
Лекция Факторы эволюции Формы естественного отбора
| Вид материала | Лекция |
- Наследственность и ее роль в патологии медицинская генетика и ее задачи Наследственность, 327kb.
- Ч. Дарвин. Сочинения,, 7194.26kb.
- Конспект лекций Москва 2006 г. Лекция № Основные источники техногенного загрязнения, 1559.24kb.
- Рабочая программа предмет биология Класс 11 Образовательная область естествознание, 51.79kb.
- «Элементарные эволюционные факторы», 91.21kb.
- Основные этапы эволюции растительного и животного мира, 173.64kb.
- Альфред Рассел Уоллес Правда о вакцинации Краткое изложение, 211.73kb.
- Темы семинаров по теории эволюции (главы реферата) Изученность группы, предковые формы,, 14.45kb.
- Лекция №7 -2011 Тема. Гравитационные склоновые процессы, 53.04kb.
- Реферат по биологии эмбриональное развитие человека, 404.29kb.
Лекция 2. Факторы эволюции
Формы естественного отбора
Различают несколько форм естественного отбора: стабилизирующую, движущую, разрывающую и дестабилизирующую. Стабилизирующая форма действует в том случае, если условия среды остаются неизменными. В этом случае преимущественно выживают особи, приспособленные к данным условиям, отклонения от среднего значения признака устраняются отбором. Учение о стабилизирующей форме отбора было разработано русским ученым И.И.Шмальгаузеном. Благодаря этой форме отбора сохраняются примитивные формы жизни, хорошо приспособленные к конкретным условиям; сохраняются средние значения различных признаков. Под контролем стабилизирующей формы отбора находятся самые различные признаки организма. Хорошим примером являются размеры ушей у зайцев (рис. 345), являющихся важным органом регуляции теплоотдачи. От их размеров зависит количество теплового излучения. Большее количество зайцев имеют уши средних размеров, короткоухие погибают от перегревания, длинноухие — от переохлаждения.
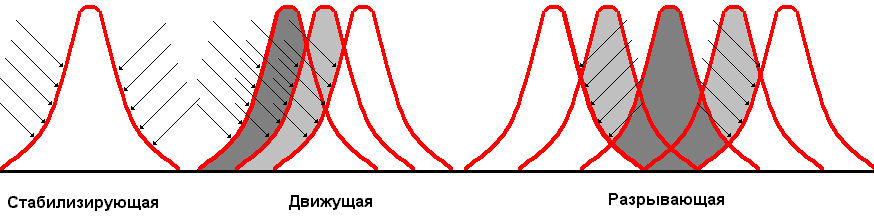
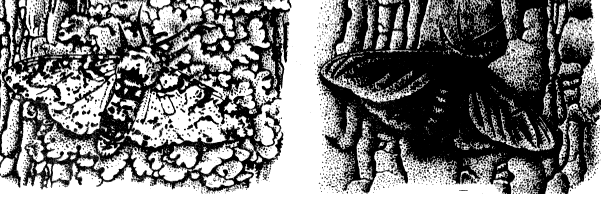
Рис. 345. Формы естественного отбора: стабилизирующая (размеры ушей у зайцев), движущая (окраска бабочки березовая пяденица), разрывающая (крылья насекомых океанических островов).

Классическим примером движущей формы естественного отбора является эксперимент с бабочкой березовая пяденица. Светлая форма мало заметна на деревьях вдали от города (на березах, покрытых лишайниками), где эта форма и преобладает. Около промышленных центров Англии, где проводился эксперимент, исчезли лишайники на березах и сами стволы стали более темными. Здесь преобладают темноокрашенные особи.
Разрывающую форму естественного отбора (дизруптивный отбор) Ч.Дарвин наблюдал на океанических островах, на которых появились бескрылые насекомые и насекомые с очень длинными крыльями. Насекомые со средними крыльями ветром сносились в море и погибали. Мадагаскарские тараканы, в отличие от Американских блаберусов, совершенно не имеют крыльев, но цепкие лапки позволяют им подниматься даже по стеклянной вертикальной плоскости. Разрывающая форма отбора приводит к полиморфизму — образованию нескольких форм, отличающихся по определенному признаку.
Дестабилизирующий отбор приводит к расширению нормы реакции признака, при этом преимущество получают особи не с узкой (как при стабилизирующем отборе), а широкой нормой реакции. Впервые эта форма отбора описана американским эволюционистом Д.Г.Симпсоном, термин дестабилизирующий отбор был предложен Н.Н.Воронцовым. Например, лягушки, живущие в водоемах с разными условиями освещенности и разнообразной растительностью имеют более разнообразную окраску, чем лягушки, живущие в водоемах с однородными условиями.
Таким образом, естественный отбор играет роль не только «сита», устраняющего появляющиеся в популяции изменения, но и играет творческую роль, накапливая наследственные изменения и определяя направление эволюции. Вместе с наследственной изменчивостью и другими факторами эволюции за 3,5 млрд. лет отбор создал все огромное разнообразие видов на нашей Земле, сохраняя как примитивные формы при стабильных условиях среды, и образуя новые виды в измененных условиях обитания.
Факторы эволюции
Кроме наследственной изменчивости и естественного отбора к факторам эволюции относят еще дрейф генов, популяционные волны и изоляцию.
Д

рейф генов как фактор эволюции был раскрыт российскими учеными генетиками Н.П.Дубининым и Д.Д.Ромашевым и зарубежными учеными американцем С.Райтом и англичанином Р.Фишером. Он касается малочисленных популяций с генофондом, ограниченным небольшим количеством генотипов в популяции. Генофонд таких популяций, в силу различных процессов, через несколько поколений может резко измениться, изменится частота встречаемости аллелей различных генов. В эксперименте С.Райт было задействовано100 линий, каждая из которых происходила от четырех самцов и четырех самок. Частота доминантного аллеля красных и рецессивного аллеля белых глаз в каждой линии исходно составляла 0,5 – т.е 50% мужских и женских гамет несли аллель красноглазости (XF), 50% несли аллель белоглазости (Хf).
Среди потомства в каждом поколении случайным образом отбирали четырех самцов и четырех самок для производства потомства. Через 16 поколений оба аллеля оказались в различных соотношениях: в 41 популяции был утрачен аллель Хf, все мухи были красноглазыми; в 29 популяциях был утрачен доминантный аллель ХF, все мухи были белоглазыми; 4 линии погибли, остальные сохранили оба аллеля, но в различных соотношениях.
Процесс случайного ненаправленного изменения частот аллелей в небольших популяциях получил название дрейфа генов, чем меньше численность, тем выше степень изменения частоты встречаемости аллелей генов.
В результате дрейфа генов происходит случайное изменение частот встречаемости различных аллелей, утрата части аллелей, что сказывается на дальнейшей эволюции вида.
В любой популяции происходят периодические колебания численности особей, причинами которых служат различные абиотические и биотические факторы среды. Наиболее ярко это проявляется у быстроразмножающихся видов, например у мышевидных грызунов примерно раз в 4 года численность возрастает многократно. Затем вновь происходит резкий спад численности. Причины уменьшения численности различны — это и недостаток кормовой базы, и пресс хищников, численность которых при обильном питании резко возрастает, и вспышки эпидемий, и природные катаклизмы. Например, при половодье или наводнении погибает большая часть популяции грызунов, генофонд ее при этом резко и случайно изменяется. . Такой период, когда популяция проходит период малой численности, получил название «бутылочное горлышко». При этом изменяется частота встречаемости различных аллелей генов, частота встречаемости редких аллелей при этом может резко возрасти или вообще исчезнуть.
Резкие возрастания численности часто наблюдаются при попадании видов в новые условия обитания, где благоприятные условия и отсутствуют хищники и паразиты. Так, например, было с распространением опунции и расселением кроликов в Австралии. Опунцию завезли в Австралию и использовали в качестве живой изгороди, в отсутствии естественных врагов она расселилась, и нашествие смогли остановить только с помощью гусениц кактусовой моли, специально привезенной с Американского континента. Благодарные австралийцы даже поставили памятник этому насекомому. Подобная же история случилась и с кроликами, когда в Австралии в 1859 году выпустили 12 пар кроликов. Через 40 лет их количество составляло несколько сотен миллионов особей, справиться с этим нашествием смогли с помощью вирусной инфекции — выпускали в популяции кроликов, зараженных вирусом миксоматоза. Первое время смертность во многих популяциях была стопроцентной, однако впоследствии она снизилась, вероятно появились линии кроликов, невосприимчивые к болезни.
Важным фактором эволюции является и изоляция, препятствующая свободному скрещиванию особей различных популяций. Наследственная изменчивость поставляет и распространяет мутации, популяционные волны и дрейф генов меняют частоту встречаемости различных аллелей, естественный отбор приводит к преимущественному выживанию особей с определенными генотипами, а изоляция препятствует скрещиванию между особями разных популяций. Различают географическую и экологическую изоляцию. Географическая изоляция обычно связана с возникновением естественных преград между популяциями одного вида. При этом не происходит распространения возникших мутаций за пределы популяции, происходит увеличение различий между генофондами различных популяций, что, в конечном счете, приводит к репродуктивной, биологической изоляции — появлению новых видов. Образование различных видов галапагосских вьюрков, сохранение яйцекладущих и сумчатых животных Австралии — результат изоляции. Такое видообразование называется географическим.
Экологическая изоляция связана с различными экологическими условиями, в которых обитают различные популяции. Движущая форма отбора приводит к изменению генофонда популяций, расхождению признаков и, в конечном счете, образованию новых видов. Так, например, образовались различные виды лютиков. Экологическая изоляция может вызываться несовпадением сроков размножения особей различных популяций, например, некоторые лососевые рыбы нерестятся через год, в четный год на нерест приходит одна популяция, в нечетный — другая. Разные популяции форели озера Севан нерестятся в разных горных реках и ручьях, что также приводит к репродуктивной изоляции и может послужить начальным этапом видообразования, называемого экологическим видообразованием.
Приспособленность организмов
Каждый организм удивительно приспособлен к определенным условиям обитания. Эта приспособленность проявляется в особенностях внешнего и внутреннего строения, в поведении, в размножении и заботе о потомстве.
Во внешнем строении ярким примерами приспособленности являются форма тела и особые средства защиты. Например, обтекаемая форма тела рыб и птиц, причудливая форма животных, затаивающихся при поджидании добычи или скрывающихся от врагов (морской конек-тряпичник, рыба-клоун). Колючки ежа и дикобраза защищают этих животных от врагов.
Яркими примерами приспособленности являются защитные окраски животных: покровительственные, предупреждающие, мимикрия (рис. 346). Покровительственная окраска у зеленого кузнечика, богомола, птиц, высиживающих яйца на земле. Предупреждающие окраски у ядовитых или жалящих животных. Например, осы, шмели, божьи коровки несъедобны и своей яркой окраской как бы предупреждают об опасности. Мимикрия — сходство с несъедобными предметами или ядовитыми животными, имеющими предостерегающую окраску. Например, бабочка-стеклянница очень похожа на осу, муха-пчеловидка — на пчелу, муха-шмелевидка — на шмеля, палочник — на веточку.
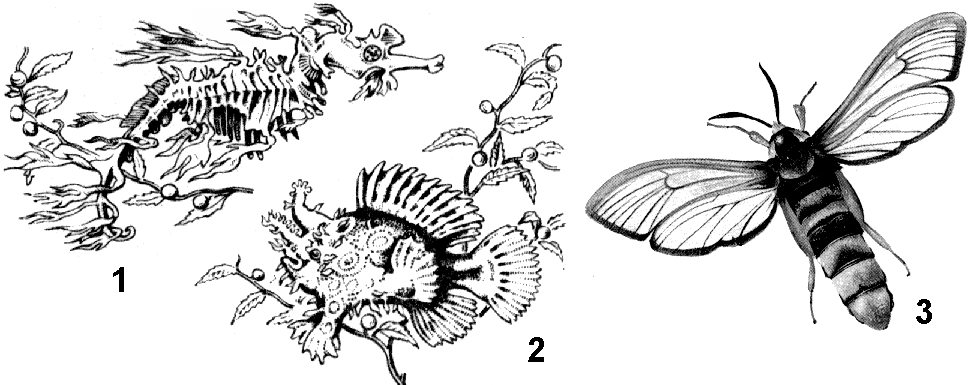
Рис. 346. Приспособительные особенности строения и окраски тела:
1 — морской конек-тряпичник; 2 — рыба-клоун; 3 — бабочка-стеклянница.
Кроме формы тела и окраски, большое значение имеет и приспособительное поведение животных. Например, многие грызуны запасают корм на зиму, некоторые животные затаиваются во время опасности, для многих характерно отпугивающее поведение.
Приспособленность проявляется и в особенностях размножения и заботы о потомстве. Многие рыбы охраняют свою икру (самец трехиглой колюшки даже строит гнездо, плавниками прогоняет воду над отложенной икрой, «пасет» первое время личинок), некоторые вынашивают икру во рту (тиляпия). Если забота о потомстве выражена слабо, то в этом случае у животных очень высокая плодовитость, как это наблюдается у беспозвоночных и низших позвоночных животных, то есть выполняются правила — «чем меньше — тем больше, чем больше — тем меньше» — чем меньше потомства, тем больше забота о нем и наоборот.
Но любая приспособленность относительна: она целесообразна только в конкретных условиях, при их изменении приспособления оказываются бесполезными для организма. Например, иголки спасают ежа на суше, в воде еж разворачивается, становится беззащитным перед лисой.
Таким образом, теория Ч.Дарвина дала ответы на главные вопросы биологической науки: как возникло многообразие и удивительная приспособленность видов. Материал для отбора дает наследственная, мутационная изменчивость, в результате полового размножения (комбинативной изменчивости) эти мутации распространяются и попадают под контроль естественного отбора. В результате отбора из множества разнообразных, ненаправленных мутаций преимущественно выживают особи с полезными для данных условий мутациями. В результате дивергенции, расхождения признаков, различия становятся настолько серьезными, что возникает генетическая изоляция, приводящая к образованию новых видов.
Вид. Критерии вида
Вид — совокупность особей, обладающих наследственным сходством морфологических, физиологических и биохимических особенностей, свободно скрещивающихся и дающих плодовитое потомство, приспособленных к определенным условиям жизни и занимающих в природе определенную область — ареал.
Признаки, по которым виды отличаются друг от друга, называются критериями вида. Различают следующие критерии вида.
Морфологический критерий подразумевает внешнее сходство особей, относящихся к одному виду. Но иногда особи одного вида очень сильно отличаются (такса и дог) или наоборот, есть виды, морфологически почти неотличимые, так называемые виды-двойники, которые не скрещиваются, генетически изолированы. Например, два вида черных крыс: у одного вида в кариотипе 38 хромосом, у другого — 42. Следовательно, одного морфологического критерия недостаточно для определения видовой принадлежности.
Основным является генетический критерий: для каждого вида характерен свой кариотип — свой хромосомный набор. Виды обычно отличаются по числу и строению хромосом. Именно этот критерий обеспечивает генетическую изоляцию, нескрещиваемость между особями разных видов. Даже если появляются межвидовые гибриды, они бесплодны, нарушается процесс образования половых клеток. Но иногда и этот критерий подводит, так как плодовитое потомство может появляться при скрещивании особей, относящихся к разным видам.
Особи одного вида сходны по всем физиологическим процессам — питанию, дыханию, выделению, размножению, что лежит в основе физиологического критерия. Особенно важны отличия в физиологии размножения: в строении полового аппарата, в сроках размножения.
Биохимический критерий — сравнение органических макромолекул у различных видов, в первую очередь сравнение ДНК и белков. По сходству в строении ДНК и белков можно с достаточной вероятностью показать, насколько близкими родственниками являются те или иные виды. Например, гемоглобин шимпанзе по последовательности аминокислот не отличается от гемоглобина человека.
Географический критерий — это территория, на которой обитает данный вид (ареал). У некоторых видов-эндемиков ареал небольшой, есть виды — космополиты, распространенные повсеместно. Но области распространения различных видов часто перекрываются, так что этот критерий не может быть решающим.
Каждый вид приспособлен к определенным условиям существования, к определенным экологическим факторам, которые составляют основу экологического критерия. Например, белый медведь приспособлен к одним экологическим факторам, бурый — к другим.
Для установления видовой принадлежности нельзя опираться на один из критериев, необходимо учитывать их совокупность.
Популяция единица вида и эволюции. Микроэволюция
Вид занимает в природе определенный ареал и существует в виде отдельных территориальных группировок особей, которые в той или иной мере изолированы друг от друга.
Совокупность особей, длительно проживающих на определенной части ареала, относительно обособленно от других совокупностей называется популяцией. Главным объединяющим фактором в популяции является панмиксия — возможность свободного скрещивания между особями одной популяции. Скрещивание особей, относящихся к различным популяциям, затруднено, так как популяции какими-либо преградами отделены друг от друга. Внутри популяции происходит мутационный процесс, а в результате полового размножения происходит распространение возникших мутаций по популяции. Мутации попадают под контроль естественного отбора и генофонд популяции, состоящий из генотипов отдельных особей, постепенно изменяется. Относительная изоляция одной популяции от другой дает возможность сохранить измененный генофонд, если отличия становятся резкими, то образуется подвид, а если возникает репродуктивная изоляция — новый вид.
По Ж.Б.Ламарку эволюционной единицей является особь, т.к. она в течение жизни упражняет или не упражняет различные органы, а затем передает приобретенные признаки следующему поколению.
По современным представлениям единицей эволюции является популяция, в которой происходит изменение генофонда, приводящее к образованию подвидов и видов.
Элементарным эволюционным явлением является стойкое изменение генофонда популяции — изменение частоты встречаемости какого-нибудь аллеля в популяции. Элементарный эволюционный материал – мутации. В популяции постоянно происходит мутационный процесс, мутации случайны и ненаправлены, но мутации, дающие преимущества организмам, в результате полового размножения (комбинативной изменчивости) постепенно распространяются по популяции.
В эволюционном процессе различают два этапа: микроэволюцию, ведущую к образованию новых видов, и макроэволюцию, в результате которой образуются надвидовые систематические единицы (роды, семейства, отряды, классы, типы). Макроэволюция протекает в большие промежутки времени и обычно на больших территориях.
Микроэволюция протекает на уровне популяции в сравнительно небольшие промежутки времени и заканчивается образованием нового вида. В основе микроэволюции и макроэволюции лежат одни и те же эволюционные процессы.
Видообразование
До тех пор, пока происходит обмен генами между различными популяциями, вид сохраняет свою целостность, так как между особями различных популяций возможно скрещивание и образование плодовитого потомства. Как только в силу различных причин возникает репродуктивная изоляция, в результате которой скрещивание между особями разных популяций становится невозможным, популяция становится самостоятельным видом (рис. 349).
Вид является наименьшей генетически устойчивой надорганизменной системой в живой природе и образование нового вида — важный этап в эволюционном процессе.
Различают два способа видообразования: географический, или аллопатрический (от греч. «алло» — разный, «патриа» — родина), и экологический, или симпатрический.
Г
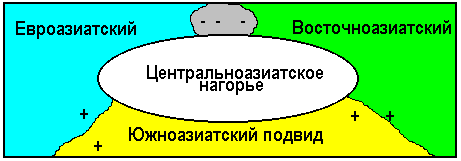
Рис. 347. Ареалы трех подвидов синицы большой.
еографическое видообразование связано или с появлением физических преград, которые приводят к изоляции популяций, или с расширением ареала, при котором популяции попадают в новые условия. И в том и в другом случае в результате микроэволюционных процессов могут образоваться новые виды. Хорошим примером аллопатрического видообразования является образование трех подвидов синицы большой (рис. 347). Южноазиатский подвид скрещивается и дает плодовитое потомство и с восточноазиатским и с евроазиатским подвидами. Но в зоне контакта евроазиатского и восточноазиатского подвидов гибриды не образуются.
Это объясняется отступлением ледника и постепенным расширением ареала птиц на север. В результате адаптаций к различным условиям образовались различные подвиды и в зоне контакта евроазиатского и восточноазиатского подвидов гибриды уже не образуются.
Экологическое видообразование происходит в пределах ареала в силу различных причин. Популяции могут адаптироваться к резко различным условиям обитания, что приводит к образованию различных видов. Например: различные виды лютиков приспособились к жизни в самых разных условиях — в поле, на лугу, в лесу, по берегам водоемов. Один вид традесканции сформировался в тенистых лесах, другой — на солнечных скалистых склонах.
Возможно внезапное образование нового вида, например, при образовании полиплоидов (в роде табака исходное число хромосом равно 12, но есть виды с 24, 48, 72 хромосомами).
В результате обитания в одинаковых условиях, у животных, которые относятся к различным систематическим группам, возникают похожие адаптации, происходит конвергенция — схождение признаков, образование сходных особенностей строения. Причем конвергенция затрагивает лишь определенные признаки, непосредственно связанные с приспособлением к среде.
Например, форма тела акулы (хрящевая рыба), ихтиозавра (вымершее пресмыкающееся), дельфина (млекопитающее) — результат конвергенции у животных различных классов (рис. 348).
Органы, которые выполняют сходные функции, но имеют различное происхождение, называются аналогичными. Например, крылья птицы или летучей мыши — передние конечности, а крылья бабочки — складки хитинового покрова на спине.
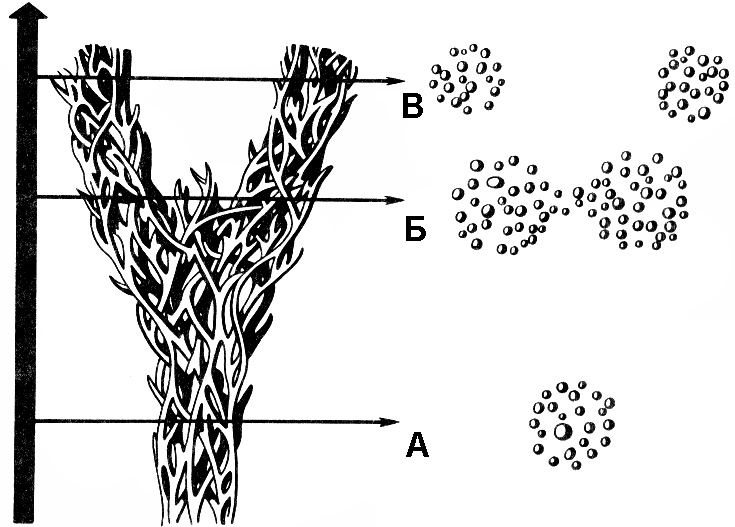
Рис. 349. Схема видообразования (каждая отдельная веточка — популяция):
А — уровень исходного единого вида; Б — момент незавершенного разделения вида; В — два новых вида.
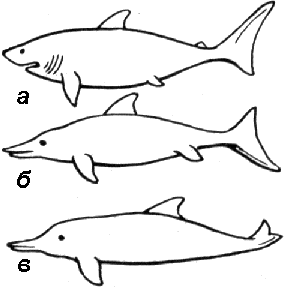
Рис. 348. Конвергенция у водных животных:
а — акула; б — ихтиозавр; в — дельфин.
Главные направления эволюции
Учение о главных направлениях и путях эволюции создано А.Н.Северцовым, который предложил различать два направления в эволюционном процессе — биологический прогресс и регресс, показал три основных пути достижения прогресса.
Биологический прогресс характеризуется увеличением численности, расширением ареала, увеличением числа популяций, ускорением процессов видообразования. В состоянии биологического прогресса находятся, например, насекомые, цветковые растения.
Для биологического регресса характеристики противоположные, в результате возможно полное вымирание этой группы организмов. По пути биологического регресса пошли динозавры, псилофиты, семенные папоротники. В настоящее время вымирание грозит многим видам растений и животных, для спасения которых созданы заповедники, заказники, они занесены в Красные книги.
Состояние биологического прогресса достигается за счет ароморфозов, идиоадаптаций и дегенераций.
Ароморфозы — морфофизиологические изменения, которое приводит к повышению уровня организации, приспосабливают организмы к новым условиям обитания. Ароморфозы приводят к образованию крупных систематических единиц — классов, типов. Например, выход на сушу растений сопровождался появлением механических, проводящих, покровных тканей. Появление семенных растений, опыление в воздушной среде, появление цветка и плода — яркие примеры ароморфозов в растительном мире. В животном мире появление классов позвоночных животных сопровождалось ароморфозами в кровеносной, дыхательной, выделительной и других системах. Например, двухкамерное сердце рыб заменилось трехкамерным у земноводных, у пресмыкающихся в желудочке появляется неполная перегородка, полностью четырехкамерным сердце стало у птиц и млекопитающих. Внутреннее оплодотворение и откладывание яиц позволили пресмыкающимся освоить сушу, роговые покровы препятствуют испарению воды через кожные покровы и служат этой же цели.
Идиоадаптации, напротив, не приводят к повышению общего уровня организации, это такие морфофизиологические изменения, которые приспосабливают организм к конкретным условиям обитания. Идиоадаптации приводят к появлению мелких систематических единиц — родов, семейств, отрядов.
Р

Рис. 350. Идиоадаптации у рыб.
ыбы приспособились к жизни в различных условиях и имеют различную форму тела и окраску (рис. 350), древние земноводные приспособились к различным условиям жизни на Земле и в результате идиоадаптаций дали хвостатых, бесхвостых и безногих земноводных. Иголки у ежа, клюв у утки или дятла, длинная шея и длинные ноги цапли — все это примеры идиоадаптаций. Ароморфозы позволяют освоить новые среды обитания, затем эволюция идет за счет идиоадаптаций (рис. 351).
Дегенерация — третий путь, с помощью которого достигается биологический прогресс (рис. 352). При этом организмы приспосабливаются к более простому образу жизни, в результате происходит упрощение организации.
Н
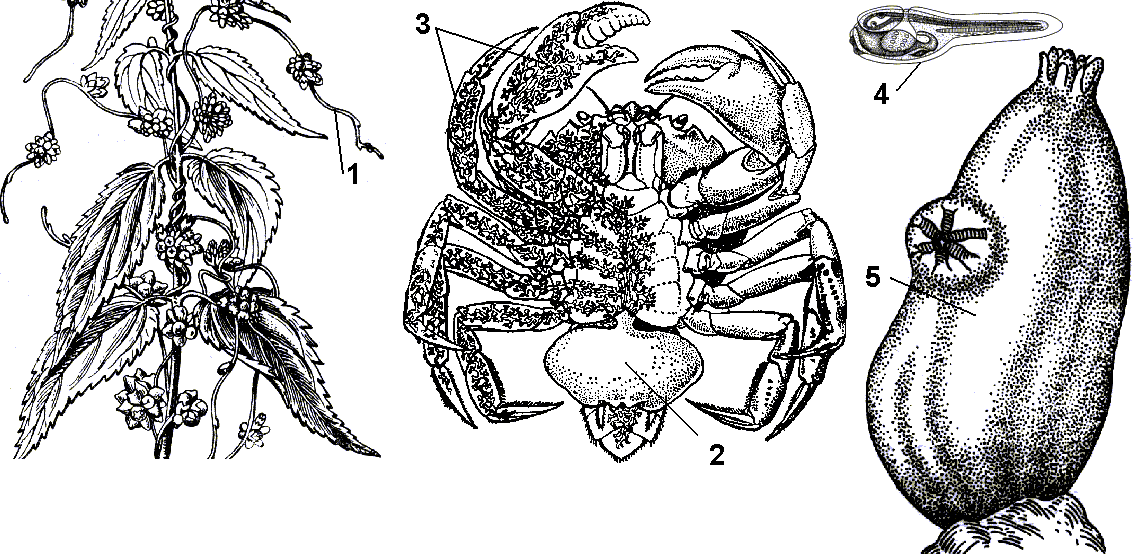
Рис. 352. Примеры дегенераций:
1 — повилика на крапиве; 2 —саккулина; 3 — корневидные отростки саккулины в теле краба; 4 — личинка асцидии с хордой и нервной трубкой; 5 — взрослая асцидия.
апример, переход к сидячему образу жизни у личиночнохордовых привел к редукции хорды и нервной трубки у взрослой асцидии. Паразитический образ жизни ленточных червей привел к полной утрате пищеварительного тракта. У паразитического цветкового растения повилики утратились корни, листья, хлорофилл и питательные вещества растение-паразит поглощает из растения-хозяина с помощью присосок.
Рак саккулина, паразитирующий на крабе, имеет форму мешка, расположенного под брюшком краба, корневидные отростки которого пронизывают все тело краба.
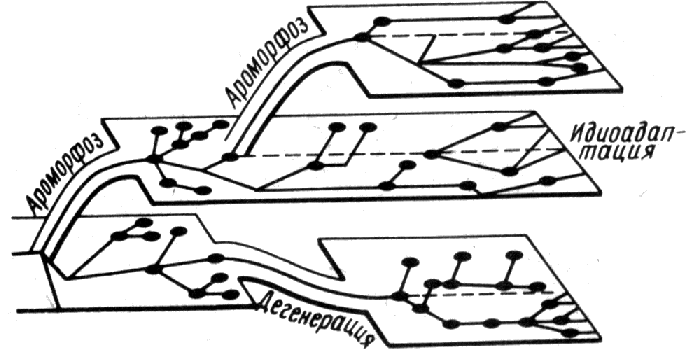
Рис. 351. Соотношение между ароморфозами, идиоадаптациями
и дегенерациями.
Таким образом, к биологическому прогрессу приводят все три пути эволюции: ароморфозы, идиоадаптации и дегенерации
Доказательства эволюции
Доказательства эволюции базируются на данных различных наук: палеонтологии, эмбриологии, анатомии, физиологии, биохимии, биогеографии.
Классические доказательства предоставляет сравнительная палеонтология, изучающая ископаемые организмы, жившие в прошлые эпохи. История развития живых организмов на Земле сохранилась в виде ископаемых остатков.
Прямым доказательством эволюции является ярусность расположения остатков живых организмов: чем более древний слой изучается, тем более примитивные формы жизни в нем находятся, в верхних слоях находят остатки более поздних форм жизни.
Обнаружены ископаемые переходные формы, позволяющие с уверенностью говорить о происхождении той или иной группы организмов. Например, псилофиты — переходная форма от водорослей к высшим наземным растениям, семенные папоротники доказывают образование голосеменных растений от папоротникообразных.
В царстве животных наиболее известны находки археоптерикса (рис. 353), первоптицы размером с голубя, но имеющей многие признаки пресмыкающихся: зубы на челюстях; по три пальца с когтями, выступающие из крыльев; хвост, состоящий из большого количества позвонков с расположенными на нем перьями; наличие брюшных ребер. Ископаемые стегоцефалы имеют признаки рыб и земноводных, котилозавры — признаки земноводных и пресмыкающихся, зверозубый ящер — признаки пресмыкающихся и млекопитающих.
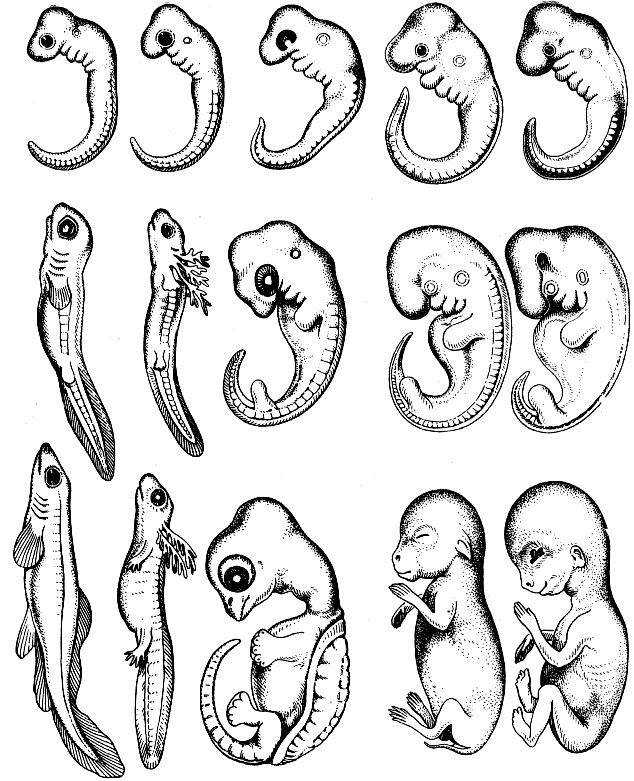
Рис. 354. Эмбрионы рыбы, саламандры, черепахи, крысы и человека сходны на ранних стадиях.

Рис. 353. Реконструкция
археоптерикса.
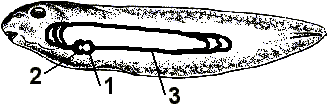
Рис. 355. Схема кровеносной системы головастика:
1 — предсердие; 2 — желудочек; 3 — круг кровообращения.
Палеонтологи по ископаемым остаткам сумели восстановить эволюцию многих групп животных — составлены филогенетические ряды лошади, хоботных, верблюдов.
Сравнительная эмбриология приводит убедительные доказательства в пользу эволюции. Еще Ч.Дарвин обратил внимание на связь между индивидуальным развитием — онтогенезом и историческим развитием вида — филогенезом. Немецкие ученые Э.Геккель и Ф.Мюллер сформулировали биогенетический закон, закон рекапитуляции: "онтогенез — есть краткое и быстрое повторение филогенеза". Например, зародыши млекопитающих на ранних стадиях развития имеют длинный хвост и жаберные щели и похожи на зародыш рыбы, затем на зародыш земноводного, затем — пресмыкающегося, имеют клоаку (рис. 354). Рекапитуляция признаков объясняется тем, что на разных стадиях включаются сохранившиеся гены далеких предков — рыбы, земноводного, пресмыкающегося, обезьяны. Головастик лягушки имеет рыбообразную форму тела, боковую линию, двухкамерное сердце и один круг кровообращения (рис. 355). Он как бы повторяет признаки рыб, далеких предков земноводных.
Сравнительная анатомия показывает, что план строения позвоночных животных одинаков у различных классов. Например, в скелете земноводных, пресмыкающихся, птиц и млекопитающих различают четыре отдела: скелет головы, туловища, конечностей и поясов конечностей. Отделы состоят из одинаковых костей, различия в строении появляются в результате приспособлений к конкретным условиям среды. Сходный план строения имеют и остальные системы органов, и отдельные органы. Для всех птиц, например характерна правая дуга аорты, а для млекопитающих — левая.
В пользу эволюции свидетельствуют рудименты — органы утратившие свои функции и находящиеся на грани исчезновения (волосяной покров на конечностях и туловище у человека, копчик — рудимент хвоста, состоящий из 3 — 4 позвонков, остатки тазового пояса у кита).
К сравнительно анатомическим доказательствам относятся и атавизмы — случаи возврата к признакам предков (у человека — случаи рождения с детей хвостом, с дополнительными парами сосков).
Физиологическими доказательствами служат сходства различных физиологических процессов у животных близких систематических групп: сходство в физиологии кровеносной, дыхательной, выделительной, пищеварительной, половой систем органов. Например, беременность человека и человекообразных обезьян продолжается около девяти месяцев и протекает сходно.
Данные биохимии, вероятно, одни из самых убедительных. Доказано, что у близкородственных групп организмов белки сходны по аминокислотному составу и сходна нуклеотидная последовательность ДНК в хромосомах. Например, гемоглобин человека и шимпанзе идентичен по аминокислотному составу, а между гемоглобином человека и гориллы отличия в двух аминокислотах. Сравнение аминокислотной последовательности в рибосомальных белках и нуклеотидной последовательности в рибосомальных РНК (как одних из наиболее консервативных) подтверждает классификацию всех основных групп живых организмов.
В настоящее время проведена гибридизация цепей нуклеотидов ДНК человека и шимпанзе. Для этого были разделены двойные цепи ДНК человека и шимпанзе, и затем одиночные цепи ДНК человека соединили с цепями нуклеотидов шимпанзе. Между комплементарными нуклеотидами восстановились химические связи, и оказалось, что ДНК человека и шимпанзе сходны на 91 — 92% (рис. 356).

Рис. 356. Метод гибридизации ДНК.
В составной ДНК человека и шимпанзе все химические связи восстановились, кроме тех мест, где нуклеотиды не комплементарны.
Кроме того, универсальность генетического кода и единство аминокислотного состава белков свидетельствуют о происхождении всех живых организмов Земли от единого предка.
Интересные факты приводит и биогеография, наука о распространении растительного и животного мира. Поверхность Земли разделена на шесть биогеографических областей: 1) Палеоарктическую (Европа, Северная Африка, Северная и Средняя Азия, Япония); 2) Неоарктическую (Северная Америка); 3) Эфиопскую (Африка к югу от Сахары); 4) Индо-малайскую (Южная Азия и Малайский архипелаг); 5) Неотропическую (Центральная и Южная Америка); 6) Австралийскую.
Фауна и флора Палеоарктической и Неоарктической областей сходны, хотя и между ними находится Берингов пролив. Сходство объясняется тем, что в недалеком прошлом существовал сухопутный мост — Берингов перешеек. Эти две области объединены Голарктическую область.
Различия в растительном и животном мире между Неоарктической и Неотропической областями объясняются тем, что Панамский перешеек появился недавно. Только немногие виды сумели проникнуть в Северную Америку (броненосец, опоссум) и из Северной Америки в Южную.
Австралия отделилась от остальных материков более 100 млн. лет назад, тогда еще не было плацентарных животных, и изоляция сохранила примитивных яйцекладущих и сумчатых млекопитающих.
Фауны и флоры различных областей отличаются потому, что виды сформировались в определенном центре происхождения и расселялись до тех пор, пока не встречали какой-то естественной преграды.
Основные положения СТЭ.
- Материалом для эволюции служат, как правило, мелкие дискретные изменения наследственности - мутации.
- Мутационный процесс, волны численности - факторы-поставщики материала для отбора - носят случайный и ненаправленный характер.
- Единственный направляющий фактор эволюции - естественный отбор, основанный на сохранении и накапливании случайных и мелких мутаций.
- Наименьшая эволюционная единица - популяция, а не особь, отсюда особое внимание к изучению популяции как элементарной структурной единицы вида.
- Эволюция носит дивергентный характер, т.е. один таксон может стать предком нескольких дочерних таксонов, но каждый вид имеет единственный предковый вид, единственную предковую популяцию.
- Эволюция носит постепенный и длительный характер. Видообразование как этап эволюционного процесса представляет собой последовательную смену одной временной популяции чередой последующих временных популяций.
- Вид состоит из множества соподчиненных морфологически, биохимически, экологически, генетически отличных, но репродуктивно не изолированных единиц - подвидов и популяций. Однако известно немало видов с ограниченными ареалами, в пределах которых не удается расчленить вид на самостоятельные подвиды, а реликтовые виды могут состоять из единственной популяции. Судьба таких видов, как правило, недолговечна.
- Обмен аллелями, "поток генов" возможны лишь внутри вида. Если мутация имеет положительную селективную ценность на территории ареала вида, то она может распространиться по всем его популяциям и подвидам. Отсюда определение вида как генетически целостной и замкнутой системы.
- Поскольку основной критерий вида - его репродуктивная изоляция, то этот критерий не применим к формам без полового процесса (огромному множеству прокариот, низшим эукариотам).
- Макроэволюция, или эволюция на уровне выше вида, идет лишь путем микроэволюции. Не существует закономерностей макроэволюции, отличных от микроэволюционных.
- Исходя из всех упомянутых положений ясно, что эволюция непредсказуема, имеет ненаправленный к некоей конечной цели характер. Иначе говоря, эволюция не носит финалистический характер.
Основные вопросы для повторения
- Кто из ученых реформировал ботанический язык - ввел около 1000 ботанических терминов, описал 1200 новых родов и 8000 новых видов?
- Кто из ученых считал, что благоприобретенные признаки передаются по наследству?
- Кто из ученых предложил теорию градации, разделил животный мир на 14 классов и расположил их на 6 ступенях?
- Кто из ученых считал, что всем организмам присуще стремление к прогрессу?
- Кто из ученых, сравнив найденные в Южной Америке ископаемые останки вымерших животных с современными, предположил их родство?
- Какой фактор Ч. Дарвин считал главным в создании человеком сортов растений и пород домашних животных?
- Какой фактор Ч. Дарвин считал главным, направляющим фактором, который приводит к приспособленности организмов к среде?
- Что по Ч. Дарвину лежит в основе естественного отбора?
- Что является единицей эволюции по Ламарку? По Дарвину?
- Какой критерий можно считать главным, основным видовым критерием?
- Между гемоглобином шимпанзе и человека в последовательности аминокислот отличий нет, а гемоглобин человека и лошади имеет 42 отличия. Какой это критерий?
- Какой вид изменчивости по Ч. Дарвину называется определенной?
- Какой вид изменчивости без изменения генотипа приспосабливает организмы к среде обитания?
- Какие виды изменчивости дают материал для эволюции, т.к. связаны с изменением генотипа или перекомбинацией генов?
- Какой вид изменчивости дает материал для эволюции, т.к. изменения одного гена влияет на проявление многих признаков?
- Какой вид изменчивости у листьев, сорванных с одного дерева?
- Какой вид изменчивости у брата и сестры?
- Какая форма изменчивости не должна использоваться селекционерами для создания нового сорта или породы?
- Какой вид изменчивости поставляет материал для искусственного отбора, т.к. приводит к изменению генов, хромосом, хромосомного набора?
- Какой вид изменчивости поставляет материал для искусственного отбора, когда человек для скрещивания подбирает особей с нужными ему признаками?
- Кто разработал учение о стабилизирующей форме естественного отбора?
- Кто разработал учение о движущей форме естественного отбора?
- Какой фактор эволюции привел к образованию на Галапагосских островах различных видов вьюрков; сохранил яйцекладущих и сумчатых млекопитающих в Австралии?
- Как называется процесс расхождения признаков при образовании новых видов?
- Чем заканчивается микроэволюционный процесс?
- Что является единицей эволюции?
- Один вид традесканции сформировался на скалистых склонах, другой — в тенистом лесу. Какой способ видообразования привел к образованию этих видов?
- Какая форма отбора сохраняет виды неизменными при неизменных условиях?
- Какая форма приводит к образованию новых видов, когда происходит изменение условий существования?
- Дизруптивный отбор?
- Дестабилизирующая форма отбора?
- Дрейф генов как фактор эволюции?
- Популяционные волны как фактор эволюции?
- Изоляция как фактор эволюции?
- Представления какого ученого не могут объяснить появление защитных окрасок у насекомых, раковин у моллюсков?
- Что по Ж. Б. Ламарку привело к приспособленности организмов?
- Что по Ч. Дарвину привело к приспособленности организмов?
- Приведите примеры аллопатрического видообразования.
- Приведите примеры симпатрического видообразования.