
 Авторефераты по всем темам >>
Авторефераты по биологии
Авторефераты по всем темам >>
Авторефераты по биологии
Типологические особенности при комплексной коррекции у крыс
Автореферат докторской диссертации по биологии
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
 СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА
СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТАCнизу к ящику крепится электродный пол. Сверху ящик закрывается прозрачной крышкой. Каждый блок имеет сложное строение и состоит из двух модулей - призм, которые в исходном состоянии жестко сцеплены друг с другом.
Во внешних стенках модулей высверлены специальные отверстия, предназначенные для крепления различных вспомогательных устройств, например, манипуляторных колец. Кроме того, в отверстия можно вставлять цилиндрические штифты, что обеспечит прочную фиксацию блоков друг с другом. Размер ящика и количество блоков в нем регламентируется только пожеланиями или потребностями экспериментатора.
В исходном состоянии весь ящик заполнен блоками. Сложное строение блоков позволяет посредством рассоединения модулей - призм создавать инструментальные среды практически любой формы: гексагональной, ромбовидной и т.д. Модульный блок может быть при необходимости модифицирован в компонент системы регистрации (с помощью крепления чувствительных датчиков (фотоэлементов, герконов) в отверстиях модулей - призм). Для того, чтобы в установке сформировать, например, простой Т-образный лабиринт, экспериментатором определяется расположение стартовой зоны и путем вынимания из ящика блоков создаются необходимой длины ход и рукава требуемого лабиринта.
Вставка кольца в отверстие модуля - призмы блока расширяет возможности установки и превращает ее в среду, на которую подопытное животное имеет возможность воздействовать и изменять ее в соответствии со своими мотивациями и рассудочно-познавательною деятельностью мозга: дергая кольца зубами или лапками, крыса перемещает блоки и изменяет конфигурацию лабиринта.
В эксперименте технически использовалось три модифицированных варианта лабиринтов, которые формировались в устройстве. Из них: вариант А - наиболее простой, имеющий Т-образную форму, архитектурно повторяющий известный классический Т-образный лабиринт. Вариант В (рис. 9) - в отличие от предыдущего варианта А, представлял усложненную структуру, формирующуюся при помощи модулей. Вариант С (рис. 9) - являлся наиболее сложным, хотя при этом в его центре имелась кольцевая развязка, с помощью которой можно было животному переходить из одного отсека в другой.
 аа
аа 
аа Вариант В аВариант С
а Рисунок 9 - Варианты лабиринтов, формирующиеся в универсальном устройстве с изменяющейся архитектурой.
аа Предварительно лабораторные животные с помощью универсальной проблемной камерыа были разделены на три типа (с низкими (тип I), средними (тип II) и высокими (тип III) когнитивными способностями) по разработанному нами методу.
В первой серии опыта у всех экспериментальных особей регистрировались показатели ориентировочно-исследовательского поведения (в течение трех суток по 3 минуты), среди которых фиксировали следующие значения: время общей двигательной активности, время замирания, количество вертикальных стоек, дефекаций, принюхиваний. Дополнительно производился этологический анализ груминга (по А.В. Калуеву). Затем были сформированы две группы, содержащие по 7 крыс из каждого типа: группа № 1 - контрольные животные, не подвергающиеся пищевой депривации и содержащиеся в обычном режиме; группа № 2 - особи, подверженные трехсуточной пищевой депривации. Далее осуществлялась вторая серия, перед которой на предварительном этапе животные всех групп подвергались трехсуточной пищевой депривации и помещались в лабиринты (последовательно по мере усложнения архитектуры). За отсеком, имеющим наружный выход, располагалось пищевое подкрепление. В тупиковых отделах этого не было, в них животные могли лишь только осязать запах пищи. В Т-образном лабиринте (вариант А), как и при других, более сложных его модификациях, было только по одному выходу. Входные отсеки при помещении крысы в установку сразу же блокировались. Данная процедура проводилась троекратно.
После десятидневного перерыва начиналась первая серия эксперимента, после окончания которой (вновь с десятидневным перерывом) приступали к выполнению второй серии, где фиксировали время поисковых действий, интенсивность побежек (количество посещенных отсеков в минуту), ошибочный коэффициента (процентное соотношение ошибочных и правильных побежек).
Анализируя в 1-е сутки тестирования время общей активности, как исследовательский компонент, обращает на себя внимание тенденция к плавному снижению данного показателя у представителей всех типов крыс на усложнение архитектуры.а У II типа крыс время движения в лабиринте С (в сравнении с вариантом А) достоверно снизилось в 1,5 раза (р<0,05).а
В следующие сутки наблюдения подобная тенденция изменения данного параметра сохранилась. При этом наиболее выраженное снижение отмечалось у животных I типа, где значение времени двигательной активности в лабиринте С (в сравнении с А) уменьшилось более чем в 2 раза (р<0,05). Кроме этого, в данной категории достоверная разность обнаруживалась между 2-ми и 1-ми сутками (в варианте С), снизившись на 46,6 с (р<0,05). В последние сутки тестирования в каждой типологической группе отмечалось незначительное уменьшение времени общей двигательной активности по мере усложнения архитектуры лабиринта (р>0,05).
Количество вертикальных стоек в 1-е сутки наблюдения по мере усложнения структуры устройства снижался у представителей I и II типа, где разница между простым и наиболее сложным вариантом лабиринта изменилась в 1,6 раза (р<0,05) и в 2,1 раза (р<0,05) соответственно. У экспериментальных особей III типа прослеживалась волнообразная тенденция изменения количества вертикальных стоек. В сформированных трех вариантах лабиринта максимальное количество стоек (в 1-е сутки) выявлено у особей, относящихся к I типу, минимальное - у крыс второй типологической выборки и соответственно среднее у крыс из III типа во всех архитектурных вариантах лабиринта.
Количество принюхиваний, как показателя, отражающего также исследовательское поведение, у представителейа I типа в 1-е сутки снизилось (в варианте С по сравнению с вариантом А) в 1,9 раза (р<0,05), во 2-е в 1,3 раза (р>0,05), а в 3-е сутки наблюдения тенденция по мере усложнения лабиринта изменилась в обратную сторону, количественные показатели которой увеличились в 1,7 раза (р>0,05).
Данный параметр у животных III типа изменялся с одинаковой динамикой (при усложнении лабиринта), незначительно снижаясь в каждый день исследования.
У II типа не было выявлено какой-либо закономерности к его изменению, а во 2-е (в лабиринте В) и 3-е (в лабиринте А) сутки этот компонент поведения отсутствовал.
Параметры, характеризующие у испытуемых лабораторных животных уровень тревожности, также имели зависимость от типологической принадлежности и уровня сложности лабиринта.
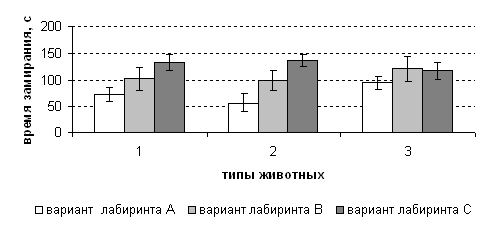
Рисунок 10 - Динамика изменения времени замирания у лабораторных крыс различной типологической принадлежности в разных вариантах лабиринта.
аа Так, показатель времени замирания, фиксируемый во время тестирования по мере усложнения структуры устройства (вариант А>В>С) в 1-е сутки наблюдения (рис. 10) возрос (в варианте С по сравнению с А) у животныха I типа в 1,8 раза (р<0,05), II типа в 2,4 раза (р<0,05) и III типа в 1,2 раза (р>0,05).
У особей I и II типа во 2-е сутки исследования выявлена схожая динамика, но при этом разность значений в сравниваемых категориях была выражено очень слабо. А в III типе вообще отсутствовала какая-либо динамика к увеличению или уменьшению данного показателя.аа
Среди показателей исследовательской сферы поведения большое внимание было уделено количеству возникающих грумингов, которые несут за собой неоднозначный характер и могут интерпретироваться по-разному. Большинство исследователей в подобных работах не проводят детальный анализ данного показателя, относя его к явлению, характеризующему составляющую тревожности. Но как было выяснено в наблюдениях Калуева А.В. (2002) этот компонент поведения может носить как признаки тревожности, так и показывать состояние поведенческого комфорта (в эксперименте на грызунах).
В связи с этим нами был проведен детальный этологический анализ груминга (по Калуеву А.В.) у крыс с разными типологическими особенностямиа в разной инструментальной обстановочной афферентации.
У животных из I типологической группы в вариантах лабиринта А и В превалировал груминг комфорта. Так, в лабиринте А данный вид груминга в долевом соотношении составил 31 %. По мере усложнения лабиринта доля, приходящая на данный вид поведенческого компонента значительно уменьшалась до 22 % (в варианте В) и до 10 % (в варианте С).
В простом варианте используемой установки (А) среди индексов тревожности доминировал индекс Г (21 %), при котором остановки животное совершает между стадиями груминга (поведение после остановки начинается с последующей стадии, а не продолжает предыдущую)а и А (19 %), отражающий незавершенный характер груминга (процент незавершенных актов). В лабиринте (В) доля данного индекса составила 16 %.
Лабораторные крысы II Цго поведенческого типа проявили максимальную выраженность компонента комфорта в наиболее простом варианте лабиринта (А), составившую 56 %. При этом у животных I типа значение данного компонента было ниже в 1,8 раза, а III типа в 2,2 раза. В лабиринте В равные доли приходились на индекс Г, Д и Ж (по 16 %). Следует отметить, что индекс Ж у II типа, в отличие от I и III, имел максимальное значение во всех вариантах лабиринта, в то время как у I типологической выборки он приобрел минимальные значения (до 3 %), в III средние (до 7 %). В зависимости от сложности лабиринта индекс Ж иногда отсутствовал. Согласно используемой нами шкале индекс Ж аустанавливался в тех случаях, когда непосредственно после стрессорного груминга наблюдались другие формы стрессорного поведения (агрессия или аутоагрессия, оценка риска, замирание-фризинг, бегство, защита и т.п.). Отношение числа стрессорных пост-груминговых реакций к общему числу всех пост-груминговых поведенческих реакций служило дополнительным косвенным признаком груминговой тревожности в эксперименте.
Таким образом, мы констатируем, что один из самых неблагоприятных компонентов отставочного поведения было присуще лабораторным животным II поведенческого типа.
Экспериментальные особи III типа имели практически одинаковую долю, приходящуюся на компонент-комфорта в различных вариантах лабиринта. Из индексов тревожности в лабиринте А и В максимальное значение составил индекс В, для которого были характерны длительные тревожногенные остановки, длящиеся больше, чем это происходит в у животного в обычной обстановочной афферентации. В варианте С, как наиболее архитектурно сложном, доминирующие значение среди компонентов тревожности приобрел индекс Б, характеризующийся нарушением нормальной последовательности стадий груминга (происходило либо выпадение очередных стадий, либо их инверсия).
Исходя из полученных данных, необходимо отметить, что разработанное устройство позволяет детально проанализировать психобиологические особенности у лабораторных крыс с различной типологической принадлежностью в различных условиях.
Параметры поведенческой сферы при применении производных дигидрокверцетина
При алгоритмическом исследовании биологической активности производных дигидрокверцетина из 11 соединений наиболее работающими в отношении поведенческой сферы проявили себя вещества НК-2 и НК-1 (в меньшей степени). Данные соединения синтезированы на кафедре органической химии химического факультета Московского педагогического государственного университета (Коротеев М.П., Кухарева Т.С., Коротеев А.М., 2007).
Проанализировано влияние НК-2 (6-диизопропиламинометил-2,3-дигидрокверцетина, С22p7NO7) на инструментальное поведение в МУ в сравнительном аспекте дигидрокверцетином (являющимся непосредственным предшественником), а также с известными веществами, обладающими антиоксидантными свойствами - мексидолом и реамберином.
Предварительно перед экспериментом животные, испытывающие голод, помещались в МУ, в котором все выходы были открыты. Подопытные особи на фоне определенной обстановочной афферентации могли воспользоваться любым выходом. Данная процедура проходила троекратно, после чего у животных контрольной и подопытных групп перед экспериментом производиласьа депривация пищи (в течение 3-х суток). На следующий день начинался эксперимент, проводимый один раз. Животные принадлежали к II поведенческому типу.
Показатели, полученные в МУ, свидетельствуют о наличии вторичных ноотропных свойств у всех исследуемых нами препаратах (табл. 5), что проявляется в виде оптимизации процессов обучения в используемом устройстве. Наиболее высокая разница по всем регистрируемым параметрам получена у экспериментальных особей, получающих мексидол.
В данной группе значение латентного периода уменьшилось на 128,0 с (р<0,05) по сравнению с контрольными особями. Значение времени достижения подкрепления также имело достоверно значимую разницу, уменьшившись на 55,9 с (р<0,05). Показатель безошибочности побежек, отражающий когнитивную составляющую процесса обучения, достоверно снизился практически в 2 раза.
В группе подопытных животных, получавших реамберин, достоверно значимая разность выявлена только между временем достижения подкрепления у экспериментальных особей на фоне введения плацебо (контроль) и реамберина (46,2 с, р<0,05). У животных, получавших дигидрокверцетин, значительно, но без достоверной разницы снизилось время достижения подкрепления на 44,3 с и показатель безошибочности побежек на 14,5 %. Латентный период у данной исследуемой группы снизился в отличие от контрольной на 116,1 с (р<0,05).
Таблица 5.
Показатели обучаемости лабораторных животных в модульном устройстве под влиянием НК-2, дигидрокверцетина, мексидола и реамберина
Показатели |
Подопытные группы |
||||
контроль |
НК-2 |
дигидро-кверцетин |
мексидол |
реамберин |
|
атентный период, (с) |
297,444,3 |
177,821,6* |
181,333,7* |
169,431,2* |
228,925,8 |
Время дос- жения под- крепления, (с) |
187,327,8 |
154,217,9 |
143,024,3 |
131,413,3* |
141,110,3* |
Показатель безошибоч-ности побе- жек, в (%) |
59,313,8 |
31,18,5* |
44,87,7 |
30,510,1* |
47,16,2 |
Примечание: * - p<0,05, апо сравнению с контрольной группой
аВ группе экспериментальных животных, получавших НК-2, аналогично снизились все показатели. Достоверно значимое уменьшение отмечалось в значениях латентного периода и показателя безошибочности побежек. Значение времени достижения подкрепления, характеризующее в большей степени энергетическую (физическую) сферу обучения животных значительно снизилось на 28,2 %, не имея при этом достоверности.аа
Полученные результаты показали, что все исследуемые антиоксиданты, в том числе и новое соединение НК-2, проявили выраженный позитивный эффект в отношении познавательной сферы, о чем свидетельствуют наблюдаемые нами значения когнитивного уровня (а именно показателя безошибочности побежек), оптимизация условно-рефлекторной деятельности. Кроме этого, мы предполагаем, что данное соединение, достоверно снизив значение латентного периода, проявляло эффект снижения общего уровня тревожности (аналогично с известными сравниваемыми препаратами). Нейротропная активность антиоксидантов мексидола и реамберина, на сегодняшний день, является общеизвестным фактом, о чем свидетельствуют как данные других исследователей (Молодавкин Г.М. и соавт., 2007; Александровский Ю.А. и соавт., 1997), так и наши предыдущие работы.
Таким образом, внутрибрюшинное введение НК-2, дигидрокверцетина, мексидола и реамберина в дозах по 25 мг/кг, оказывало стимулирующее воздействие на элементарную рассудочную деятельность животных, улучшая способность к обучениюа достижения подкрепления в модульном устройстве.
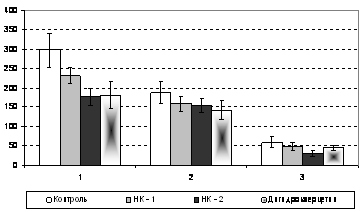
Рисунок 11 - Поведенческие показатели в модульном устройстве при применении дигидрокверцетина и его производных (НК-1 и НК-2). По оси абсцисс: 1 - латентный период (в секундах), 2 - время достижения подкрепления (в секундах), 3 - показатель безошибочности побежек (в %).
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
Авторефераты по всем темам >>
Авторефераты по биологии