Книга вторая Дж. Эдвард Морган-мл. Мэгид С. Михаил Перевод с английского
| Вид материала | Книга |
- Книга первая Дж. Эдвард Морган-мл. Мэгид С. Михаил Перевод с английского, 10010.77kb.
- А. Конан-Дойль новоеоткровени е перевод с английского Йога Рàманантáты, 2314.23kb.
- Copyright Сергей Александровский, перевод с английского Email: navegante[a]rambler, 619.61kb.
- "книга непрестанности осириса " 177, 7373.41kb.
- Н. М. Макарова Перевод с английского и редакция, 4147.65kb.
- Трудового Красного Знамени гупп детская книга, 2911.61kb.
- Трудового Красного Знамени гупп детская книга, 2911.77kb.
- Уайнхолд Б., Уайнхолд Дж. У 67 Освобождение от созависимости / Перевод с английского, 11462.2kb.
- Малиновской Софьи Борисовны Специальность: журналистика Специализация: художественный, 969.08kb.
- Духовные истины в психических явлениях перевод с английского 3-е издание Москва «Философская, 1557.75kb.
Клиническая
Анестезиология
книга вторая
Дж. Эдвард Морган-мл. Мэгид С. Михаил
Перевод с английского под редакцией
академика PAMH А. А. Бунятяна,
канд. мед. наук A. M. Цейтлина
Издательство БИНОМ
Москва
Невский Диалект
Санкт-Петербург
2000
УДК 616-089.5 ББК Р451 М79
Перевод с английского: канд. мед. наук Горелов В. Г., Добродеев А. С., канд. мед. наук Селезнев M. H., канд. мед. наук Цейтлин A. M., Шатворян Б. P.
Дж. ^ Эдвард Морган-мл., Мэгид С. Михаил
М79 Клиническая анестезиология: книга 2-я.— Пер. с англ. — M.-СПб.: Издательство БРШОМ-Невский Диалект, 2000. 366 с., ил.
В книге рассмотрены физиологические основы проведения анестезии у пациентов с сопутствующими заболеваниями сердечно-сосудистой системы, органов дыхания, нервными и психическими расстройствами, нарушениями водно-электролитного баланса и кислотно-основного состояния. В отдельных главах представлены методы проведения анестезиологического пособия в пред-, интра- и постоперационном периодах при хирургических вмешательствах на сердце и сосудах, легких и трахее, пищеводе, головном и спинном мозге и позвоночнике, почках и других органах мочевыделительной системы. Детально освещены вопросы проведения инфузионной терапии — показания, методы, виды растворов, осложнения и альтернативные варианты.
Для врачей-анестезиологов, реаниматологов, студентов медицинских учебных заведений.
Все права защищены. Никакая часть этой книги не может быть воспроизведена в любой форме или любыми средствами, электронными или механическими, включая фотографирование, магнитную запись или иные средства копирования или сохранения информации, без письменного разрешения издательства.
^ ISBN 5-7989-0165-3 (Издательство БИНОМ) ISBN 5-7940-0044-9 (Невский Диалект) ISBN 0-8385-1470-7 (англ.)
Издание на русском языке: © Издательство БИНОМ, Невский Диалект, перевод, оформление, 2000.
Original edition copyright
© 1996, All Rights Reserved.
Published by arrangement with the Original Publisher,
Appleton & Lange a Simon & Schuster Company
Раздел IV
Анестезиологическое
пособие
Физиология кровообращения л q
и анестезия
Анестезиолог должен иметь фундаментальные знания по физиологии кровообращения, которые необходимы как для понимания научных основ специальности, так и для практической работы. В этой главе обсуждаются вопросы физиологии сердца и большого круга кровообращения, а также патофизиологии сердечной недостаточности. Малый (легочный) круг кровообращения рассматривается в главе 22, физиология крови и обмен веществ — в главе 28.
Система кровообращения состоит из сердца и кровеносных сосудов. Она предназначена для снабжения тканей кислородом и питательными веществами и удаления продуктов метаболизма. Сердце перекачивает кровь через две сосудистые системы. В малом круге кровообращения кровь обогащается кислородом и избавляется от углекислого газа. В большом круге — доставляет кислород к тканям и поглощает продукты метаболизма, которые затем элиминируются через легкие, почки или печень.
Сердце
Анатомически сердце представляет собой единый орган, но функционально оно делится на правый и левый отделы, каждый из которых состоит из предсердия и желудочка. Предсердия служат как проводниками для крови, так и вспомогательными насосами для заполнения желудочков. Желудочки выполняют роль главных насосов, перека-
чивающих кровь. Правый желудочек получает дезоксигенированную кровь из большого круга кровообращения и перекачивает ее в малый круг. Левый желудочек получает оксигенированную кровь из малого круга кровообращения и перекачивает ее в большой круг. Четыре клапана обеспечивают однонаправленный поток крови через каждую камеру. Насосная функция сердца обеспечивается сложной последовательностью электрических и механических явлений.
Сердце состоит из специализированной попе-речнополосатой мышечной ткани, заключенной в соединительнотканный каркас. Клетки сердечной мышцы — кардиомиоциты — подразделяются на предсердные, желудочковые, водителей ритма и проводящей системы. Способность кардиомиоцитов к самовозбуждению и их уникальная организация позволяют сердцу функционировать как высокоэффективному насосу. Последовательные соединения между отдельными кардиомиоцитами (вставочные диски), имея низкое сопротивление, обеспечивают быстрое и упорядоченное распространение электрического импульса в каждой камере сердца. Волна возбуждения распространяется от одного предсердия к другому и от одного желудочка к другому по проводящим путям. Связь между предсердиями и желудочками осуществляется не непосредственно, а через АВ-узел, поэтому возбуждение передается с задержкой. За счет этого происходит наполнение желудочка при сокращении предсердия.
^ Потенциалы действия кардиомиоцитов
Мембрана кардиомиоцита проницаема для ионов К4, но относительно непроницаема для ионов Na'. Мембраносвязанная Ка+/К4-зависимая АТФ-аза перекачивает ионы K+ внутрь клетки, а ионы Na" из клетки (глава 28). Концентрация К4 внутри клетки выше, чем во внеклеточном пространстве. Концентрация Na', наоборот, выше во внеклеточном пространстве, чем внутри клетки. Относительная непроницаемость мембраны для кальция поддерживает высокий градиент концентрации кальция между внеклеточным пространством и цитоплазмой. Выход K+из клетки по градиенту концентрации приводит к потере суммарного положительного заряда внутри клетки. Анионы не сопровождают ионы К4, поэтому возникает электрический потенциал, причем внутренняя поверхность клеточной мембраны заряжается отрицательно по отношению к наружной. Таким образом, мембранный потенциал покоя формируется в условиях равновесия между двумя противоположными силами: движением K+ по градиенту концентрации и электрическим притяжением отрицательно заряженным внутриклеточным пространством положительно заряженных ионов К".
В норме мембранный потенциал покоя кардиомиоцита желудочка варьируется от -80 до -90 мВ. Если мембранный потенциал становится менее отрицательным и достигает пороговой величины, то в кардиомиоците, как и в клетках других возбудимых тканей (нерв, скелетная мышца), возникает потенциал действия, т. е. происходит деполяризация (рис. 19-1 и табл. 19-1). Потенциал действия вызывает преходящее увеличение мембранного потенциала кардиомиоцита до +20 мВ. В отличие
от потенциала действия нейрона (гл. 14), в потенциале действия кардиомиоцита за пиком следует фаза плато, которая длится 0,2-0,3 с. Потенциал действия скелетной мышцы и нерва обусловлен лавинообразным открытием быстрых натриевых каналов мембраны, потенциал действия кардиомиоцита вызывается открытием как быстрых натриевых каналов (фаза начальной быстрой репо-ляризации), так и медленных кальциевых каналов (фаза плато). Кроме того, деполяризация сопровождается преходящим уменьшением проницаемости мембраны для калия. В последующем проницаемость мембраны для калия восстанавливается, натриевые и кальциевые каналы закрываются и мембранный потенциал возвращается к исходному уровню.
После деполяризации клетки рефрактерны (невосприимчивы) к деполяризующим стимулам до наступления 4-й фазы. Эффективный рефрак-терный период равен минимальному интервалу между двумя импульсами, вызвавшими распространение возбуждения. В быстропроводящих кар-диомиоцитах эффективный рефракторный период практически равен продолжительности потенциала действия. В медленнопроводящих кардиомио-цитах, напротив, эффективный рефрактерный период может превышать продолжительность потенциала действия.
^ Возникновение и распространение сердечного импульса
В норме электрические импульсы в сердце вырабатываются синусовым узлом — группой специализированных клеток, расположенных в стенке правого предсердия впереди от отверстия верхней полой вены. Мембрана этих клеток характеризуется повышенной проницаемостью для натрия и,
^ ТАБЛИЦА 19-1. Потенциал действия кардиомиоцита
| Фаза | Название | Событие | Движение ионов |
| О | Деполяризация | Активация (открытие) быстрых Na^-каналов и снижение проницаемости мембраны для Ю | Na^ поступает в клетку |
| 1 | Начальная быстрая реполяризация | Инактивация Na'-каналов и преходящее увеличение проницаемости мембраны для К4 | К* выходит из клетки |
| 2 | Плато | Активация медленных Са2+-каналов | Ca2+ поступает в клетку |
| 3 | Конечная быстрая реполяризация | Инактивация Са2+-каналов и увеличение проницаемости мембраны для K+ | K+ выходит из клетки |
| 4 | Потенциал покоя или | Восстановление исходной проницаемости мембраны кардиомиоцитов предсердий и желудочков | К* выходит из клетки |
| | Диастолическая деполяризация | Медленный ток Na+ и, возможно, Ca2+ внутрь клеток, приводящий к спонтанной деполяризации | Na+ поступает в клетку ? Ca2+ поступает в клетку |
возможно, кальция. Медленный ток натрия приводит к снижению потенциала покоя клеток синусового узла (-50— -60 мВ против -80— -90 мВ в кардиомиоцитах желудочков) и имеет три важных следствия: постоянную инактивацию быстрых натриевых каналов; потенциал действия
с порогом -40 мВ, обусловленный в первую очередь движением ионов через медленные кальциевые каналы; регулярную спонтанную деполяризацию. В диастолу поступление натрия в клетку приводит к тому, что мембрана клетки постепенно становится все менее отрицательной; когда дости-
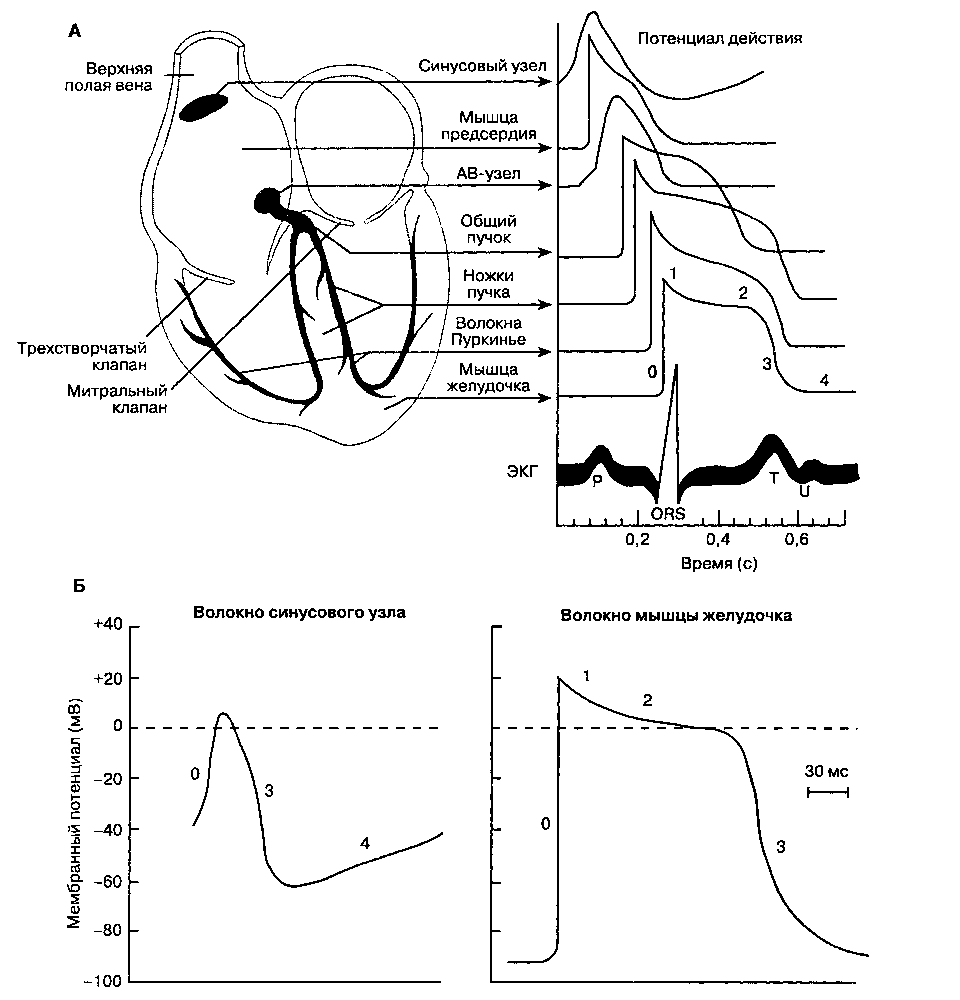
Рис. 19-1. Потенциалы действия сердца. А. Характеристика потенциалов действия различных отделов сердца. Б. Клетки пейсмекера синусового узла характеризуются отчетливой спонтанной диастолической деполяризацией, в них значительно слабее выражены четкие фазы потенциала действия, в отличие от клеток миокарда предсердия или желудочка. Смотрите табл. 19-1 для объяснения различных фаз потенциала действия. (С изменениями. Из: Ganong W. F. Review of Medical Physiology, 16th ed. Appleton & Lange, 1993.)
гается пороговый потенциал, то открываются кальциевые каналы, уменьшается проницаемость мембраны для калия и развивается потенциал действия. Восстановление нормальной проницаемости для калия возвращает клетки синусового узла в состояние покоя.
Импульс, возникший в синусовом узле, в нормальных условиях быстро распространяется по предсердиям pi к АВ-узлу. Специализированные волокна проводят возбуждение как к левому предсердию, так и к АВ-узлу. АВ-узел расположен с правой стороны межпредсердной перегородки, впереди от отверстия венечного синуса и над перегородочной створкой трехстворчатого клапана. В АВ-узле выделяют три отдельные области: верхнюю, среднюю и нижнюю. Средняя область АВ-уз-ла не обладает внутренней спонтанной активностью (автоматизмом), в то время как верхняя и нижняя области способны вырабатывать импульсы. В физиологических условиях водителем ритма является синусовый узел, потому что частота его спонтанной диастолической деполяризации выше, чем в верхней и нижней областях АВ-узла, где она составляет 40-60 мин"1. Любой фактор, уменьшающий частоту деполяризации синусового узла или увеличивающий автоматизм верхней и нижней областей АВ-узла, способствует возникновению АВ-узлового ритма.
Импульсы из синусового узла в норме достигают АВ-узла через 0,04 с и покидают его через последующие 0,11 с. Эта задержка связана с низкой скоростью проведения возбуждения в тонких волокнах внутри АВ-узла, что в свою очередь определяется активацией медленных кальциевых каналов. Напротив, проведение импульса между примыкающими друг к другу клетками в предсердиях и желудочках определяется активацией и инактивацией быстрых натриевых каналов. Волокна, отходящие от нижней части АВ-узла, образуют пучок Гиса. Эта специализированная группа волокон проходит в межжелудочковую перегородку, а затем разделяется на левую и правую ножки, разветвляющиеся в сложную сеть волокон Пуркинье, которые деполяризуют оба желудочка. Волокна Гиса-Пуркинье разительно отличаются от ткани АВ-узла: они имеют наиболее высокую скорость проведения в сердце, что приводит к почти одновременной деполяризации эндокарда обоих желудочков (в норме это занимает 0,03 с). Распространение возбуждения от эндокарда к эпикарду через миокард требует еще 0,03 с. Таким образом, импульсу, возникающему в синусовом узле, в нормальных условиях необходимо менее 0,2 с для деполяризации всего сердца.
Галотан, энфлюран и изофлюран угнетают автоматизм синусового узла. С другой стороны, данные препараты оказывают лишь весьма умеренное непосредственное влияние на АВ-узел, увеличивая время проведения возбуждения и рефрактерность. Сочетание этих эффектов позволяет объяснить высокий риск возникновения AB-узловой тахикардии в тех случаях, когда на фоне ингаляционной анестезии для лечения синусовой брадикардии применяют холиноблокаторы: скорость водителей ритма в АВ-узле возрастает в большей степени, чем таковая в синусовом узле. Электрофизиологические эффекты ингаляционных анестетиков на волокна Пуркинье и миокард желудочков имеют сложный характер из-за участия в процессе вегетативной нервной системы. Описаны как антиаритмические, так и аритмогенные свойства анестетиков. Первые могут быть обусловлены прямым подавлением потока Ca2+ внутрь клетки, вторые — потенцированием действия катехоламинов (гл. 7). Для возникновения аритмогенного эффекта необходима активация как Q1-, так и (3-адренорецепторов.
Токсическая концентрация местных анестетиков в крови сопряжена с риском развития электрофизиологических нарушений в сердце. Лидокаин способен оказывать антиаритмическое действие при низкой концентрации в крови (гл. 48). При высокой концентрации в крови местные анестети-ku подавляют проведение возбуждения в сердце, связываясь с быстрыми натриевыми каналами. Самые мощные местные анестетики — бупивакаин и, в меньшей степени, этидокаин и ропивакаин, по-видимому, оказывают наиболее сильное влияние на сердце, особенно на волокна Пуркинье и миокард желудочков. Бупивакаин связывается с инак-тивированными быстрыми натриевыми каналами (и затем медленно диссоциирует), что приводит к возникновению тяжелой синусовой брадикардии, остановки синусового узла, а также опасной для жизни желудочковой аритмии.
Антагонисты кальция представляют собой органические химические соединения, которые блокируют ток кальция внутрь клетки через медленные каналы. Антагонисты кальция из группы дигидро-пиридинов (нифедипин) перекрывают канал, а вера-памил и, в меньшей степени, дилтиазем связываются с деполяризованным инактивированным каналом.
^ Механизм сокращения
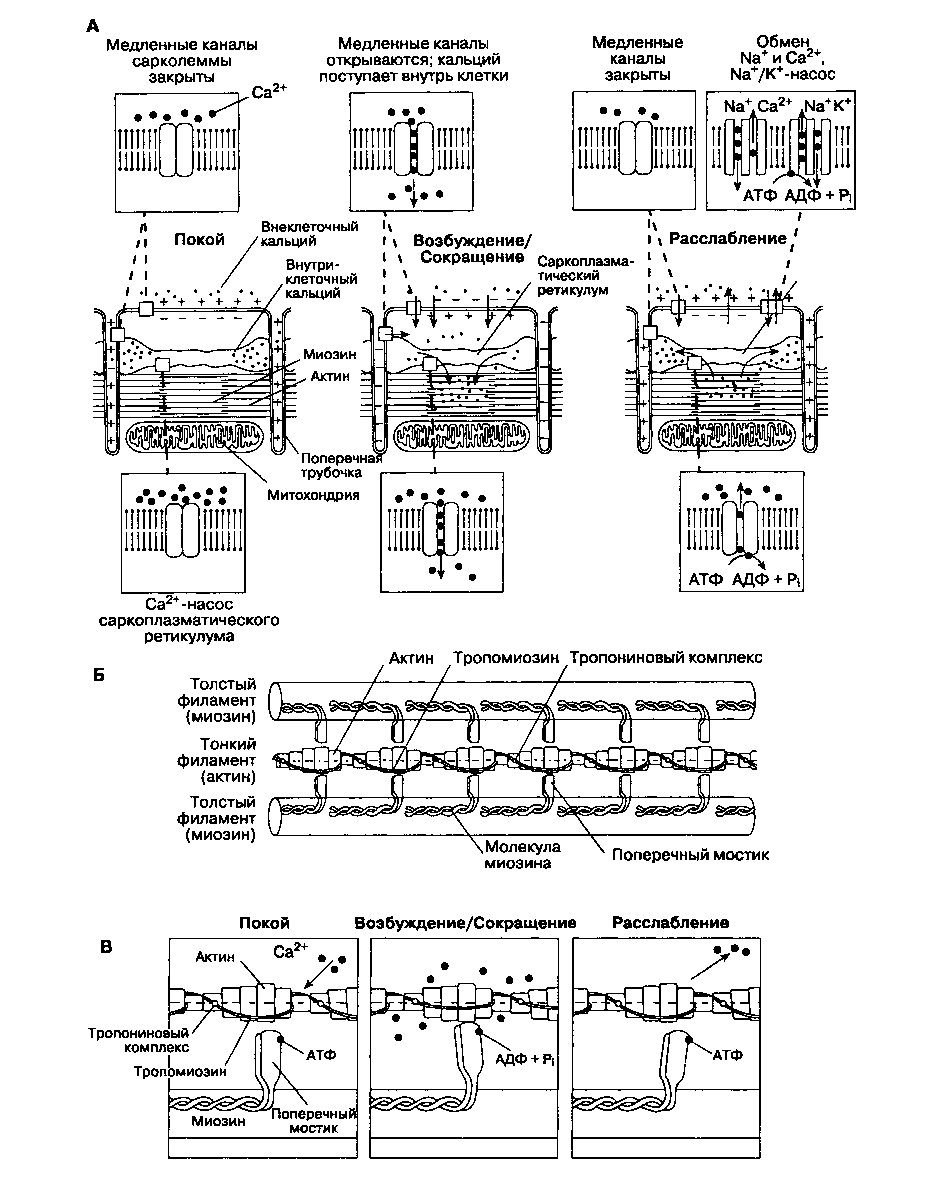
Клетки миокарда сокращаются в результате взаимодействия двух сократительных белков — актина и миозина. Эти белки фиксированы внутри клетки как во время сокращения, так и при рас-
слаблении. Сокращение клетки происходит, когда актин и миозин взаимодействуют и скользят друг относительно друга (рис. 19-2). Это взаимодействие в норме предотвращается двумя регулирующими белками: тропонином и тропомиозином. Молекулы тропонина присоединены к молекулам актина на одинаковом расстоянии друг от друга. Тропомиозин располагается в центре актиновых структур. Увеличение концентрации внутриклеточного кальция (приблизительно с 10~7 до 10~5 моль/л) приводит к сокращению, поскольку ионы кальция связывают тропонин. Кальций изменяет конформацию тропонина, что обеспечивает открытие активных участков у молекул актина, способных взаимодействовать с миозиновыми мостиками. Активные участки на миозине функционируют как Mg-зависимая АТФ-аза, чья активность возрастает при увеличении концентрации кальция внутри клетки. Мостик миозина последовательно соединяется и разъединяется с новым активным участком актина. При каждом соединении потребляет ся АТФ.
^ Сопряжение возбуждения с сокращением
Количество кальция, необходимое для запуска механизма сокращения, превышает таковое, попадающее в клетку через медленные каналы во время 2-й фазы потенциала действия. То небольшое количество кальция, которое входит в клетку (через медленные каналы и механизм двунаправленного ЫауСа2+-трансмембранного обмена), запускает процесс высвобождения гораздо большего количества кальция, хранящегося в клетке (кальцийзависимое высвобождение кальция). Запасы связанного кальция находятся внутри цистерн саркоплазматического ретикулума и, в меньшем количестве, в Т-трубочках. Сила сокращения прямо пропорциональна величине начального потока кальция внутрь клетки. Во время расслабления, когда медленные каналы закрыты, внутриклеточный кальций снова поглощается саркоплазматическим ретикулумом; это активный процесс, требующий участия АТФ-азы мембраны (фосфоламбан). Кроме того, кальций удаляется из клетки посредством трансмембранного обмена внутриклеточного кальция на внеклеточный натрий, а также с помощью активного транспорта АТФ-азой мембраны. Таким образом, процесс расслабления сердечной мышцы также требует АТФ.
^ Количество доступного внутриклеточного Ca2+, скорость его доставки и удаления определяют, соответственно, максимальное развиваемое напряжение, скорость сокращения и скорость расслабле-
ния. Симпатическая стимуляция увеличивает силу сокращения: катехоламины, взаимодействуя с Pi-адренорецепторами, изменяют активность стимулирующего G-белка (глава 18), что вызывает увеличение внутриклеточной концентрации цАМФ (глава 12), а это, в свою очередь, повышает концентрацию внутриклеточного Ca2+. Увеличение внутриклеточной концентрации цАМФ приводит к рекрутированию дополнительных открытых кальциевых каналов. Более того, адреномиметики ускоряют расслабление, потенцируя эффекты фосфоламбана. Ингибиторы фос-фодиэстеразы (теофиллин и амринон), предотвращая метаболизм внутриклеточного цАМФ, действуют аналогично. Дигиталис повышает внутриклеточную концентрацию кальция, ингибируя мембраносвязанную Ка+/К+-зависимую АТФ-азу; в результате происходит незначительное увеличение внутриклеточной концентрации Na+, что способствует прохождению через механизм трансмембранного натрий-кальциевого обмена большего количества Ca2+. Глюкагон, активируя специфические норадренергические рецепторы, увеличивает сократимость за счет повышения уровня внутриклеточного цАМФ.
Высвобождение ацетилхолина, сопряженное со стимуляцией блуждающего нерва, наоборот, снижает сократимость, увеличивая уровень цГМФ и ингибируя аденилатциклазу: эти эффекты опосредованы ингибирующим G-белком. При ацидозе блокируются медленные кальциевые каналы и, соответственно, подавляется сократимость сердца из-за неблагоприятного влияния на внутриклеточную кинетику кальция. Предполагают, что ингаляционные анестетики подавляют сократимость, уменьшая поступление Ca2" в клетки во время деполяризации, изменяя кинетику его высвобождения и поглощения саркоплазматическим ретикулумом, а также снижая чувствительность сократительных белков к кальцию. По силе воздействия на кинетику кальция ингаляционные анестетики располагаются в следующем порядке: галотан > энфлюран > изофлюран > закись азота. Механизм прямого угнетающего действия неингаляционных анестетиков на сократимость миокарда до конца не изучен, но предполагают, что он аналогичен описанному выше.
^ Иннервация сердца
Парасимпатические волокна иннервируют главным образом предсердия и проводящую систему. Ацетилхолин взаимодействует со специализированными м-холинорецепторами сердца (M2), что