
 Авторефераты по всем темам >>
Авторефераты по биологии
Авторефераты по всем темам >>
Авторефераты по биологии
Типологические особенности при комплексной коррекции у крыс
Автореферат докторской диссертации по биологии
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
 СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА
СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТАТаблица 1
Поведенческие параметры в универсальной проблемной камере у животных с низкими когнитивными способностями, получавших мексидол
ПА |
Мотивация |
1 сутки |
2 сутки |
3 сутки |
4 сутки |
5 сутки |
ВП |
Оборонительная |
173,71,3 |
143,29,2 |
98,49,3 |
71,314,2 |
57,712,9* |
Пищевая |
235,732,1 |
196,819,9 |
104,423,1 |
84,314,2 |
77,718,6** |
|
Питьевая |
123,47,7 |
94,54,3 |
81,63,9 |
66,34,8 |
54,83,7** |
|
ИП |
Оборонительная |
17,11,4 |
14,80,9 |
16,41,8 |
13,2 1,7 |
10,20,7* |
Пищевая |
23,10,6 |
21,41,3 |
21,11,7 |
19,31,5 |
17,40,9* |
|
Питьевая |
13,71,2 |
12,50,3 |
11,80,7 |
10,11,1 |
9,30,8* |
|
КП |
Оборонительная |
41,34,4 |
48,75,7 |
49,33,1 |
55,36,1 |
63,44,9* |
Пищевая |
21,12,4 |
29,43,1 |
37,56,1 |
41,15,1 |
48,34,3** |
|
Питьевая |
25,71,2 |
29,42,5 |
32,72,4 |
34,32,7 |
35,52,8** |
|
КСОП |
Оборонительная |
13,11,3 |
9,21,4 |
5,41,8 |
3,11,1 |
2,70,9** |
Пищевая |
17,81,1 |
11,31,4 |
8,60,7 |
5,51,2 |
3,21,3* |
|
Питьевая |
14,31,9 |
13,11,2 |
7,40,6 |
4,30,8 |
3,80,7** |
Примечания: * - помечены показатели ПА при р<0,05, а** - при р<0,01, а*** - при р<0,001(сравнивались 1 и 5 сутки тестирования).
Наиболее высокую достоверно значимую разницуа среди параметров когнитивной сферы имел КСОП. Так, при оборонительной мотивации в контроле он уменьшился в 2,7 раза (р<0,01), тогда как в подопытной группе в 4,9 раз (р<0,001, к 5 суткам в сравнении с 1); в пищевой методике у интактных особей к 5 суткам снизился в 2,3 раза (р<0,01), у исследуемых в 5,6 раза (р<0,001); в питьевом поведении у контрольных уменьшился в 4,7 раза (р<0,01) и у подопытных в 3,8 раза (р<0,01). Таким образом, выявлено достоверно значимое влияние мексидола на параметры поведения в ПК при ИЭС у экспериментальных животных.
Количественные значения качественных и количественных показателей поведения в структуре пищевой и питьевой мотивации подробно отражены в таблице 1.
КП при оборонительной мотивации у животных, получающих эмоксипин, превышал аналогичный показатель контрольных в 1 сутки на 8,4 % (р<0,05), на 2 сутки на 8,1 % (р>0,05), на 3 сутки на 10,4 % (р>0,05), на 4 сутки на 4,3 % (р>0,05) и в 5 сутки на 10 % (р<0,05). В пищевой методике КП имел аналогичную тенденцию. В 1 сутки он превысил значение аналогичного показателя контрольной группы на 5,3 % (р>0,05), во 2 сутки на 9,3 % (р>0,05), в 3 сутки на 19,4 % (р<0,05), в 4 сутки на 25 % и в 5 сутки на 27,1 % (р<0,05). При выработанной питьевой мотивации КП в 1 сутки незначительно увеличился на 2,8 % (р>0,05), на 2 сутки на 3,8 % (р>0,05), на 3 сутки на 2,9 % (р>0,05), на 4 сутки на 6,5 % (р<0,05)а и в последние сутки на 7,9 % (р<0,05).
В оборонительном поведении наименьшие значения КСОП наблюдались у животных, получавших эмоксипин. В 1 сутки КСОП у подопытной группы был ниже на 46,9 % (р<0,01), во 2 сутки на 41,7 % (р<0,05), в 3 сутки на 30,6 % (р>0,05), в 4 сутки на 17,3 % (р>0,05), в 5 сутки на 40,3 % (р<0,05). В структуре пищевой мотивацииа у данного параметра прослеживалась аналогичная динамика. В 1 сутки КСОП снизился на 35,1 % (р<0,05), во 2 сутки на 34,6 % (р<0,05), в 3 сутки на 42,4 % (р<0,05), в 5 сутки на 37,7 % (р>0,05). При сформированном ПИР КСОП был достоверно ниже во все дни исследования, кроме последнего: в 1 сутки уменьшился на 46,4 %а (р<0,05), во 2 сутки на 55,5 % (р<0,05), в 3 сутки на 52,2 % (р<0,05), в 4 сутки на 55,8 % (р<0,05) и в последний день тестирования на 29,5 % (р>0,05).
Влияние пантолизата на поведенческую сферу лабораторных животных с низкими когнитивными способностями
В данной части исследования мы наблюдали за изменением поведения при ИЭС крыс с низкими когнитивными способностями под воздействием пантолизата, продукта, полученного в результате переработки отходов фармацевтического производства пантов (Ярцев В.Г., 1994).
По аналогичному алгоритму перед формированием ИЭС в ПК производилась выработка инструментальных рефлексов: оборонительного, пищевого и питьевого (количественные значения показаны в таблице 2).
Таблица 2
Время формирования условных рефлексов контрольных животных и животных, получавших пантолизат
Рефлексы |
1 сутки |
2 сутки |
3 сутки |
4 сутки |
5 сутки |
|
Контроль |
ИРАИ |
481,32,3 |
398,530,6 |
301,843,8 |
213,442,1 |
184,516,9* |
ИПР |
494,77,4 |
385,671,3 |
294,018,6 |
204,029,4 |
113,325,6* |
|
ПИР |
386,14,3 |
303,439,5 |
214,759,4 |
181,423,5 |
99,716,3* |
|
Опыт |
ИРАИ |
394,60,8 |
343,221,5 |
294,79,8 |
223,81,3 |
184,624,6* |
ИПР |
407,84,5 |
334,918,3 |
241,72,4 |
179,53,0 |
107,414,5* |
|
ПИР |
313,78,9 |
286,421,3 |
206,54,8 |
135,79,7 |
76,51,7** |
Примечания: * - помечены показатели ПА при р<0,05, ** - при р<0,01, (сравнивались 1 и 5 сутки обучения)
В УПК при оборонительном поведенииа параметры мотивационно-энергетической сферы изменялись у подопытных животных. ВП уменьшилось в 1 сутки на 14,3 (р<0,05), во 2 сутки на 26,7 % (р<0,05), в 3 сутки на 19,6 % (р<0,05), в 4 сутки на 38,2 % (р<0,05) и в 5 сутки на 30,2 % (р<0,05); ИП снизилась в 1 сутки на 11 % (р>0,05), во 2 сутки на 20,4 % (р<0,05), в 3 сутки на 41,5 % (р<0,05), в 4 сутки на 38,7 % (р<0,05), в 5 сутки на 43,6 % (р<0,05). Параметры когнитивной сферы позитивно изменялись во все дни исследования. КП возрос в 1 сутки на 17,4 % (р>0,05), во 2 сутки на 35,3 (р<0,05), в 3 сутки на 32,6 % (р<0,05), в 4 сутки на 32,3 % (р<0,05) и в последние сутки тестирования на 71,6 % (р<0,05); КСОП достоверно уменьшился в 1 сутки на 25 % (р<0,05), во 2 сутки на 23,3 % (р<0,05), в 3 сутки на 43,2 % (р<0,05), в 4 сутки на 58,7 % (р<0,05) и в 5 сутки на 59,8 % (р<0,05).
В структуре пищедобывательного поведенияа ВП у исследуемых особей, получавших пантолизат, имело тенденцию в сторону снижения, уменьшаясь в 1 сутки на 30,8 % (р<0,05), во 2 сутки на 32,7 % (р<0,05), в 3 сутки на 36,5 % (р<0,05), в 4 сутки на 30,7 % (р>0,05), в 5 сутки на 17,6 % (р>0,05). ИП снизилась в 1 сутки на 19,0 % (р<0,05), во 2 сутки на 16,7 % (р>0,05), в 3 сутки на 23,2 % (р<0,05), в 4 сутки на 25,6 % (р<0,05), в 5 сутки на 36,2 % (р<0,01). КП при данном виде поведения у подопытных особей увеличился в 1 сутки на 25,7 % (р>0,05), во 2 сутки на 29,6 % (р<0,05), в 3 сутки на 30,2 % (р<0,05), в 4 сутки на 24,5 % (р<0,05) и в 5 сутки на 29,4 % (р<0,01). КСОП в опытной группе в течение пяти суток снижался, стремясь к оптимальному единичному значению. По сравнению с контролем в 1 сутки КСОП был ниже на 23,3 % (р<0,05), во 2 сутки на 41,7 % (р<0,01), в 3 сутки на 35,4 % (р<0,05), в 4 сутки на 46,4 % (р<0,05) и в последние сутки исследования на 46,6 % (р<0,05). При питьевой мотивации наблюдалась аналогичная тенденция изменения параметров ПА при ИЭС у лабораторных животных. ВП снизилось в 1 сутки на 13,0 % (р<0,05), во 2 сутки на 22,1 % (р<0,05), в 3 сутки на 24,5 % (р<0,05), в 4 сутки на 25,0 % (р>0,05), в 5 сутки на 24,1 % (р<0,05). ИП уменьшилась в 1 сутки на 24,2 % (р<0,05), во 2 сутки на 29,3 % (р<0,05), в 3 сутки на 24,5 % (р<0,05), в 4 сутки (р>0,05), в 5 сутки на 22,5 % (р<0,05).
КП достоверно возрос в 1 сутки на 41,6 % (р<0,05), во 2 сутки на 38,1 % (р<0,05), в 3 сутки на 26,9 % (р<0,05), в 4 сутки на 23,3 % (р<0,05) и в 5 сутки на 23,8 % (р<0,05). КСОП изменялся также достоверно в лучшую сторону, снизившись в 1 сутки на 17,3 % (р<0,05), во 2 сутки на 25,8 % (р<0,05), в 3 сутки на 24,3 % (р<0,05), в 4 сутки на 34,0 % (р<0,05), в 5 сутки 50,5 % (р<0,05).
Таким образом, полученные результаты показывают высокую биологическую активность пантолизата по отношению к параметрам высшей нервной деятельности у лабораторных крыс.
Методика тестирования лабораторных крыс в модульном устройстве на примере пищевого поведения
В данной экспериментальной серии нами разработана методика изучения психобиологических особенностей в новом модульном устройстве с применением известного ноотропного вещества - пирацетама. В дальнейшем эта экспериментальная разработка использовалась при алгоритмическом скрининге биологических свойств новых веществ.
Для анализа способности достижения пищевого подкрепления использовали разработанное нами модульное устройство. Предлагаемая конструкция (рис. 6) включает в себя стартовый (1) и четыре целевых модуля (2, 3, 4, 5). В боковых cтенках стартового модуля вырезаны по два отверстия: вход (6) и выход (7). Выходы (7) прикрыты дверцами (8), открывающимися только во внутрь (1). Целевые модули (2, 3, 4, 5) идентичной конструкции включают в себя разделительную стенку (9), целевую часть (10), в которой размещается подкрепление, входной тоннель (11) с аверзивным препятствием и выходной коридор (12). Перед входом в тоннель в стартовом модуле (1) размещается педаль (13), ярко окрашенная, соединенная с дверцей (8). При нажатии на педаль (13) происходит открытие затвора дверцы (8).
В стартовый модуль (1) помещается животное. В целевую часть (10) кладется подкрепление. Во входном тоннеле (11) располагается аверзивное препятствие (решетка с импульсным пороговым током силой 0,1 - 0,2 мА) Животное обоняет подкрепление и пытается добраться до него по входному тоннелю (11). Аверзивное препятствие мешает ему. В попытках достичь подкрепления животное методом проб и ошибок (Зорина З.А., Полетаева И.И., 2003) нажимает на педаль (оперантное поведение). Дверца (8) выхода падает во внутрь целевого модуля и открывает доступ к подкреплению. Животное по выходному коридору (12), как по обходному пути, добирается до подкрепления, минуя аверзивное препятствие.









Рисунок 6 - Схема модульного устройства, используемого для изучения способности животных к достижению подкрепления (вид сверху).
При исследовании в модульном устройстве фиксировали следующие показатели: латентный период (время от момента помещения животного в устройство до первой побежки (правильной или ошибочной)); время достижения подкрепления (среднее значение периода от момента первой побежки (правильной или ошибочной) до нахождения подкрепления в последнем модульном отсеке). Общая продолжительность тестирования составляла 600 с. При достижении подкрепления через тоннель с аверзивным раздражителем, побежка расценивалась как ошибочная, через выход, открываемый после нажатия на педаль - правильной и, соответственно, отсутствие побежек в тот или иной выход - как отказ от поиска подкрепления.
В работе использованы три группы крыс: I группа (n=11) - контрольные животные, не подвергающиеся пищевой депривации и содержащиеся в обычном режиме; II группа (n=11) - особи, подверженные трехсуточной пищевой депривации; III группа (n=11) - животные, подверженные трехсуточной депривации пищи и дополнительно получавшие внутрибрюшинно пирацетам (300 мг/кг) за 30 мин до начала эксперимента. Физиологический раствор крысам из I и II исследуемой группы вводился в эквивалентном количестве аналогичным образом.
На предварительном этапе эксперимента, все животные подвергались суточной пищевой депривации, после чего помещались в модульной устройство, в котором все выходы были открыты. Контрольные и экспериментальные особи имели возможность в установке оценить обстановочную афферентацию и воспользоваться любым выходом (без встречи с аверзивным раздражителем) для достижения подкрепления. Такая процедура проходила троекратно с интервалом в 48 часов. После десятидневного перерыва начинался эксперимент, к началу которого животные из экспериментальных групп были подвержены суточному лишению пищи. Тестирование проводилось в течение трех дней в одно и то же время. С каждыми сутками исследования соответственно увеличивалась продолжительность пищевой депривации.
По результатам исследования экспериментальные животные из контрольной группы во все дни тестирования не осуществляли правильного выбора в модульном устройстве (табл. 3). При этом количество отказов от поиска в данной группе с каждым днем увеличивалось наряду с уменьшением числа особей выбирающих неверный выход. Среднее значение латентного периода и времени достижения подкрепления в контрольной выборке во 2 и 3 сутки наблюдения не определялись (табл. 3), т.к. доминирующая часть животных не принимала участия в пищедобывательном поиске.
Таблица 3
Изменение поведенческих параметров в модульном устройстве под влиянием пирацетама на фоне увеличивающейся пищевой мотивации
Показатели |
Группа |
Сутки тестирования |
||
1 сутки |
2 сутки |
3 сутки |
||
атентный период, с |
I |
341,444,3 |
- |
- |
II |
214,238,9 + |
173,427,1 |
152,051,1 |
|
III |
127,223,4 * |
аа 107,329,9 * |
а 81,331,6 |
|
Время достижения подкрепления, с |
I |
187,327,8 |
- |
аа - |
II |
115,731,1 + |
98,318,4 |
а 82,016,6 |
|
III |
94,321,5 + |
62,714,9 * |
44,217,8 * |
|
Примечание. * - p<0,05 по сравнению с II группой; + - p<0,05 по сравнению с контрольной группой
Анализируя полученные данные II группы, отмечалась положительная тенденция к увеличению количества крыс, осуществляющих переход из стартового в целевые модули (через аверзивный раздражитель, либо минуя его), хотя единичные особи вырабатывали ситуативный отказ от поведенческих действий (табл. 4). Поградиентно возрастающая пищевая мотивация в период проведения опыта оказала позитивный эффект на средние значения количественных показателей в II исследуемой группе (табл. 3), где в 1 сутки тестирования имелась достоверно значимая разница (у обоих показателей) с контрольным значением (р<0,05). На протяжении остальных дней исследования фиксировалось уменьшение средних значений наблюдаемых параметров.а
а Таблица 4
Количественное распределение животных по правильности выхода и отказу от поиска из стартового модуля
Сутки тестирования |
Отказались от поиска |
Выбрали неправильный выход |
Выбрали правильный выход |
||||||
I |
II |
III |
I |
II |
III |
I |
II |
||
1 сутки |
7 |
3 |
1 |
4 |
5 |
6 |
- |
3 |
5 |
2 сутки |
9 |
1 |
1 |
2 |
4 |
- |
- |
6 |
10 |
3 сутки |
10 |
1 |
1 |
1 |
3 |
- |
- |
7 |
10 |
III экспериментальная группа имела минимальное количество крыс, не участвующих в пищедобывательном поведении (ежедневно по 1 шт.). В 1 сутки наблюдения практически половина животных совершила неправильную побежку, а во 2 и 3 день наблюдения все особи совершили верные действия (за исключением вышеупомянутой). Средняя величина латентного периода в 1 и 2 сутки имела достоверно значимую разницу в сравнении со II группой и также с I группой в 1 сутки (р<0,05). Аналогичное значение времени достижения подкрепления в 1 сутки исследования достоверно различалось в сравнении с I группой, во 2 и 3 сутки в сравнении со II (р<0,05), снизившись к последнему дню наблюдения более чем в 2 раза.
Отсутствие пищедобывательной деятельности (на 3 сутки) в модульном устройстве у животных контрольной выборки является закономерным и легко объяснимым фактом, связанным с низким уровнем выраженности пищевой потребности. Но при этом определенная часть животных осуществляла переход из стартового модуля к целевому через аверзивный раздражитель к подкреплению. Мы полагаем, что данное явление можно объяснить наличием у крыс определенного изначального памятного следа и главным образом технической особенностью архитектуры применяемого нами модульного устройства.
Увеличение пищевой потребности у особей II экспериментальной группы способствовало снижению количественного распределения животных по правильности выхода и отказу от поиска из стартового модуля. Соответственно, на этом же фоне с дополнительным введением пирацетама у III исследуемой группы выявлялась достоверно значимая связь со сравниваемыми выборками.
В результате нами апробирована принципиально новая методика изучения поведенческих особенностей лабораторных крыс, которую можно использовать при анализе психобиологических особенностей животных, подвергающихся воздействию каких-либо факторов.
Универсальное устройство с изменяемой архитектурой, как инструментальный способ, позволяющий изучать врожденные и приобретенные формы поведения
Целью данной экспериментальной серии явилось разработка устройства для изучения различных форм поведения в условиях разного уровня выраженности пищевой потребности с учетом типологических особенностей.
Разработанное нами устройство (патент РФ № 2332166, БИ 24 от 27.08.08) включает в себя квадратный ящик, заполненный квантами: блоками - параллелепипедами (рис. 7).
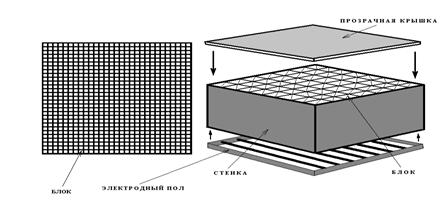
а аа Б
Рисунок а7 - Исходный вид устройства с универсальной изменяющейся архитектурой (А - вид сверху, Б примерный вид в изометрии).

Рисунок 8 - Строение модульного блока, являющегося квантом устройства.
|
Страницы: | 1 | 2 | 3 | 4 | 5 | 6 | |
Авторефераты по всем темам >>
Авторефераты по биологии