Дней защиты, 74.25kb.
- Хищные птицы и совы трансформированных степных экосистем предкавказья 03. 02. 08-03, 1415.26kb.
- Психофизиологические основы адаптации и дезадаптации, 377.67kb.
- Шапша Доклад «Воспитание экологической культуры в процессе обучения», 93.62kb.
- Дохристианские славянские верования, 222.09kb.
- Положение о Городской школьной научно практической конференции «Экология большого города», 43.27kb.
- Утверждено решением Кировского областного, 30.66kb.
- Экология питания и механизм адаптации к среде обитания моих предков- жителей Приилимья, 67.01kb.
- Выполни задание. Подумай, какие экологические проблемы возникают на всех этапах производства, 48.27kb.
В главе рассматриваются представления об адаптивных стратегиях с точки зрения экологической физиологии растений. В рамках этих представлений рассматриваются механизмы взаимодействия нескольких донорно-акцепторных систем (ДАС) (Р.Уиттекер, 1980; Б.М.Миркин, Л.Г.Наумова, 1998). Проблема классификации растений по признакам сходства различия их функционирования в экосистемах не нова. Отличительной чертой таких классификаций является поиск систем адаптивности растений к разным местообитаниям через особенности их функционирования, т.е. способность выживать, используя различные морфологические и физиологические механизмы растений их жизненных стратегий (Р.Уиттекер, 1980; Б.М.Миркин, Л.Г.Наумова, 1998 Л.Г.Раменский, Глизон 1975,1987). Совершенно очевидно, что объектом классификации является все многообразие первичных адаптивных реакций (ПАР) на условия обитания – от молекулярно-биологических до онтогенетических. Совокупность частных ПАР складывается из двух множеств: множества видоспецифичных комбинаций, поскольку каждый вид наделен уникальным набором ПАР к меняющимся условиям среды. Это множество описывается через «принцип экологической индивидуальности видов Раменского-Глизона (Л.Г.Раменский, Р.Глизон 1975,1987). В данном
 случае при исследовании ксерофитов как экологической группы наблюдается дивергенция признаков. Например, в условиях острого дефицита воды суккуленты Sempervivum caucasicum, S. pumilum снижают расход воды, редуцируя листья и усиливая кутикулу, а Calligonum aphyllum в аналогичных условиях образует очень глубоко проникающую корневую систему. Существование двух пересекающихся множеств первичных адаптивных реакций: по видовой принадлежности и к факторам среды – ставит проблему использование в качестве основы уже существующих классификаций и создание систем нового типа. (Р.Уиттекер, 1980). Синтаксономия также не может быть использована для этой цели, поскольку в разных сообществах популяции одного и того же вида могут проявлять разные черты. Например, «хорионы», предложенные Ю.Р. Шеляг-Сосонко (1969). По тем же причинам нельзя использовать и классификацию жизненных форм. Часто можно найти много общего в реакциях таких далеких форм, как фанерофиты и однолетние терофиты, например, черты рудеральности у многих маревых, например, Polycnemum arvense, Chenopodium foliosum, Atriplex sagittata, Сeratocarpus arenarius, Kochia prostrata, Kalidium foliatum, Halocnemum strobilaceum и многих других.
случае при исследовании ксерофитов как экологической группы наблюдается дивергенция признаков. Например, в условиях острого дефицита воды суккуленты Sempervivum caucasicum, S. pumilum снижают расход воды, редуцируя листья и усиливая кутикулу, а Calligonum aphyllum в аналогичных условиях образует очень глубоко проникающую корневую систему. Существование двух пересекающихся множеств первичных адаптивных реакций: по видовой принадлежности и к факторам среды – ставит проблему использование в качестве основы уже существующих классификаций и создание систем нового типа. (Р.Уиттекер, 1980). Синтаксономия также не может быть использована для этой цели, поскольку в разных сообществах популяции одного и того же вида могут проявлять разные черты. Например, «хорионы», предложенные Ю.Р. Шеляг-Сосонко (1969). По тем же причинам нельзя использовать и классификацию жизненных форм. Часто можно найти много общего в реакциях таких далеких форм, как фанерофиты и однолетние терофиты, например, черты рудеральности у многих маревых, например, Polycnemum arvense, Chenopodium foliosum, Atriplex sagittata, Сeratocarpus arenarius, Kochia prostrata, Kalidium foliatum, Halocnemum strobilaceum и многих других.В результате полевых наблюдений была проведена инвентаризация ксерофитов Предкавказья и сделана попытка проанализировать различные приспособления, связанные с проявлениями типов эколого-ценотических стратегий. У видов с разными типами стратегий выделяются различные способы адаптаций: показано, что тип адаптивной стратегии может реализоваться через подавляющее большинство проявлений жизнедеятельности популяций (структуру популяции, жизненный цикл, дифференциацию ниш и конкурентоспособность, репродуктивные процессы, реакцию на экологический стресс и т.д.). На основании составленного нами конспекта ксерофитов Предкавказья (486 видов) было выделено 2 основные экологические группы, характеризующиеся различными адаптивными стратегиями: патиенты и эксплеренты (табл. 7).
Таблица 7. Соотношение различных экологических групп флоры ксерофитов Предкавказья.
| Экологическая группа | Кол-во видов | Процент участия, % |
| Патиенты | 454 | 93,4 |
| Эксплеренты | 32 | 6,6 |
Нами установлено, что патиентов во флоре ксерофитов Предкавказья 454 вида, тогда как эксплерентов 32 вида. Следовательно, процент участия патиентов во флоре Предкавказья составляет 93,4 %, а эксплерентов 6,6 % .
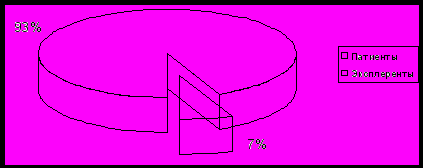
Диаграмма 8 Процентное соотношение патиентов и эксплерентов флоры ксерофитов Предкавказья.
На основании полученных результатов, мы можем сделать вывод, что патиенты являются преобладающей экологической группой вследствие более высокой экологической пластичности. Под влиянием внешних условий устойчивость растений может меняться, и происходит смещение точек максимума и минимума, изменение ширины интервала, в пределах которого развитие растений происходит без повреждений.
5.1.Анализ анатомо-морфологических и физиологических адаптаций
Ксерофиты – растения сухих местообитаний, способные благодаря ряду приспособительных признаков и свойств переносить перегрев и обезвоживание, поэтому для них характерны особые анатомо-морфологические, экологические, физиологические, биохимические адаптации (В.А.Алексеев, 1948; Е.Н.Максимов, 1952; П.А.Генкель, 1967; М.В.Гусев, 1974; В.Н.Жолкевич, 1984; Ю.В.Гамалей, 2003). Все приспособления у ксерофитов на наш взгляд возникали по двум направлениям:
- Регулирование транспирации при дефиците влаги.
- Развитие различных приспособлений, служащих для накопления, или для сохранения воды. Часто один и тот же вид ксерофитов представляет соединение нескольких приспособлений, но удобнее в целях наглядности разобрать их в отдельности.
Для регулирования транспирации в конкретных экологических условиях Предкавказья, или для предотвращения последствий увеличения транспирации, в процессе эволюции возникли следующие приспособления:
Уменьшение площади поверхности листа. Многие ксерофиты имеют мелкие, узкие, сильно редуцированные листовые пластинки. Это особенно наглядно при сравнении облика родственных видов из сухих и влажных местообитаний: достаточно вспомнить, например, мелкие рассеченные листья P.astrachanica и крупные листовые пластинки Potentilla argentea, обитающей на более сырых лугах и опушках; узкие проволоковидные листья F.valesiaca и листья – Festuca pratensis. В особо засушливых пустынных местообитаниях листья некоторых древесных и кустарниковых пород редуцированы до едва заметных чешуек, некоторые стипаксерофиты сухую, жаркую погоду при дефиците влаги способны сворачивать в свои листья, так что даже широкие листья становятся трубчатыми или нитевидными. Это явление наблюдается у злаков, растущих на солончаковой почве, например, Stipa capillata, Stipa dasyphylla. Защита листьев от потерь влаги на транспирацию достигается благодаря развитию мощных покровных тканей – толстостенного, иногда многослойного эпидермиса, часто несущего различные выросты и волоски, которые образуют густое «войлочное» опушение поверхности листа. Сильно опушенные листья имеют степные ксерофиты: Nepeta kubanica, Salvia aethiops, S.tesquicola, S.nemorosa, S.fugax. Интенсивность освещения, действующая на фотосинтезирующие органы, и органы транспирации связаны с профильным положением. У таких афилльных видов фотосинтез осуществляют зеленые ветви, например: Ephedra distachia, Calligonum aphyllum, Corispermum marschallii, Salicornia europaea. Сюда относятся многие растения со сложными и простыми листьями, особенно из числа низких кустарников, например, многие виды Ephedra distachia, Asphodeline taurica. Некоторые двудольные растения осуществляют движения листьев, опушения например, Isatis sabulosa, Crambe cordifolia. Различаются формы и видоизменения листа у ксерофитов. Игловидный или пиноидный лист, имеет длинную, линейную, острую форму и ассимилирующая ткань расположена более или менее в центре; ложбинок или кроющих волосков не бывает (Juniperus oblonga, J.sabina). Эрикоидный лист – это завернутый лист, края которого свернуты вверх или вниз; образуется, таким образом, покрытая опушением бороздка, в которой скрыты устьица; устьица могут быть приподняты над уровнем эпидермиса. Эрикоидные листья часто коротки, линейны. У других видов поверхность листа покрыта слоем толстой кутикулы или воскового налета. Можно упомянуть о форме листа у суккулентов семейства толстянковые. Так как здесь лист часто является, независимо от толщины, более или менее цилиндрическим, линейным, удлиненным или лопатковидным, не имеет ни зубчиков, ни других надрезов, например, Sempervivum caucasicum, S.pumilum, Hylotelephium caucasicum, Sedum caucasicum, S.spurium, S.subulatum (см. табл. 8).
Таблица 8. Число устьиц на 1 мм2 поверхности и длина жилок мм2 на см2 у мезофитов и ксерофитов.
| Вид | Длина жилок, мм/см | Число устьиц на 1мм поверхности | |
| нижней | верхней | ||
| Мезофиты: Dactylis glomerata – ежа сборная Festuca pratensis – овсяница луговая Phleum pratense – тимофеевка луговая Bromopsis inermis – костер безостный Bromopsis riparia – костер береговой Ксерофиты: Phleum phleoides – тимофеевка степная Stipa capillata – ковыль волосатик Stipa lessingiana – ковыль Лессинга K. sabuletorum – тонконог песчаный Koeleria luersennii – тонконог Люерсена Koeleria cristata – тонконог гребенчатый Festuca sclerophylla – беломятлик жестколистный Festuca valesiaca – овсяница валезская Festuca ovina – овсяница овечья Elytrigia gracillima – пырей стройный Elytrigia elongata – пырей удлинённый Elytrigia pseudocesia – пырей ложносизоватый Elytrigia intermedia (Host) Nevski – пырей средний | 172 268 307 310 314 420 660 650 732 657 745 660 534 654 657 657 564 678 | 37 104 47 48 72 132 153 156 163 176 178 168 176 178 210 | 0 0 0 0 0 0 0 110 102 104 137 123 134 123 140 |
Интересный материал дает сравнение близкородственных видов. Так при сравнении изменения степени ксерофильности у 5 видов подорожниковых встречающихся в Предкавказье (табл. 9) можно выявить четкие различия между видами одного рода по их анатомо-морфологическим признакам и транспирации.
Таблица 9. Изменение степени ксерофильности у некоторых видов подорожников.
| Вид | Плотность жилок мм /см | Число устьиц на 1 мм | Транспирация, % |
| Plantago tenuiflora P. cornuti P. maxima P. salsa P. lanceolata L. | 543 601 405 476 689 | 432 546 384 465 478 | 65 78 34 56 100 |
Одними из объектов нашего исследования в сравнении двух важных физиологических процессов фотосинтеза и дыхания служили различные виды ксероморфных и мезоморфных злаков, обитающих повсеместно в Предкавказье. Все опытные растения мы расположили следующим образом в порядке нарастающей степени их ксерофилизации. В связи с поставленной задачей сравнительного изучения физиологических особенностей у ксерофитов и мезофитов в исследовании брались представители обоих экологических типов растений, в большинстве случаев принадлежащих к одному роду. Для выяснения зависимости фотосинтеза мезоморфных и ксероморфных злаков от водного режима были поставлены две серии опытов. В первой серии опытов, изучалась зависимость фотосинтеза от водного режима внешней среды в течение всей вегетации, опытные растения выращивались в опытных вегетационных сосудах Вагнера в условиях различной влажности почвы: 70 и 25% от полной влагоемкости почвы. Вторая серия опытов была поставлена для выяснения степени влияния оводненности ассимилирующих тканей на фотосинтезирующую активность сравнительно у ксерофитов и мезофитов. Методика определения фотосинтеза проводилась на отрезанных листьях (Н.Бриллиант, 1936). Способность к фотосинтезу и дыханию в летних опытах определялась широко известным эвдиометрическим методом с последующим анализом газа прибором Р.Бонье и Х.Манжен (1926). При сравнении более ксероморфного костра берегового с мезоморфным костром безостым мы видим, что ксерофит обладает более высокой интенсивностью фотосинтеза. При сравнении более ксероморфного костра берегового с мезоморфным костром безостым мы видим, что ксерофит на обеих влажностях почвы обладает более высокой интенсивностью фотосинтеза. Если же сопоставлять показатели фотосинтеза в условиях разной влажности почвы, видно, что у костра берегового более низкой влажности отвечает несколько большая величина фотосинтеза, тогда как у костра безостого мы имеем некоторое снижение фотосинтеза (на 7-13%) на 25% влажности почвы (табл. 10).
Таблица 10. Интенсивность фотосинтеза и дыхания у мезофитов и ксерофитов в условиях разной влажности почвы: 70% и 25% от полной влагоемкости
(в мг СО2 на 100 см2 за 1 час).
| Вид | Фотосинтез | Дыхание | ||||
| | влажность почвы % при 70% (%) | ксерофит в % от мезофита | влажность почвы при 25% | влажность почвы 70% | влажность почвы 25% | ксерофит от мезофита в % |
| 1.Костер безостый 2.Костер береговой 3.Тимофеевка луговая 4.Тимофеевка степная 5.Ежа сборная 6.Тонконог гребенчатый 7.Овсяница луговая 8.Овсяница валезская | 10.1 14.8 6.2 10.1 10.1 12.4 10.7 10.4 | 100 108 103 85 94 67 105 84 | 91 108 101 95 94 67 67 84 | 1.1 0.9 1.2 1.3 0.9 2.3 2.4 2.3 | 1.6 1.3 1.8 1.4 1.0 1.1 1.3 2.6 | 100 172 109 107 115 256 191 114 |
Костер береговой обладает также и более высокой интенсивностью дыхания по сравнению с костром безостым как на 70%, так и на 25 % влажности почвы, причем на низкой влажности почвы у обоих костров дыхание несколько повышено. Опытные данные, относящиеся к двум видам тимофеевки, показывают, что в условиях оптимальной влажности почвы ксероморфная степная тимофеевка обладает более высокой фотосинтетической способностью, чем тимофеевка луговая. У тимофеевки степной в условиях низкой влажности почвы фотосинтез во всех случаях был выше фотосинтеза тимофеевки луговой (на 31% в среднем). На снижение влажности почвы тимофеевка степная реагировала почти одинаковым снижением фотосинтеза. Дыхание тимофеевки степной всегда несколько превышало дыхание тимофеевки луговой. Однако недостаток влаги в почве оба вида, в противоположность кострам, реагировали почти одинаковым снижением интенсивности дыхания. При сопоставлении величин фотосинтеза и дыхания у тонконога и ежи мы видим, что интенсивность обоих процессов у ксерофита всегда выше, чем у мезофита. На пониженное увлажнение почвы ксероморфный тонконог и мезоморфная ежа реагируют по-разному. Так, фотосинтез тонконога, выращенного на низкой влажности почвы, составлял в среднем 65-46% от интенсивной ассимиляции тонконога, находившегося в условиях оптимального увлажнения, а дыхание 69-81%.Таким образом, можно сделать следующие выводы:
- газообмен у исследованных ксероморфных злаков, обитающих в условиях оптимального увлажнения, находится на более высоком уровне, чем у мезофитов;
- ксерофиты, обитающие в условиях низкой влажности почвы, также показали в основном более высокую интенсивность фотосинтеза и дыхания по сравнению с соответствующими мезофитами;
- на уменьшение влажности почвы различные типы ксерофитов и мезофитов реагируют по-разному.
Большинство луговых ксерофитов (тимофеевка степная, тонконог гребенчатый и овсяница валезская) снижает интенсивность своего ассимиляционного процесса в значительно большей степени, чем мезофиты. Изложенные выше данные дают полное право заключить, что длительная адаптация к разнообразным экстремальным условиям среды привела к такой физиолого-структурной перестройке растений, которая способствовала повышению с одной стороны, трансформации необходимых для жизнедеятельности внешних факторов, с другой - уровня их гомеостаза для обеспечения нормального хода процессов жизнедеятельности и устойчивости к указанным неблагоприятным условиям. Видимо, в первые годы жизни, у молодых растений под действием неблагоприятных факторов нарушается нормальный ход жизнедеятельности, вследствие чего значительно ослабляется синтез хлорофилла. При этом в качестве компенсационной реакции увеличивается содержание слабосвязанной с липопротеидным комплексом формы хролофилла, способствуя тем самым активации фотосинтеза. В дальнейшем, с возрастом растет синтез хлорофилла, что способствует ослаблению экстремальных факторов, влияющих на его распад, увеличивается прочность его связи с липопротеиновым комплексом. Данные по хлорофиллу подтверждают наши предположения о том, что у растений, находящихся в восходящем периоде онтогенеза, адаптивные реакции к неблагоприятным условиям направлены к повышению трансформационной способности
Нами исследованы физиологически активные соединения содержащиеся в различных экологических группах ксерофитов Предкавказья. Для данного исследования были выбраны модельные виды с учетом максимально полного отражения экологического разнообразия представленных ксерофитов. Растения были разделены на 4 группы по распространению, встречаемости, экологической приуроченности видов.
1 группа представлена характерными для Предкавказья стипоксерофитами -Stipa capillata L. - Ковыль волосатик, S. lessingiana - К. Лессинга, K. sabuletorum - песчаный, E. elongata - П. удлинённый,
2 группа представлена настоящими ксерофитами -Chenopodium foliosum - Марь олиственная, Ch. glaucum L. - М. сизая, Ch. glaucum L. - М. Сизая, Ch. botrys L. - М. душистая, Atriplex sagittata - Лебеда блестящая, Atriplex sagittata - Лебеда блестящая,
3 группа представленна суккулентами -Sempervivum caucasicum - Молодило кавказское, S. oppositifolium - О. супротиволистный, S. subulatum - О. шиловидный,
4 группа представлена схожей группой ксерофитов суккулентами имеющими ряд дополнительных приспособлений в виде млечников участвующих в биохимических реакциях данной группы растений- Euphorbia glareosa - Молочай хрящеватый, E. stepposa - М. степной, E. procera - М. мохнатый, E. petrophila - М. скалолюбивый, E. boissieriana Woronow - М. Буасье.
При оценке биологической активности растительных выделений, почвы мы использовали биотесты, которые были достаточно универсальными, высоко чувствительными и краткосрочными, так как работа проводилась в нестерильных условиях и состав изучаемых веществ мог быстро измениться. Используемые нами биотесты просты в осуществлении, позволяют провести необходимое для математической обработки число параллельных повторений. В анализе мы учитывали существование и других взаимодействующих факторов, например, поглотительную способность почв, деструктивное действие микрофлоры.
Таблица 11. Средние показатели влияния водных вытяжек разных групп ксерофитов на развитие корневой системы биотестов (огурец “Мироновский - 1”).
| Средняя длина корней растений биотестов | Вода конц. 0; контроль | Вытяжка конц. 1:1 1группа | Вытяжка конц. 1:1, 2 группа | Вытяжка конц. 1:1, 3 группа |
| L 1 сут. мм | 40.7 | 43.4 | 42.2 | 39.2 |
| L 2 сут. мм | 75.9 | 81.7 | 63.6 | 75.9 |
| d L 2 мм | 35.2 | 38.3 | 21.4 | 36.7 |
| L 3 сут. мм | 129.6 | 113.5 | 114.7 | 114.5 |
| d L 3 мм | 53.7 | 31.8 | 51.1 | 38.6 |
| L б.к. 3 мм | 17 | 14.7 | 23.2 | 15 |
| L 4 сут. мм | 196 | 196.1 | 160.1 | 197.4 |
| d L 4 мм | 61 | 82.6 | 45.4 | 82 |
| L б.к. 4 мм | 52.5 | 60 | 60 | 52.8 |
| dL мм | 149.9 | 152.7 | 117.9 | 158.2 |
Примечание:, L - длина корней в мм, dL - прирост корней в мм, L б.к. - длина боковых корней.
Дальнейшие исследования показали, что основными источниками физиологически активных веществ исследуемых экологических групп ксерофитов являются вегетирующие листья, корневые остатки и прижизненные корневые выделения в процессе нормального метаболизма растений. Наиболее высокой токсичностью отличались вытяжки (1:10) из листового опада и зеленых листьев исследуемых групп ксерофитов (табл. 11). Эти вытяжки имели слабокислую реакцию (рН 5.3 - 5.5) и их ингибирующий эффект нельзя отнести за счет концентрации водородных ионов, так как с увеличением разведения их активность падает, а рН не изменяется. В листовом опаде и листьях исследуемых ксерофитов содержится большое количество легко экстрагируемых водой органических соединений. Высокое содержание органических веществ в экстрактах из листового опада и зеленых листьев в значительной степени определяет их токсичность. Водные экстракты (1:10) также задерживали прорастание семян растений - биотестов и семян исследуемых групп растений. Вытяжки из корневого опада (1:10) подавляли в больших концентрациях рост растений биотестов. В процессе разложения корневых остатков исследуемых растений на протяжении 6 месяцев происходит повышение их токсичности. Это вызвано, по-видимому, образованием в процессе разложения корней новых токсичных веществ [6].
Основное внимание мы уделяли изучению водорастворимых физиологически активных веществ, которые в исследуемых нами группах ксерофитов были выявлены и идентифицированы как вещества фенольной природы. Как нами было установлено, фенольные соединения в зависимости от химического строения и концентрации ингибируют или стимулируют рост растений биотестов. Они, вероятно, выполняют функцию промежуточных компонентов в процессе образования гумусовых веществ. Чтобы определить, какое влияние оказывают эти соединения на растения, они нами были количественно определены в почвенном опаде ксерофитов на стационарных площадках в осенний и весенний периоды. За основу нашего метода был взят механизм десорбции органических веществ из почвы с помощью незаряженного катионита КУ- 2-8, функциональная сульфогруппа которого имеет достаточную плотность зарядов, чтобы отделить от почвенного поглощающего комплекса (ППК) подвижные фенольные соединения. Такой подход, на наш взгляд, в какой-то степени моделирует растворяющую и поглощающую способность корней.
Содержание фенольных соединений в почвенном опаде исследуемых ксерофитов на 4-х стационарных площадках, в весенний и осенний периоды математически обработаны и представлены на диаграммах (9, 10) и в таблицах (12, 13, 14, 15). На диаграммах видно, что наибольшее содержание фенолов поддающихся десорбции из почвы с помощью ионообменной смолы КУ- 2 - 8, илюируется смесью растворителей ацетон: вода (1:2). В почве, где произрастают исследуемые нами ксерофиты, фенолов содержится соответственно в первой группе-117 мг, во второй группе- 246,4 мг; в третьей группе- 260 мг; в четвертой группе - 307,8 мг на 100 г. почвы в весенний период.
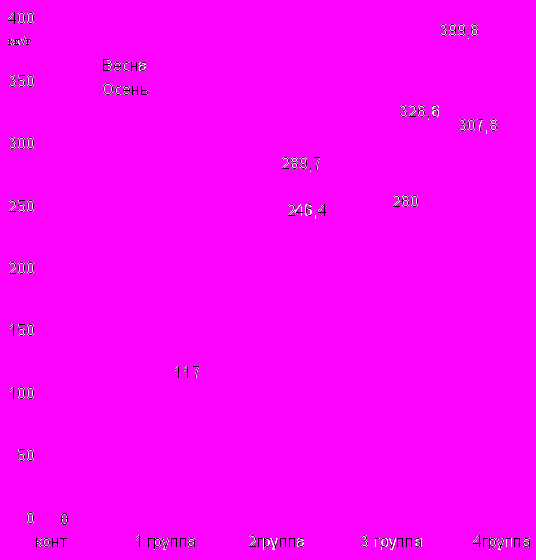
Диаграмма 9 Содержание фенолов в почвенном опаде исследуемых групп ксерофитов.
| Таблица 12. Содержание фенолов в почвенном опаде исследуемых групп ксерофитов в весенний период. | ||||
| Группы ксерофитов | Показания ФЭКа | Среднее | По шкале | Количество соединений (мг/кг) |
| контроль | 0 | 0 | 0 | 0 |
| 1группа | 0,37 - 0,39 | 0,38 | 0,058 | 117 |
| 2группа | 0,08 - 0,08 | 0,08 | 0,1232 | 246,4 |
| 3группа | 0,19 - 0,20 | 0,195 | 0,03 | 260 |
| 4группа | 1,0 - 1,0 | 1 | 0,1539 | 307,8 |
| Таблица 13. Содержание фенолов в почвенном опаде исследуемых групп ксерофитов в осенний период. | ||||
| Группы ксерофитов | Показания ФЭКа | Среднее | По шкале | Количество соединений (мг/кг) |
| контроль | 0 | 0 | 0 | 0 |
| 1группа | 0,21 - 0,21 | 0,21 | 0,0323 | 120,71 |
| 2группа | 0,1 - 0,08 | 0,09 | 0,014 | 289,7 |
| 3группа | 0,20 - 0,20 | 0,2 | 0,03 | 326,6 |
| 4группа | 0,25 - 0,23 | 0,2 | 0,0369 | 399,6 |
В исследуемых группах ксерофитов в весенний период содержание фенолов было следующим: 1 группа - 136, 66 мг, 2 группа - 176 мг, 3 группа - 179 мг, 4 группа - 240 мг на 100 г почвы. В осенний период также проводились аналогичные исследования и получены следующие результаты соответственно от 120,7 мг до 399, 6 мг.
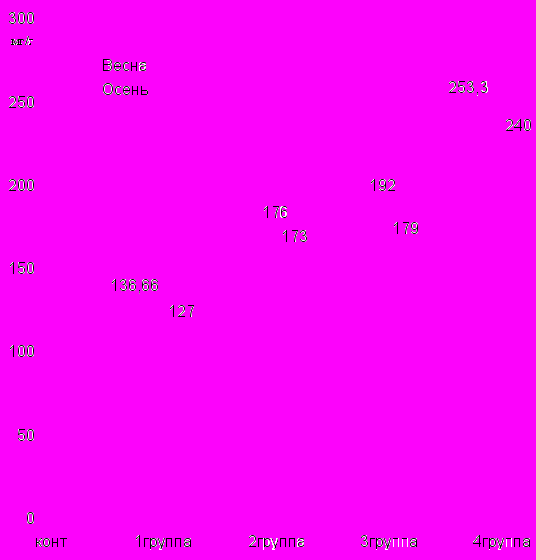
Диаграмма 10 Содержание фенолов в почвенном опаде исследуемых групп ксерофитов в весенний и осенний периоды.
| Таблица 14. Содержание фенолов в почвенном опаде исследуемых групп ксерофитов в весенний период. | ||||
| Группы ксерофитов | Показания ФЭКа | Среднее | По шкале | Количество соединений (мг/кг) |
| контроль | 0 | 0 | 0 | 0 |
| 1 группа | 0,115 - 0,120 | 0,12 | 0,02 | 136,66 |
| 2 группа | 0,09 - 0,8 | 0,09 | 0,01 | 176 |
| 3 группа | 0,26 - 0,29 | 0,27 | 0,04 | 179 |
| 4 группа | 0,03 - 0,04 | 0,35 | 0,05 | 240 |
| Таблица 15. Содержание фенолов в почвенном опаде исследуемых групп ксерофитов в осенний период. | ||||
| Группы ксерофитов | Показания ФЭКа | Среднее | По шкале | Количество соединений (мг/кг) |
| контроль | 0 | 0 | 0 | 0 |
| 1 группа | 0,113 - 0,112 | 0,01 | 0,02 | 127,6 |
| 2 группа | 0,095 - 0,07 | 0,08 | 0,01 | 173,3 |
| 3 группа | 0,52 - 0,46 | 0,49 | 0,07 | 192 |
| 4 группа | 0,40 - 0,56 | 0,48 | 0,07 | 253,33 |
В результате анализа полученных данных, мы пришли к выводу, что исследуемые физиологически активные вещества, относятся к группе фенольных соединений флавоноидов - кверцетина и мирицетина. Наши данные совпадают с результатами других исследователей (Э.Т.Оганесян, 1968). Кверцетин и мирицетин относятся к многочисленной группе флавоноидов содержащихся в высших растениях (М.Н.Запрометов1980). Было установлено, что в почве исследуемых групп ксерофитов содержатся фенолы. Их количества и сезонное увеличение имеют тенденцию к накоплению. В результате физиологически активные вещества содержатся в почвенном опаде ксерофитов на всех исследуемых стандартных площадках, что подтверждается химическими анализами, результатами биологической активности и визуально (усиленной интенсивностью окраски раствора).
Нами была проведено моделирование влияния флавоноидов - кверцетина и мирицетина, содержащихся в исследуемых группах ксерофитов на рост корней растений - биотестов. С этой целью нами были взяты химически чистые вещества и с помощью SPECOL подобраны три группы растворов различной концентрации для кверцетина и мирицетина. В этих трех группах растворов в трехкратной повторности, в течение четырех суток методом (таблица 16).
Таблица 16. Данные по моделированию среднего роста корней биотестов (огурец Мироновский-1) с использованием раствора кверцетина.
| Средняя длинна биотестов | Вода 0 | Раствор 0,1 | Раствор 0,01 | Раствор 0,001 |
| L 1 сут. мм | 28 | 30.4 | 38.1 | 42.5 |
| L 2 сут. мм | 69.3 | 70 | 82.5 | 81.1 |
| d L 2 мм | 41.3 | 39.6 | 44.4 | 38.6 |
| Прирост % 2 | 100 | 101 | 119 | 117 |
| L 3 сут. мм | 119.5 | 118 | 156 | 121 |
| d L 3 мм | 50.2 | 48 | 73.5 | 39.9 |
| Прирост % 3 | 100 | 95 | 130 | 101 |
| L б.к. 3 мм | 14.8 | 15.7 | 24.5 | 23.8 |
| L 4 сут. мм | 177.6 | 156.6 | 213 | 178.2 |
| d L 4 мм | 58.1 | 38.6 | 57 | 57.2 |
| L б.к. 4 мм | 31.9 | 34.4 | 48 | 37.5 |
| Прирост % 4 | 100 | 88 | 119 | 100 |
| DL мм | 149.6 | 126.2 | 174.9 | 135.7 |
| б.к. % | 100 | 107 | 150 | 117 |
Примечание: L - длина корней в мм, dL- прирост корней в мм, L б.к. - длина боковых коней мм, DL - суммарный прирост мм, б.к % - суммарный прирост боковых корней в %.
Было установлено, что раствор кверцетина (концентрация 0,1) является ингибитором и замедляет рост корней растений - биотестов. Раствор (концентрация 0,01) стимулирует развитие корневой системы проростков. Раствор (концентрация 0,001) также незначительно стимулировал рост корней за четверо суток. Развитие боковых корней растений- биотестов в растворе кверцетина также характеризовалось незначительным стимулированием. Вероятно 0,01 - это та концентрация при которой кверцетин является физиологически активным веществом и присутствует в листьях, листовом и почвенном опаде исследуемых групп ксерофитов и соответствует естественной концентрации. В исследуемых растворах мирицетина использовались концентрации 0,1; 0,01; 0,001. Нами проводилось аналогичное исследование с использованием того же растения - биотеста огурец сорта “ Мироновский 1 “. Было установлено, что наиболее стимулирующим рост и развитие корней растений - биотестов оказался раствор мирицетина концентрации 0,001. Прирост корней в растворе мирицетина в течение четырех суток был равен 58,1 мм или 142 % (таблица 17)
Интересно отметить, что развитие боковых корней в растворе мирицетина в течение четырех суток было иным, чем в растворе кверцетина той же концентрации.
Таблица 17. Данные по моделированию среднего роста корней биотестов
(огурец Мироновский-1) с использованием раствора мирицетина.
| Средняя длинна | Вода 0 | Раствор 0,1 | Раствор 0,01 | Раствор 0,001 |
| L 1 сут. мм | 28 | 45 | 55,7 | 55,3 |
| L 2 сут. мм | 69,3 | 91,2 | 106 | 123,8 |
| d L 2 мм | 41,3 | 46,2 | 50,3 | 68,5 |
| % 2 | 100 | 131 | 152 | 178 |
| L 3 сут. мм | 119,5 | 158,7 | 171,3 | 186,6 |
| d L 3 мм | 50,2 | 67,5 | 65,3 | 62,8 |
| % 3 | 14,8 | 30,8 | 34,2 | 44,5 |
| L б.к. 3 мм | 100 | 132 | 143 | 156 |
| L 4 сут. мм | 177,6 | 203,1 | 189,8 | 252,2 |
| d L 4 мм | 58,1 | 44,4 | 18,5 | 65,6 |
| L б.к. 4 мм | 31,9 | 51 | 56,5 | 73,5 |
| % 4 | 100 | 114 | 106 | 142 |
| dL мм | 149,6 | 158,1 | 134,1 | 196,9 |
| б.к. % | 100 | 159 | 177 | 230 |
Примечание: L - длина корней в мм, dL- прирост корней в мм, L б.к. - длина боковых коней мм, DL - суммарный прирост мм, б.к % - суммарный прирост боковых корней в %.
Таким образом, можно предположить, что кверцетин и мирицетин, содержащиеся в исследованных группах ксерофитов являются биологически активными соединениями. Созданная нами модель действия кверцетина и мирицетина показывает, что при малых концентрациях эти вещества стимулируют рост и развитие растений - биотестов, а при более высоких замедляют или ингибируют их. Действие кверцетина и мирицетина на разные части корневой системы различны, но общее воздействие их аналогично действию физиологически активных соединений, образующихся в исследуемых группах ксерофитов. Можно предположить, что поступление в почву столь больших количеств фенольных веществ и их аккумуляция в корнеобитаемой среде имеют серьезные экологические последствия и существенно влияют на рост и развитие ксерофитов.
Изложенные выше данные и теоретические положения позволяют заметить, что длительная адаптация к разнообразным экстремальным условиям среды привела к такой физиолого-структурной и биохимической перестройке в различных исследуемых нами группах ксерофитов, которая способствовала повышению уровня его гомеостаза для объяснения нормального хода процессов жизнедеятельности и устойчивости к указанным неблагоприятным условиям. При произрастании на определенном месте одних растений, некоторые из подвижных экзометаболитов могут накапливаться в почве. Их присутствие отражается на росте и развитии растений, образующих с ними фитоценозы.
При определении суммы фенолов в почве мы изучали исследуемые группы в весенний и осенний период (диаграмма 11).
Степень воздействия отдельных экологических факторов на биосинтез фенолов окончательно не выяснена, но, вероятно, основным фактором является свет. В процессе подготовки растения к листопаду (осенью) происходит более полный отток органических веществ в том числе и фенольных в почву, которые за зимнe - весенний период вместе с талыми водами смываются. Под влиянием многих факторов концентрация фенольных соединений в исследуемых группа растений ксерофитов различна. Одной из таких причин является стресс, испытываемый растениями, в условиях аридного климата. Можно предположить, что стимулирование синтеза фенольных соединений у ксерофитных видов может воздействовать на способность растений выживать в этих условиях в результате метаболических изменений. Высокие концентрации фенольных соединений в условиях засушливого климата на наш взгляд являются своего рода адаптацией в суровых окружающих условиях. При таких экстремальных условиях значительная часть доступной энергии частично расходуется на поддержание жизненно важных функций растения.

Диаграмма 11 Определение оптической плотности вытяжки разных групп ксерофитов в весенний и осенний периоды.
При этих условиях более консервативные индивидуумы, в частности суккуленты такие как -Sempervivum caucasicum - Молодило кавказское, S. oppositifolium - О. супротиволистный, S. subulatum - О. шиловидный, и схожей по своим физиологическим свойствам группой ксерофитов Euphorbia glareosa - Молочай хрящеватый, E. stepposa - М. степной, E. procera - М. мохнатый, E. petrophila - М. скалолюбивый, E. boissieriana Woronow - М. Буасье и настоящие ксерофиты Ковыль волосатик, Марь олиственная могут выживать и репродуцировать, доминировать, образуя собственные фитоценозы. Следовательно, способность к самоингибированию и ингибированию окружающих растений при помощи фенольных соединений является гибким механизмом адаптации.