
Е. П. Лаврова Научный руководитель Чапоров В. Н
| Вид материала | Документы |
- Информационно-поисковая система “Научный потенциал вуза”, 31.25kb.
- Магнитоакустическая эмиссия магнетитовых и титаномагнетитовых руд железорудных месторождений, 290.93kb.
- Председателем Оргкомитета конференции является научный руководитель ниу вшэ профессор, 51.47kb.
- Председателем Оргкомитета конференции является научный руководитель ниу вшэ профессор, 56.04kb.
- Научный совет по философии образования и проблемам методологии исследования в образовании, 166.51kb.
- Кирилла Юрьевича Лаврова 2- 11 апреля 2008 года Пресс-релиз программа, 150.93kb.
- Программа программный комитет исаев Александр Сергеевич, академик ран, д б. н., научный, 427.64kb.
- Диакона Александра (Урбановича) (научный руководитель иерей Александр Тимофеев), затрагивающая, 1909.34kb.
- Общий менеджмент Батяшев В. И. Повышение эффективности производства замороженных полуфабрикатов, 983.44kb.
- Магистерская программа «маркетинг-менеджмент» (научный руководитель – доцент,, 16.48kb.
Amanita fulva W.G. Smith., A. muscaria (L.) Pers., Anthracobia macrocystis (Cooke) Boud., Artomyces pyxidatus (Pers.: Fr.) Jülich.
(= Clavicorona pyxidata), Calvatia utriformis (Bull.: Pers.) Jaap.(= C. caelata, Handkea utriformis), Cantharellus cibarius Fr.: Fr., Clavariadelphus
ligula (Schaeff.: Fr.) Donk., Cortinarius armillatus (Fr.: Fr.) Fr.,
C. sanguineus (Wulfen: Fr.) Fr., Collybia confluens (Pers.: Fr.) P. Kumm., Coltricia perennis (L.: Fr.) Murrrill, Coprinus micaceus (Bull.: Fr.) Fr., Crucibulum leave (Huds.) Kambly, Fomitopsis pinicola (Sw.: Fr.) P. Karst., Hygrocybe conica (Scop.: Fr.) P. Kumm., Hypholoma fasciculare (Huds.: Fr.) P. Kumm., H. sublateritium (Fr.) Quèl., Hypoxylon multiforme (Fr.) Fr., Kuehneromyces mutabilis (Schaeff.: Fr.) A. H. Sm., Leccinum scabrum (Bull.: Fr.) Gray, L. varicolor Watling, Lepiota cristata (Bolton: Fr.) P. Kumm., Lycoperdon perlatum Pers : Pers., L. pyriforme Schaeff.: Pers., Macrolepiota
procera (Scop.: Fr.) Marasmius scorodonius (Fr.: Fr.) Fr., Singer, Mycena pura (Pers.: Fr.) P. Kumm., M. rosella (Fr.) P. Kumm., Otidia
leporina (Batsch) Fuckel., Paxillus involutus (Batsch: Fr.) Fr., Peziza
badia Pers.: Fr., Pholiota flammans (Fr.) P. Kumm., Piptoporus
betulinus (Bull.: Fr.) P. Karst., Ramaria eumorpha (P. Karst.) Corner
(= R. invalii), R. stricta (Pers.: Fr.) Quèl., Rutstroemia luteovirescens (Roberge) White, Stropharia aeruginosa (M.A. Curtis: Fr.) Quèl., Trichia varia (Pers.) Pers., Xerocomus subtomentosus (L.: Fr.) Quèl.
Найдены новые местонахождения вида, занесенного в Красную книгу Тверской обл. (2002) (Geastrum quadrifidum Pers.: Pers.). Отмечено неизвестное ранее местообитание редкого для области вида ( Micromphale perforans (Hoffm.: Fr.) Singer (= Marasmius perforans (Hoffm. : Fr.) Fr.), который был найден в 1901 г. В.Г. Траншелем (1901).
CПИСОК ЛИТЕРАТУРЫ
1. Курочкин С.А., Медведев А.Г. Материалы к флоре Тверской области.
Ч. 3: Грибы. Тверь, 1998.
2. Красная книга Тверской области. Тверь, 2002.
3. Траншель В.Г. Список грибов, собранных в Валдайском уезде Новгородской губернии // Тр. пресновод. биол. ст. Санкт-Петерб. о-ва естествоиспыт. 1901. Т.1. С. 160 – 203.
Секция зоологии
В.В. ЕГОРОВ
Научный руководитель – Зиновьев А.В.
АБСОЛЮТНАЯ ЧИСЛЕННОСТЬ И ПЛОТНОСТЬ ГНЕЗДОВАНИЯ ХИЩНЫХ ПТИЦ СЕВЕРО-ЗАПАДА ФИРОВСКОГО РАЙОНА ТВЕРСКОЙ ОБЛАСТИ
Без знания количества обитающих на определенной территории хищных птиц невозможно осуществление долгосрочных практических мер по их охране. Результирующая экологических связей птиц с окружающей средой – численность. Ее долгосрочные изменения, тенденция движения – наиболее четкий количественный критерий реакции на состояние среды (Галушин, 1974).
Исследования проводились в период с апреля по август 2005-2008гг. на территории Тверьохотобъединения «Егерь». Площадь исследуемого стационара 250 км2. Стационарные методики исследования: визуальные наблюдения с высоких точек, наблюдения на акваториях крупных озер (Новиков, 1949) сочетались с маршрутными учетами (Осмоловская, Формозов, 1952).
Абсолютная численность и плотность гнездования
За период работы на стационаре на площади 250 км2 нами установлено гнездование 37 пар дневных хищных птиц, принадлежащих к 11 видам. Абсолютная численность хищных птиц на стационаре соответствует в среднем за три года 31 паре. Суммарная плотность населения равна 14,8 пар на 100 км2 общей площади (таблицы 1, 2). Для лесопокрытой площади этот показатель возрастает до 31,5 пар на 100 км2. Доля участия разных видов в суммарной численности не одинакова. Абсолютно доминируют два вида: канюк (27%) и перепелятник (27%). Им значительно уступает скопа (11%). Эти три вида вместе составляют 55% всех хищных птиц стационара. Из остальных 8 видов, на которые в сумме приходится 45%, малочисленными следует считать беркута, дербника и орлана-белохвоста. На их долю приходится по 2,7%. Низкая численность осоеда (2,7%), по-видимому, объясняется возможным недоучетом из-за скрытого образа жизни и охоты в лесных угодьях. Редки в условиях стационара также болотный лунь (5,41%) и черный коршун (5,41%).
По сравнению с 1989 г. плотность гнездования большинства хищных птиц в значительной степени изменилась у 8 видов хищников: канюк, тетеревятник, перепелятник, скопа, чеглок, орлан-белохвост, осоед, болотный лунь.
Снижение плотности таких хищников как канюк, тетеревятник и осоед, вероятно, связано с пространственным перераспределением гнездовий дневных хищных птиц, вследствие снижения антропогенного пресса на территорию стационара и сопредельных территорий и как следствие перераспределения кормовой базы и увеличения числа гнездопригодных территорий вне стационара. Таким образом, уменьшение плотности произошло за счет распределения прежнего числа гнездовых пар на больших по сравнению с 1980 г. территориях.
Снижение численности орлана-белохвоста, по-видимому, связано с сокращением кормовой базы озера Серемо. В середине 1990 г. произошел сильный замор рыбы. Вероятно, восстановление кормовой базы приведет к восстановлению численности орлана.
Таблица 1
Численность и плотность гнездования дневных хищных птиц на стационаре 2008 г. в сравнении с данными 1989 г.
| Вид | Число пар на стационаре | Число пар на 100 км2 общей площади | Число пар на 100 км2 леса | |||
| 2008 | 1989 | 2008 | 1989 | 2008 | 1989 | |
| Канюк | 10 | 17 | 4,0 | 7,0 | 8,6 | 14,7 |
| Перепелятник | 10 | 5 | 4,0 | 2,0 | 8,6 | 4,3 |
| Скопа | 4 | 2 | 1,6 | 0,8 | 3,6 | 1,8 |
| Чеглок | 3 | 1 | 1,2 | 0,4 | 2,7 | 0,9 |
| Тетеревятник | 2 | 7 | 0,8 | 2,8 | 1,72 | 6,0 |
| Черный коршун | 2 | 2 | 0,8 | 0,8 | 1,8 | 1,8 |
| Болотный лунь | 2 | 1 | 0,8 | 0,4 | – | – |
| Орлан-белохвост | 1 | 2 | 0,4 | 0,8 | 0,9 | 1,8 |
| Беркут | 1 | 1 | 0,4 | 0,4 | 0,9 | 0,9 |
| Дербник | 1 | 1 | 0,4 | 0,4 | 0,9 | 0,9 |
| Осоед | 1 | 2 | 0,4 | 0,8 | 0,9 | 1,8 |
| Итого | 37 | 41 | 14,8 | 16,4 | 31,5 | 35,6 |
Рост численности чеглока и болотного луня, по всей видимости, также связан со снижением антропогенного пресса на территорию стационара. Однако, в отличие от канюка, тетеревятника и осоеда, чеглок и болотный лунь на территории стационара имеют весьма ограниченные возможности для гнездования, то есть увеличение плотности произошло за счет роста численности на прежних гнездовых территориях.
Рост численности скопы, по-видимому, связан с улучшением кормовой базы на озере Шлино.
Таблица 2
Абсолютная численность гнездовых пар хищных птиц на территории стационара в период исследований в сравнении с данными 1986 г.
| Вид | Число гнездовых пар | |
| в среднем за 1984 – 1987 гг. | в среднем за 2005 – 2008 гг. | |
| Канюк | 17 | 9 |
| Перепелятник | 5 | 10 |
| Скопа | 2 | 3 |
| Чеглок | 1 | 2 |
| Тетеревятник | 7 | 1 |
| Черный коршун | 1 | 1 |
| Болотный лунь | 1 | 1 |
| Орлан-белохвост | 2 | 1 |
| Беркут | 1 | 1 |
| Дербник | 1 | 1 |
| Осоед | 1 | 1 |
| Итого: | 44 | 31 |
Сравнительное богатство авифауны северо-запада Фировского района объясняется водораздельным положением территории стационара и редким для Тверской области сочетанием ландшафта верховых болот и котлованных озер с лесопольным ландшафтом смешанных лесов. Наряду с этим, снижение антропогенного пресса приводит к постепенному восстановлению кормовой базы, что является определяющим в данном случае фактором стабилизации численности хищных птиц.
СПИСОК ЛИТЕРАТУРЫ
- Галушин В.М. Хищные птицы и современная среда: конспект проблемы // Материалы VI Всесоюзной орнитологической конференции. Ч.1. М., 1974. С. 42 – 45.
- Новиков Г.А. Полевые исследования экологии наземных позвоночных животных. М., 1949.
- Осмоловская В.И., Формозов А.Н. Методы учета численности и географического распространения дневных и ночных хищных птиц. // Методы учета численности географического распространения наземных позвоночных. М., 1952.
- Беляков В.В. О численности и распределении дневных хищных птиц на территории Калининской области. // Ученые записки Калининского государственного пединститута. Калинин, 1964, С. 20 – 40.
- Красная книга Тверской области / Ред. А.С. Сорокин. Тверь, 2002.
А. А. КРУЖКОВА
Научный руководитель – Самков М.Н.
ИСПОЛЬЗОВАНИЕ СВЕТО-АРОМОЛОВУШКИ
ДЛЯ СБОРА НОЧНЫХ ЧЕШУЕКРЫЛЫХ
В истории науки постоянно происходили усовершенствования методик и средств исследования окружающего мира, часто обусловленные техническим прогрессом. То же самое наблюдалось и в эволюции светоловушек. Так электролампы сменили пламя свечи, но они привлекают насекомых лишь одним своим свойством – светом. На данном этапе развития науки, уровне знаний о биологии ночных чешуекрылых и других фотоксенов – этого уже недостаточно. Как показывает практический опыт, комбинированные методики дают большую эффективность. Поэтому было решено использовать еще один привлекающий фактор – запах брожения.
Свето-аромоловушка представляет собой модификацию стандартной Пенсильванской светоловушки (Горностаев, 1984; Ламперт, 2003), т.е. лопасти (рисунок, 5) сделаны из двух пластин прозрачного оргстекла, в верхней их части имеется прорезь, позволяющая свободно помещаться энергосберегающей лампе (Навигатор, NCL-2U, E27, 15 Вт, 2700K) теплого свечения (рисунок, 4).
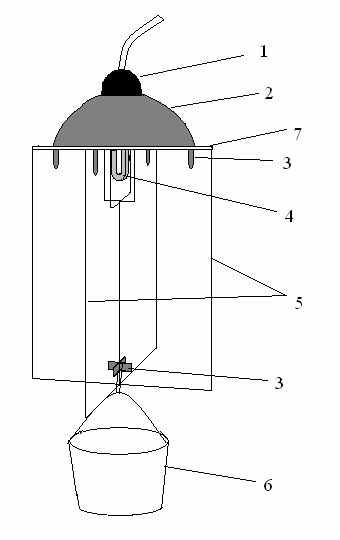
Рисунок. Структура свето-аромоловушки:
1 – патрон лампы, 2 – металлический колпак, 3 – металлические уголки,
4 – лампа, 5 – лопасти из оргстекла, 6 – емкость,
7 – поперечная пластина оргстекла
Сверху лампочка прикрыта металлическим колпаком (рисунок, 2), в который вставлен патрон (рисунок, 1). К нижней части крестовины крепится емкость (рисунок, 6) со слаборазбавленным пивом. Раствор периодически, по мере необходимости, заменялся свежим. Такая ловушка была установлена на балконе третьего этажа в микрорайоне Мамулино.
Использовалась она с 6 июня по 3 сентября 2007 г, т.е. 90 дней. Данным способом было поймано 69 видов совок (Noctuidae) из 98 собранных аромоловушкой, сачком и свето-аромоловушкой. Из них 30 видов было поймано только данной ловушкой. Общее количество особей составило 185. Среди данных видов обнаружено 11 редких: Catocala fraxini L., Mythimna imbecilla F., Heliophobus reticulata G., Noctua umbrosa Hbn., Paradiarsia mista Frr., Amphipoea lucens Frr., Amphipoea oculea L., Celaena haworthii Curt., Hydroecia petasitis Doub., Photedes fluxa Hbn., Hoplodrina lenta Tr., (Мержеевская, 1971; Ламперт, 2003; Бабочки мира, 2007).
В 2008 г с 27 мая по 9 сентября (106 дней) было поймано 47 видов из 55 зафиксированных с помощью аромоловушки, сачка и свето-аромоловушки. Общее количество особей составило 156. В это число входит 1 исчезающий вид – Polia bombycina Hufn., и 4 редких: Noctua umbrosa Hbn., Amphipoea oculea L., Caradrina selini Bsd., Hydroecia petasitis Doub. (Мержеевская, 1971; Ламперт, 2003; Бабочки мира, 2007).
СПИСОК ЛИТЕРАТУРЫ
1. Горностаев Г.Н. Лет насекомых на искусственные источники света. Л., Наука, 1984. Т.66. С. 101 – 151.
2. Ламперт К. Атлас бабочек и гусениц. Места обитания. Особенности строения. Поведение. Размножение. Техника ловли. Коллекционирование. / К. Ламперт. Мн.; Харвест, 2003.
3. Мержеевская О.И. Совки (Noctuidae) Белоруссии // Наука и техника. Мн., 1971.
4. Бабочки мира: [электрон. ресурс]. 2007. 1 DVD-диск. 2,47 Гб. (Медиаэнциклопедия DVD).
О.А. ЕГОРОВА
Научный руководитель – Емельянова А.А.
НЕКОТОРЫЕ РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ ХРОМОСОМНЫХ РАС
ОБЫКНОВЕННОЙ БУРОЗУБКИ (SOREX ARANEUS L.)
НА ТЕРРИТОРИИ ВАЛДАЙСКОЙ ВОЗВЫШЕННОСТИ
В 2007 г. на территории Валдайской возвышенности обнаружена граница контакта трех хромосомных форм бурозубки обыкновенной: «Селигер», «Западная Двина» и «Москва». Теоретически существование областей контакта трех хромосомных рас было очевидно, но ранее локализовать их не удавалось. Данные расы отличаются друг от друга по сочетанию вариантов диагностических хромосом: раса Москва – (gm, hi, kr, no, pq), раса Западная Двина- (gm, hk, ip, no, qr) и раса Селигер (g, hn, ik, m/q, o, pr). Форма «Селигер» имеет небольшой ареал западнее и южнее оз. Селигер, в поперечнике не более 100 км. Южнее, в верховьях Западной Двины и Ловати распространена форма «Западная Двина», а восточнее – форма «Москва», имеющая обширный ареал в междуречье Верхней Волги и Оки. При этом формы «Селигер» и «Западная Двина» распространены на территории последней (Поздневалдайской) ледниковой области, а форма «Москва» – во внеледникой области и зона контакта между ними проходит по границе Вепсовской ледниковой стадии (16 – 15 тыс. лет назад). Детальный анализ распространения трех хромосомных рас в области контакта их ареалов весьма перспективен, т.к. позволяет решить ряд вопросов эволюционной и популяционной биологии, таких, как реконструкция процессов формирования ареала современного вида, изучение особенностей популяционно-генетической структуры вида, а также морфологии и экологии внутривидовых генетических форм и мн. др.
Сбор материала проводился в составе экспедиции ИПЭЭ РАН в июне-июле 2007 – 2008 гг. на территории Валдайской возвышенности. Зверьки отлавливались специальными живоловками с проволочным трапиком конструкции Н.А. Щипанова. Хромосомные препараты готовили стандартным методом из клеток красного костного мозга и селезенки. Хромосомная раса бурозубки определялась по комбинациям диагностических хромосом в кариотипе (g h i k m n o p q r), в соответствии с международной номенклатурой хромосом этого вида.
В ходе работ было обнаружено, что обширный полигон хромосомной изменчивости рас в районе озера Любино (Андреапольский р-н, Тверская обл.) характеризуется наличием множественных зон контакта двух из трех указанных рас. Отмечено 11 таких локалитетов совместного обитания рас Западная Двина, Москва и Селигер в различных сочетаниях. На юго-восточном берегу озера Любино, на небольшой территории 1000мх1000м, зафиксирована область соприкосновения ареалов всех трех рас: Западная Двина, Москва и Селигер (см. рисунок). Вероятно, между ними существует очень ограниченный поток генов, о чем свидетельствует парапатрическое распространение рас с редкой гибридизацией и узкими напряженными гибридными зонами. Встреча гибридных особей в исследованной зоне не превысила 20 %.
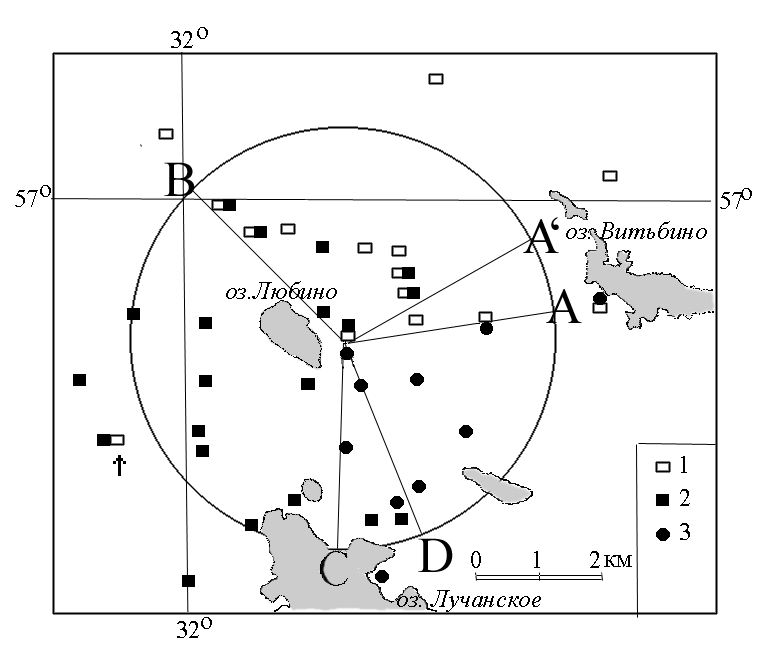
Рисунок. Места находок хромосомных рас обыкновенной бурозубки в области контакта ареалов трех хромосомных рас:
Wd, Mo – ближайшие находки рас Западная Двина и Москва, соответственно, в пределах распространения расы Селигер;
1 – раса Селигер, 2 – раса Западная Двина, 3 – раса Москва
А.О. ЗВЕЗДИН
Научный руководитель – Костин В.В.
МЕТОДИКА ИЗМЕРЕНИЯ КРИТИЧЕСКИХ СКОРОСТЕЙ
ТЕЧЕНИЯ ДЛЯ МОЛОДИ РЫБ
С ПОМОЩЬЮ РАСХОДНОГО СПОСОБА
Критическая скорость течения – это верхняя граница того интервала скоростей, в пределах которого возможно удержание рыб в потоке. Её величина равна минимальной скорости потока, который сносит рыб (Павлов, 1979). Удержание в потоке происходит посредством реореакции. Реореакция – врожденная реакция рыб направленная на сохранение места обитания в потоке и выражающаяся в активном движении рыб против течения.
Факторов влияющих на величину критической скорости множество: видовая принадлежность рыб, их размеры и форма тела, температура воды, физиологическое состояние рыб, степень накормленности, уровень мотивации рыб и др. Также к этим факторам относятся гидродинамические условия водоема. Известно, что рыбы одного вида, взятые из водоемов с разными гидродинамическими условиями, различаются по показателю критической скорости течения (Павлов, 1979).
Настоящее исследование, направленное на изучение реореакции рыб, проведено в два периода: с 19,06,08 по 22,06,08 и с 24,06,08 по 4,07,08. В течение каждого периода проводились 2 серии опытов с рыбами из двух водоёмов: из реки Суноги и из реки Ильд (Некоузский район, Ярославская область). Указанные реки различались по своим гидродинамическим характеристикам. Река Сунога – небольшие заводи на участке где проводился отлов рыб, в зоне подпора Рыбинского водохранилища, стоковое течение не выражено, ветровые течения различного направления. Река Ильд – в районе деревни Верхнее Никульское, постоянное, быстрое течение.
Объектами исследования были личинки плотвы ( Rutilus rutilus L.) и леща (Abramis brama L.) на этапах развития D2 и E (Коблицкая, 1981). Длина личинок плотвы составляла 11 – 18,5 мм; леща – 11.9 – 15.9 мм.
Определение критических скоростей проводили как необходимый первый этап исследования параметров реореакции рыб – для настройки скорости потока воды в рабочих установках по определению типа и параметров реореакции.
Установка для определения критических скоростей представляла собой стеклянную трубку, в которой создавали регулируемый по скорости ток воды. Ток воды обеспечивали водяным насосом. Концы трубки закрывали заградительными сетками. Трубка была зафиксирована на горизонтальной поверхности с нанесенными на ней зрительными ориентирами – черные полоски шириной в 1 см и интервалом 5 см. Помещенную внутрь личинку рыбы выдерживали около одной минуты без течения и столько же со слабым течением для адаптации к условиям эксперимента. Затем скорость потока воды быстро и равномерно увеличивали. В момент, когда рыба переходила к бросковому движению или сносилась потоком, её извлекали из установки, путем слива в пересадочную емкость током воды, и отправляли на камеральную обработку. В процессе камеральной обработки производили измерение длины каждой рыбы от конца рыла до конца хорды, определение вида и этапа развития по А.Ф.Коблицкой (1981). Зафиксированную скорость рассчитывали расходным методом. Диаметр трубки составлял 10 мм. При известных: диаметре трубки, из которой истекает вода и объеме воды, измеренном за 1 мин, рассчитывали абсолютную скорость воды и скорость воды относительно длины тела рыбы. У рыб данной серии критические скорости определяли в выборке из 30 особей. Результат выборки усредняли.
Всего в указанные сроки проведено 120 экспериментов, соответственно со 120 рыбами.
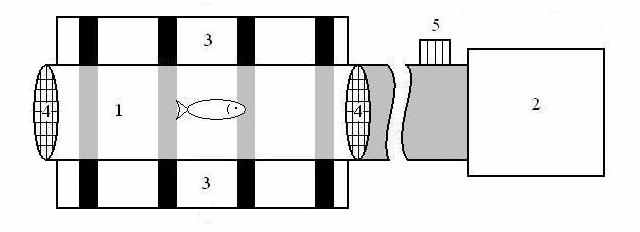
Рис.1. Схема установки для определения критических скоростей:
1 – стеклянная трубка, 2 – насос, 3 – зрительные ориентиры,
4 – заградительные решетки, 5 – регулировочный вентиль,
–
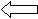 направление тока воды
направление тока водыРанее подобные эксперименты, как с молодью, так и с взрослыми рыбами проводились в гидродинамических лотках, скорость в которых измерялась при помощи небольшого винта. Скорость его вращения регистрировалась компьютеризированным комплексом. Подобный метод является одним из наиболее точных, но следует отметить, что на точность измерения влияет турбулентность потока воды, довольно высокая из-за ширины лотка (более 10 см). Небольшой, всего 10 мм, диаметр трубки, использованный в установке, сводит к минимуму влияние турбулентности. Основная погрешность остается в точности замера объема вытекаемой воды. Таким образом, описанный выше метод измерения критических скоростей является наиболее точным при работе с личинками рыб небольших размеров.
СПИСОК ЛИТЕРАТУРЫ
1. Коблицкая А.Ф. Определитель молоди пресноводных рыб. М., 1981.
2. Павлов Д.С. Биологические основы управления поведением рыб в потоке воды. М., 1979.
А.С. ВОЛКОВА
Научный руководитель – Чернецов Е.А.
МЕТОДИКА ИССЛЕДОВАНИЯ ЛОКОМОЦИИ ЗМЕЙ
Для изучения способов движения змей используется фотографии, видеонаблюдения и описание следов на земле, но практически не уделяется внимание на работу мышц, брюшных щитков и особенности скелета. Учитывая это, следует разрабатывать новые методики исследования локомоции змей. Ниже описаны приёмы и приборы, разработанные для решения поставленной задачи.
Змеи при движении по поверхности земли используют четыре типа передвижения: 1) латеральный волнообразный, или просто «змеевидный», которым пользуется большинство видов змей; 2) прямой или «гусеничный» ход, используют который тяжёлые змеи (например, Boa restricter); 3) боковой ход используется на песках (Epha sp.); 4) подтягивание или смешанный тип (Брем, 2000; Волкова, 2007).
Для испытания данных методик и приборов был использован полоз (Elaphe sp.). Это некрупная змея (длинна 1,1 – 1,2 м и обхват 2 – 2,5 см), цвет коричнево-серый или коричневый с чёрными продольными полосами. Голова плавно переходит в шею, цвет оливковый. Брюшные щитки крупные коричнево-розового цвета. Тип локомоции «змеевидный», скорость полоза 5 км/час. Место обитания пустыни и полупустыни.
Для определения типа локомоции, которые использует змея в узком пространстве были взяты несколько труб различной длинны и диаметра. При прохождении их полозом, описывался способ движения, можно произвести расчет скорости змеи при данном типе локомоции и в заданном пространстве. Например, в трубе с диаметром 12 – 15 см змея двигалась
S-образно и могла свернуться клубком, во – второй трубе (диаметр
5 – 7 см) использовался тот же тип, но амплитуда изгибов была меньше. В трубе с диаметром 3 – 4 см изгибы почти не наблюдались, полоз использовал «гусеничный» ход.
Для сбора данных (частота колебаний щитков, измерение скорости движения змеи) были разработаны следующие устройства.
Прибор № 1 (рис. 1) представляет собой платформу с выпуклым желобом. Платформа или основание – длинна 1,2 м; ширина 8 см; три отверстия под микрофоны или какие-либо датчики. Длинна между
1 и 2 микрофоном 35 см; между 2 и 3 – 38 см. Жёлоб прозрачный, состоящий из двух листов пластика, низ выстлан мешковиной. От микрофонов идут три выхода для подключения к различным устройствам (диктофон, осциллограф, ноутбук, ПК и другая аудио – и видеоаппаратура). В целях снижения стресса и нагрузок у змеи жёлоб покрывается шторкой с липучками.
Прибор № 2 (рис. 2) – платформа различного размера (в данном случае 25 см на 40 см, из ДСП) покрытая гладким материалом (стеклом или пластиком) и одним отверстием с подвешенным микрофоном. Гладкая поверхность устройства предназначено, для того чтобы свести к минимуму трение щитков о поверхность и силу толчка. Над отверстием закрепляется железная или картонная «уздечка». Она сделана, для того, чтобы змея проползала над микрофоном.
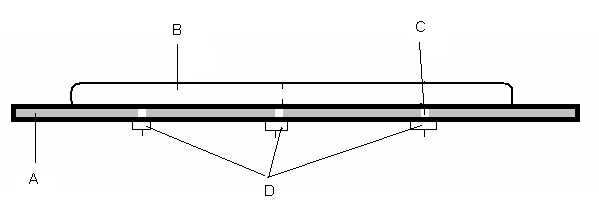
Рис. 1. Камера для фиксации параметров движения змей (по: Волкова, 2007)
A – платформа или основание; B – жёлоб; C – отверстие с микрофоном;
D – выходы на приборы записи данных
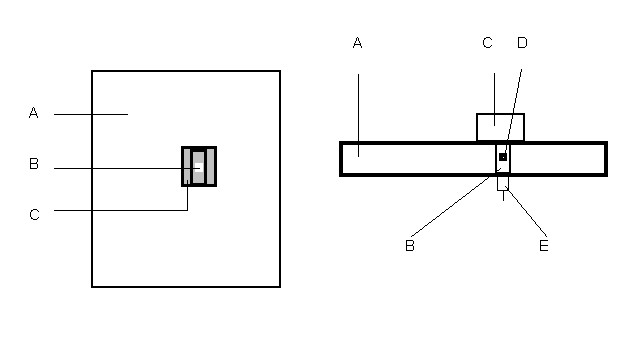
Рис. 2. Камера (дополнительные проекции)
A – платформа, покрытая гладким материалом; B – отверстие для микрофона;
C – «уздечка»; D – микрофон; E – выход на приборы записи данных
Анализ и обработка собранной информации, в данном случае аудиозаписей, производится на ПК и с помощью программы Cool Edit Pro.
СПИСОК ЛИТЕРАТУРЫ
1. Брэм А. Рептилии. М., 2000.
2. Волкова А.С. Камера для определения некоторых параметров движения змей // Материалы науч. Конф. Студентов и аспирантов, апр. 2007 г. Тверь, 2007. С. 75 – 76.