
 Авторефераты по всем темам >>
Авторефераты по ветеринарии
Авторефераты по всем темам >>
Авторефераты по ветеринарии
Морфология больших слюнных желез млекопитающих с разным типом питания
Автореферат докторской диссертации по ветеринарии
|
Страницы: | 1 | 2 | 3 | |
 СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА
СКАЧАТЬ ОРИГИНАЛ ДОКУМЕНТА3.2. Видовые особенности электронно-микроскопического строения аоколоушной и нижнечелюстной слюнных желез
Околоушная слюнная железа яка, крупного рогатого скота, китайского водяного оленя, свиньи, собаки и леопарда, наряду с общими закономерностями строения секреторных клеток, имеет существенные видовые различия.
У всех названных видов животных железистые клетки, контактируя между собой, окаймляют центральный проток ацинуса, просвет которого сообщается с секреторными капиллярами, проходящими между боковыми поверхностями граничащих клеток.
У жвачных секреторные канальцы расширенные и ветвистые, у собаки, леопарда и свиньи канальцы расширены только в базо-латеральной области.а В апикальной части плазмолеммы граничащих клеток сближены и образуют замыкающие контакты. В просвет ацинуса и секреторных канальцев обращены ворсинки апикального и латерального участков поверхности эпителиальных клеток концевых отделов. У собаки на апикальной поверхности клеток они короткие и редкие. У леопарда в этой области ворсинки не обнаружены. У свиньи микроворсинки отмечены на базо-латеральной поверхности гландулоцитов. У яка, крупного рогатого скота и китайского водяного оленя ворсинки апикальной, латеральной и базо-латеральной поверхностей клеток многочисленные и длинные. С помощью длинных и многочисленных ворсинок у жвачных осуществляется тесный контакт большой области внутриклеточного пространства и секреторных канальцев, что обеспечивает быстрое проникновение воды сквозь секреторные клетки ацинусов. Наши результаты согласуются с даными ряда авторов (van Lennep E.W. et al., 1977; Kim J.S. et al., 1994; Mansouri S.H., Atri A., 1994). Мы предполагаем, что возможная функция ворсинок заключается в продвижении серозного секрета из просвета концевого отдела во вставочный проток. Многочисленные микроворсинки и глубокие впячивания базальной плазматической мембраны, свидетельствуют о выработке концевыми отделами железы жвачных большого количества ионов. Последние обеспечивают довольно высокую щелочность (рН 8,1) и буферные свойства слюны жвачных, в рационе которых преобладает целлюлоза, что имеет большое значение для нейтрализации кислот, образующихся вследствие деятельности целлюлозолитических бактерий (сбраживание клетчатки и других углеводов).
Ядра гландулоцитов у яка и крупного рогатого скота полигональной формы, образуют много лопастей и располагаются в базальной части клеток. Многолопостная форма ядра увеличивает площадь ее поверхности для ядерно-цитоплазматических взаимодействий и, тем самым, оказывает влияние на скорость метаболических реакций (Свенсон К., Уэбстер П., 1980). У свиньи, собаки, леопарда, и китайского водяного оленя ядро овальной или несколько уплощенной формы и, в зависимости от секреторного цикла, может располагаться в центральной или базальной части клетки.
Железистые клетки околоушной железы яка, крупного рогатого скота содержат многочисленные митохондрии, разветвленные внутриклеточные канальцы и элементы агранулярной эндоплазматической сети. Пластинчатый аппарат развит слабо, а гранулярная эндоплазматическая сеть практически не выявляется. Это согласуется с характеристикой концевых отделов околоушной железы у овец (van Lennep E.W., 1977), коз (Kim J.S. et al., 1994), верблюда (Mansouri S.H., Arti A., 1994). Между тем, як, в отличие от крупного рогатого скота, получает корма с большим количеством клетчатки. Особенностью секреторных клеток околоушной железы у данного вида является наличие глубоких впячиваний базальной плазматической мембраны и мелких единичных секреторных гранул, а также многочисленных митохондрий. В рационе крупного рогатого скота содержится больше переваримого белка (Васильев К.А., 1991). Секреторные клетки околоушной железы этого вида содержат меньше митохондрий и больше секреторных гранул.
Одной из важных характеристик потребляемого корма китайским водяным оленем является высокое содержание лигнинов и таннинов (Tiexier H. et al., 1997), которые известны своей способностью, ингибировать процессы пищеварения, вызывая системную интоксикацию. Клетки ацинусов околоушной железы данного вида характеризуются хорошо развитыми гранулярной эндоплазматической сетью и пластинчатым аппаратом. Они вырабатывают большое количество секреторных гранул, которые, вероятно содержат белки, принимающие участие в нейтрализации танинов и лигнинов. Это является приспособительной реакцией организма, сформированной в процессе эволюционной адаптации. Характерной особенностью некоторых клеток ацинусов в околоушной железе китайского водяного оленя является сжатие ядра большими вакуолями, располагающимися в апикальной части клеток. Подобное явление было отмечено M.Stolte и Ito S. (1996) у косули, антилопы, тара, оленя-лира, благородного оленя, милу, европейского муфлона, африканского буйвола, черной лошадиной антилопы.
Железистые клетки околоушных желез плотоядных животных характеризуются сильным развитием гранулярного эндоплазматического ретикулума и пластинчатого аппарата. Им свойственна полярность, которая проявляется в базальном расположении основной массы цистерн гранулярного эндоплазматического ретикулума и надъядерной локализацией пластинчатого комплекса. Ядра располагаются в средней или базальной части клетки, а секреторные гранулы занимают надъядерную зону.
Гландулоциты околоушной железы свиньи характеризуются базальным расположением уплощенных ядер, сильным развитием пластинчатого аппарата, базально смещенной и слабо развитой гранулярной эндоплазматической сетью, обильным накоплением секрета, заполняющим почти всю клетку. Секреторные гранулы часто сливаются между собой.
Секреторные гранулы в околоушных железах изученных видов млекопитающих характеризуются видовой специфичностью. У яка они округло-овальной формы, мелкие, средней электронной плотности. У крупного рогатого скота секреторные гранулы гомогенные, округлой или овальной формы. Они заключены в электронно-плотную капсулу, различаются по размерам и электронной плотности. Крупные гранулы средней электронной плотности, мелкие характеризуются более высокой электронной плотностью. У китайского водяного оленя секреторный материал представлен округлыми гранулами двух типов - крупными с умеренной электронной плотностью и более мелкими - относительно высокой электронной плотности. Последние иногда содержат располагающиеся по периферии электронно-плотные включения. Мы считаем, что различная плотность секреторных гранул железы зависит от степени их зрелости. Оба типа гранул окружены узким электронно-прозрачным ободком. У свиньи большая часть гранул гомогенна и обладает низкой электронной плотностью. Обычно гранулы окружены мембраной.
Основу гранул другого типа составляет неплотный матрикс, в который вкраплен плотный материал. Секреторный материал в клетках ацинусов околоушной железы собаки представлен в виде округлых электронно-плотных гранул, которые окружены узким электронно-прозрачным ободком. Внутри большей части секреторных гранул содержатся сферические тельца высокой электронной плотности. Каждое тельце располагается чаще эксцентрично или в середине. У еопарда секреторные гранулы заполняют практически всю цитоплазму, в большинстве своем округлой формы, различной электронной плотности и размеров, заключены в электронно-плотную капсулу и окружены электронно-прозрачным ореолом. Среди них встречаются единичные гранулы палочковидной и полулунной формы. Некоторые гранулы содержат мелкие электронно-плотные тельца - включения, окруженные очень тонким ореолом слабой электронной плотности. Тельца располагаются чаще эксцентрично.
В серомукозных клетках нижнечелюстной слюнной железы изученных нами животных секреторные гранулы полиморфны: центральная часть их содержит участки более высокой электронной плотности.
В отличие от ацинарных клеток околоушных желез, зрелые гранулы которых обладают весьма высокой электронной плотностью, в мукоцитах нижнечелюстной железы зрелые секреторные гранулы, расположенные в апикальной части цитоплазмы, также имеют низкую электронную плотность.
Вставочные протоки у некоторых изученных нами видов животных характеризуются наличием микроворсинок на апикальной поверхности эпителиоцитов. Единичные и короткие микроворсинки покрывают апикальную поверхность эпителиальных клеток вставочных протоков околоушной железы собаки и леопарда. Длинные микроворсинки отмечены на апикальной и латеральной поверхности клеток вставочных протоков околоушной железы китайского водяного оленя. В тоже время, у крупного рогатого скота апикальная плазмолемма клеток вставочных протоков одноименной железы микроворсинок не образует.а Местами отмечены небольшие округлой формы выпячивания апикальной плазмолеммы в просвет протока. У яка среди эпителиоцитов вставочных протоков присутствуют клетки, апикальная плазмолемма которых образует листообразные выпячивания в просвет протока.
Степень выраженности латеральных складок эпителиоцитов вставочных протоков неодинакова не только у разных видов, но и варьирует у различных клеток одного вставочного протока. В клетках вставочных протоков околоушной и нижнечелюстной слюнных желез у яка, крупного рогатого скота, свиньи, собаки и леопарда впячивания ав базальной зоне плазматических мембрана не выражены. Апикальные поверхности соседних клеток образуют тесные контакты - десмосомы.
Цитозоль эпителиоцитов вставочных протоков околоушной железы крупного рогатого скота, яка, свиньи, собаки, леопарда, а также нижнечелюстной слюнной железы жвачных, содержит цитофиламенты. Это согласуется с данными W.H. Wilborn, J.M. Shackleford (1969), C.D. Bloom, R. Carlcoo (1974), E.W. van Lennep et all. (1977), J.L. Boshell, W.H. Wilborn (1978).
Эпителиальные выстилки вставочных протоков у разных видов животных существенно различаются по наличию или отсутствию, а также качественному показателю секреторных гранул.
Секреторные гранулы обнаружены нами в эпителиоцитах вставочных протоков околоушной железы свиньи, собаки, а также леопарда и китайского водяного оленя, что согласуется с данными аряда исследователей (Boshell J.L., Wilborn W.H., 1978; Suzuki S. et al., 1975, 1980; Yamada K., 1977). Не выявлены секреторные гранулы в эпителиоцитах протоков околоушной железы крупного рогатого скота, что не противоречит данным J.M. Shackleford, W.H. Wilborn (1969), S. Suzuki et al., (1981e), E.W. van Lennep et al., (1977), E. Raubenheimer (1987).
В цитоплазме эпителиоцитов вставочных протоков присутствуют мелкие вакуоли.
Вставочные отделы переходят в исчерченные протоки. Базальная исчерченность эпителиоцитов исчерченных протоков, видимая в световой микроскоп, обусловлена многочисленными выростами базальной плазмолеммы, между которыми находятся ряды вертикально расположенных митохондрий, что согласуется с данными A. Tamarin,а L.M. Sreebny (1965), В. Tandler (1963, 1993б).
Видовые различия организации базальной части эпителиоцитов исчерченных протоков, сводятся к количеству и глубине складок базальной плазмолеммы, форме и размерам базальных митохондрий. Степень базальной складчатости выражена неодинаково в исчерченных протоках околоушной и нижнечелюстной слюнной железы. Слабо развита складчатость базальной плазмолеммы эпителиоцитов исчерченных протоков околоушной слюнной железы у жвачных, что согласуется с данными S.H. Mansouri, A. Atri (1994).
Поверхность эпителиоцитов, обращенная в просвет протока, варьирует у разных видов млекопитающих. У китайского водяного оленя в околоушной железе она несет нечастые невысокие микроворсинки, на наличие которых у других видов указывает B.Tandler et al. (2001).а У яка, апикальный полюс и цитоплазма эпителиоцитов исчерченных протоков этой железы на границах со смежными клетками образуют выросты, напоминающие листочки книжки многокамерного желудка (рис.3А). Сканирующая микроскопия, в свою очередь, показала, что данные складки-выпячивания возвышаются над апикальной поверхностью, граничащих друг с другом эпителиоцитов, образуя полость шестигранной формы, что придает внутренней поверхности протока вид, напоминающий соты (рис.3Б). По нашему мнению, данные рельефные образования в околоушной слюнной железе яка способствуют удержанию слюны на апикальной поверхности клеток исчерченных протоков. Тем самым они увеличивают функциональную поверхность эпителиоцитов, продлевают время контакта секрета с клетками протоков и облегчают уравновешивание солевого состава слюны и плазмы крови. Чем медленнее скорость движения слюны по вставочным и исчерченным протокам, тем больше времени остается для контакта секрета с эпителиальными покровами, что оказывает влияние на качественный состав слюны.
В апикальной части цитоплазмы клеток исчерченных протоков нами обнаружены секреторные гранулы в околоушной железе собаки, леопарда, рогатого скота и свиньи, что подтверждает данные S. Suzuki et al. (1975), T. Nagato, B. Tandler (1986a) и других. Наличие серозно-подобных гранул в апикальной зоне цитоплазмы эпителиоцитов исчерченных протоков у многих видов млекопитающих свидетельствует о том, что они вовлечены в секрецию органических продуктов.
![]()
![]()
Б
аА
а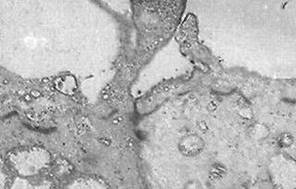 а
а
Рис.3 А - Цитоплазматические выросты апикальной поверхности эпителиоцитов исчерченных протоков околоушной слюнной железы яка. ТЭМ, ув. 4000; Б - як. Исчерченный проток околоушной слюнной железы. SEM, ув.2000.
Помимо гранул, апикальная часть цитоплазмы клеток исчерченных протоков изучаемых нами видов млекопитающих содержит многочисленные мелкие везикулы, которые также встречаются и у других млекопитающих (Hand A.R. 1979, 1987; Sato A., Miyoshi S., 1998; Yamamoto-Hino M. et al., 1998). Функциональное значение везикул в литературе дискутируется.
В эпителии исчерченных протоков выявлено наличие пяти типов клеток. Первый тип клеток характеризуется широкой базальной частью и суженной апикальной, электронно-плотной цитоплазмой, центрально расположенными ядрами полигональной формы (что увеличивает их активную поверхность). Базальная плазмолемма их сильноскладчатая. Многочисленные митохондрии, расположенные между складками цитолеммы, характеризуются большим количеством параллельных крист. В перинуклеарной зоне сосредоточенно умеренное количество митохондрий.
Второй тип клеток характеризуется узкой базальной и расширенной апикальной частью, которая, зачастую, не достигает просвета протока.а Базальная плазмолемма складок не образует. Ядра клеток округлые или округло-овальные, крупные. Цитоплазма клеток электронно-прозрачная, содержит умеренное количество округлых, иногдаа сильно набухших, митохондрий, часть из которых превращается в электронно-прозрачные вакуоли. Особенно крупные вакуоли наблюдаютсяа в базальной части клеток.
Клетки первого и второго типов составляют основную массу эпителиальной выстилки стенки исчерченных протоков.
Третий тип - особые УсветлыеФ клетки - грушевидной формы, узкая часть которых лежит на базальной мембране. В электронно-прозрачной цитоплазме располагаются органеллы и электронно-плотные округлые гранулы. Гранулы содержатсяа как в апикальной, так и в базальной части клеток. Как правило, эти клетки располагаются возле УтемныхФ. Мембраны, выстилающие боковые поверхности смежных УсветлыхФ и УтемныхФ клеток, вместе с цитоплазмой образуют неглубокие складки, соответствующие по форме углублениям соседней клетки.
Четвертый тип клеток представлен так называемымиа УтемнымиФ клетками - узкими, с электронно-плотной цитоплазмой, в которой содержатся органеллы. Ядра клеток узкие, вертикально-вытянутые. Апикальные полюсаа клеток снабженыа мелкими единичными микроворсинками.
Пятый тип - базальные клетки - са электронно-прозрачной цитоплазмой, малым количеством элементов гранулярной эндоплазматической сети и многочисленными полисомами. Митохондрий в клетках мало, и они мелкие. Единичные митохондрии присутствуют в дистальной части эпителиального покрова исчерченных протоков. Наши результаты подтверждают полученные ранее данные об ультаструктуре базальных клеток у крыс, мышей, крупного рогатого скота, овец, свиньи, зубра, европейского благородного оленя, европейской косули, горного козла Урала, кабана, хоря золотистого а(Бабаева А.Г., Шубникова Е.А., 1979; Зеленевский Н.В., 1992; Зеленевский Н.В. с соавт., 1994).а
В экскреторных протоках базальные клетки располагаются в виде сплошной узкой линии.
Миоэпителиальные клетки окружают концевые отделы, вставочные и исчерченные выводные протоки больших слюнных желез млекопитающих.
В отношении расположения миоэпителиальных клеток мнение исследователей противоречиво. Большинство авторов считают, что миоэпителиоциты располагаются между секреторными клетками и базальной мембраной (Погодина Л.С. с соавт., 1974; Фоменкова Т.Е., 1975; Бабаева А.Г., Шубникова Е.А., 1979; Franke et al., 1980; Konig B.Jr., Masuko T.S., 1998). По данным Н.В. Зеленевского (1992), миоэпителиальные клетки располагаются кнаружи от базальной мембраны, а Д.Н.Зеленевский (2007) утверждает, что они находятся между листками базальной мембраны.
По нашим данным, миоэпителиальные клетки располагаются между секреторными клетками и базальной мембраной.
Миоэпителиоцитыа обнаружены нами вокруг серозных и слизистых концевых отделов, во вставочных, и в проксимальной части исчерченных протоков.а Количество миоэпителиальных клеток вокруг концевых отделов зависит от вязкости слюны. В околоушной слюнной железе они располагаются в местах перехода концевых отделов во вставочные протоки, а в нижнечелюстной - они болееа многочисленны вокруг слизистых и смешанных ацинусов, чем серозных.
Среди миоэпителиоцитов присутствуют светлые и темные клетки. Светлые клетки малочисленны и характеризуются большим размером, электронно-прозрачной цитоплазмой, короткими и толстыми отростками, малым количеством неравномерно распределенных миофиламентов. Большинство миоэпителиоцитов представленоа темными клетками с электронно-плотной цитоплазмой, содержащей многочисленные миофиламенты.а Наши результаты не расходятся с даннымиа Е.А. Шубниковой (1980), Y. Nagahsima и K. Ono (1985) о наличии темных и светлых миоэпителиальных клеток в больших слюнных железах млекопитающих. В отношении природы и функционального значения тех и других видов клеток мы склонны придерживаться мнения А.Л. Зашихина и др. (2004)а о высоком уровне дифференциации, как темных, так и светлых миоэпителиоцитов и считаем их зрелыми клетками, находящимися в различной фазе функциональной активности.
3.3. Закономерности пространственной организации волоконных структур соединительной ткани больших слюнных желез млекопитающих по материалам сканирующей электронной микроскопии
В составе единого соединительнотканного остова больших слюнных желез у всех изученных видов млекопитающих присутствуют элементы оболочечного (капсула, оболочки долей, долек, стенки ацинусов), межоболочечного (междолевая, междольковая, межацинозная соединительная ткань),а переходного (паравазальная, парадуктульная, параневральная соединительная ткань) типов, что согласуются с данными О.В.Куваевой (2002). Строение элементов определяется выполняемой ими функцией в составе соединительнотканного остова, а варианты изменчивости - характером взаимоотношений друг с другом и смежными образованиями.
Концевые отделы и протоковая система больших слюнных желез, окруженные миоэпителиальными клетками, окутаны тонкой фибриллярной или фибриллярно-волоконной сетью. Количество фибрилл, их организация, варьируют у изученных видов животных и не одинаковы в различных железах. Между тем, прослеживается определенная закономерность в организации соединительнотканных фибрилл вокруг концевых отделов:
- В железах, где гландулоциты ацинусов вырабатывают жидкий серозный секрет, миоэпителиальные клетки, окружающие концевые отделы, сосредоточены, главным образом, в местах перехода ацинусов во вставочные протоки. Фибриллы, налегая друг на друга в нескольких взаимно перпендикулярных направлениях, образуют узелковое переплетение (рис.4А);
- аВокруг концевых отделов, гландулоциты которых вырабатывают более густой мукозный секрет, миоэпителиальные клетки расположены вокруг концевых отделов. Фибриллы, плотно прилегая друг к другу, образуют тонкие, но широкие волокна с волнистым ходом (рис.4Б).
аа Соединительная ткань вокруг концевых отделов обеспечивает устойчивую силу противодействия сжимающему и растягивающему действию миоэпителиальных клеток во время продвижения секрета, а также упругость при механических движениях, вызванных жеванием или сосанием. Наличие тонкой филаментной сети вокруг ацинусов и протоков больших слюнных подтверждает существование обратной эпителиально- мезенхимальной связи, которая проявляется в способности эпителиальных тканей продуцировать коллаген и влиять на состояние самих фибробластов (Hosoyamada Y., Sakai T., 2003; Разумов В.В., 2006).
Концевые отделы и протоки, покрытые фибриллярной сетью, объединяются пластом компактно расположенных уплощенных и цилиндрических коллагеновых и эластических спирально-волнистых волокон. Основную массу волокнистого остова, окружающего группы ацинусов и протоковую систему составляют уплощенные слегка волнистые
А
аБ
а а
а 
Рис.4 А - фибриллы на поверхности ацинуса околоушной железы свиньи. SEM, ув. 10 000; Б - фибриллы на поверхности ацинуса нижнечелюстной железы свиньи. SEM, ув. 10 000.
б
аа
аБ
аА
а а
а
Рис.5 А - межацинозная соединительная ткань околоушной железы собаки. SEM, ув. 10 000; Б - парадуктальная соединительная ткань околоушной железы яка: а - основные волокна; б - связочные волокна. SEM, ув. 3 000.
коллагеновые волокна, располагающиеся компактно в одной плоскости, параллельно друг к другу вдоль длинной оси либо в разных направлениях или в беспорядке. Волокнистость неодинаково выражена как по длине волокна, так и в разных волокнах. Это позволяет им выдерживать большое растяжение при нагрузке по направлению хода волокна.
Между тонкой фибриллярной сетью иа волоконным пластом существует пространство, в котором иногда наблюдается наличие кровеносных капилляров. Оно может служить упругой подушкой против давления, создаваемого накоплением секреторных гранул и сжатием миоэпителиальных клеток во время секреции. По мнению ряда авторов (Schmalbruch, H. 1974; Ohtani О. et al., 1988; Watanable L. et al., 1997), оноа необходимоа также и для сократительной активности клеток.
Элементы межоболочечного (междолевая, междольковая, межацинозная соединительная ткань) типа представляют переходный остов, имеющий неориентированный тип строения, относительно большие межволоконные пространства и выраженную спиральность волокон цилиндрической формы (рис.5А). Переходный остов выполняет редукторную функцию между соседними структурными единицами с различной механической подвижностью.
В конструкции соединительнотканного остова больших слюнных желез млекопитающих мы выделяем две системы коллагеновых волокон, различных по своему функциональному значению. Первая включает в себя основную массу параллельных коллагеновых волокон, идущих в направлении основных механических напряжений, связанных с накоплением и продвижением секрета (рис. 5Б,а). Вторая система представлена тонкими связочными волокнами, расположенными поперечно по отношению к основным, и выполняет вспомогательную интегрирующую функцию, обеспечивая взаимосвязь основных коллагеновых волокон и других структурных элементов желез, и поддерживает их функциональную архитектонику (Рис. 5Б,б).а Система связочных волокон описана и в других органах (Омельяненко Н.П. и др., 1977; Омельяненко Н.П., 1983, Омельяненко Н.П., Жеребцов Л.Д., 1983).
По нашим данным, в строме всех слюнных желез изученных видов животных присутствуют тканевые базофилы. Это согласуется с данными Н.В.Зеленевскогоа (1992), А.Б.Денисова (2003), Д.Н.Зеленевского (2007).а Мы не отмечали наличие тканевых базофилов в соединительнотканной капсуле. В большем количестве они наблюдаются в строме желез, где вырабатывается слизистый секрет. Тканевые базофилы располагаются преимущественно периваскулярно, но часть из них встречается и в межсосудистых участках соединительной ткани.
У представителей плотоядных и всеядных животных бросается в глаза обилие в железах плазматических клеток, что согласуется с данными А.Г.Бабаевой, Е.А.Шубниковой (1979), Н.В.Зеленевского (1992), в несколько меньшем количестве они присутствуют у представителей жвачных. Плазматические клетки располагаются как небольшими группами, так и раздельно, периваскулярно или в соединительной ткани.
В строме слюнных желез нами отмечено наличие лимфоцитов, особенно в большом количестве у всеядных и плотоядных животных. Лимфоцитам придается большое значение в иммуногенезе (Marsh M.N., 1975; Зуфаров К.А. с соавт., 1979).
Локализация лимфоцитов не ограничивается стромой желез, они находятся и внутриэпителиально.а У всеха изученных видов животных нами отмечены два типа внутриэпителиальных лимфоцитов. Первый тип клеток характеризуется эксцентрично расположенным ядром и широкой зоной цитоплазмы. Нуклеоплазма содержит довольно плотно располагающиеся зерна гетерохроматина под внутренней ядерной мембраной. Поверхность клеток неровная в виду наличия небольших углублений и выступов плазмолеммы. Цитоплазма содержит значительное число параллельных пластинок зернистой эндоплазматической сети, богата свободными рибосомами и полисомами. Для второго типа внутриэпителиальных лимфоцитов характерно высокое ядерно-протоплазматическое отношение. Ядро клеток крупное округло-овальное содержит большие скопления глыбок гетерохроматина под внутренней мембраной. Узкая цитоплазма богата свободными рибосомами и полисомами, содержит короткие единичные пластинки гранулярного эндоплазматического ретикулума и мелкие овальные митохондрии. Слабые выступы плазмолеммы придают поверхности клеток неровный вид. Данная морфология лимфоцитов соответствует описанию иммуноглобулиннесущиха лимфоцитов клеток крови и костного мозга (Зуфаров К.А., Тухтаев К.Р., 1987).
Лимфоциты, находясь в эпителии и мигрируя периодически в просвет и строму, обеспечивают передачу информации об антигенных структурах, которые попадают на поверхность эпителия или проникают в него (Аруин Л.И., 1981). Они относятся к диффузной лимфоиднойа ткани (Зуфаров К.А., Тухтаев К.Р., 1987), которая входит в состав единойа иммунной системы слизистых оболочек (Bienenstock J., 1974).
|
Страницы: | 1 | 2 | 3 | |
Авторефераты по всем темам >>
Авторефераты по ветеринарии