
Л. Г. Наумова Б. М. Миркин Основы общей экологии Б. М. Миркин, Л. Г. Наумова Основы общей экологии. Учебник
| Вид материала | Учебник |
СодержаниеГлава 3. Основные принципы аутэкологии 3.1. Принцип экологического оптимума 3.2. Принцип индивидуальности экологии видов 3.3. Принцип лимитирующих факторов |
- «Болото как экосистема», 301.19kb.
- Учебно-методический комплекс по дисциплине ен. Ф. 06. Основы экологии Для специальности, 3772.08kb.
- Р. А. Наумова городского округа город Буй Костромской области программа, 122.49kb.
- Ю. В. Олейников Философско-методологические основы экологического знания, 76.92kb.
- Недостаток кислорода: миф или реальность, 337.03kb.
- Рабочая программа по общей экологии 9 класс Пояснительная записка, 270.09kb.
- Н. Э. Баумана Научно-образовательный центр «Инновационная педагогика в техническом, 397.06kb.
- Задачи курса: - овладение основами экологии; - формирование научных, 101.29kb.
- Экология как научная, 94.09kb.
- Ю. А. Александров Основы радиационной экологии Учебное пособие, 5090.11kb.
Глава 3. Основные принципы аутэкологии
В любой таксономической группе (среди насекомых, ракообразных, мхов, голосеменных и др.) виды разнообразны по требованиям к условиям среды и способности переносить неблагоприятные условия. Тем не менее, есть несколько экологических правил – принципов аутэкологии, которые проявляются в любой группе, хотя из этих правил всегда возможны исключения.
^
3.1. Принцип экологического оптимума
На градиенте любого экологического фактора распространение вида ограничено пределами толерантности (рис. 4). Между этими пределами есть отрезок, на котором условия для конкретного вида наиболее благоприятны и потому формируется самая большая биомасса и высокая плотность популяции. Это его экологический оптимум. Слева и справа от оптимума условия для жизни вида менее благоприятны. Это зоны пессимума, т.е. угнетения организмов, когда падает плотность популяции и вид становится наиболее уязвимыми к действию неблагоприятных экологических факторов (включая и влияние человека).
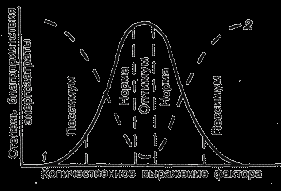
Рис. 4. Схема распределения вида на градиенте среды (по Шилову, 1998). 1 – степень благоприятствования условий среды для организма, 2 – величина энергозатрат на поддержание жизнедеятельности.
В зоне оптимума жизнь организма наиболее комфортна, и он затрачивает на ее поддержание минимальное количество энергии. В зонах пессимума для поддержания жизнедеятельности приходится тратить энергии много больше и включать специальные «механизмы выживания». Например для согревания в холоде теплокровные животные затрачивают энергию, накопленную в жировых тканях. Растения в условиях пессимума затрачивают большую часть продуктов фотосинтеза на дыхание и медленно растут.
Симметричная колоколовидная кривая, показанная на рисунке 4, является достаточно схематизированным приближением к реальной картине отношений видов к факторам среды. Часто распределения видов имеют асимметричный характер, т.е. кривая распределения более пологая с одной стороны и более крутая – с другой. Кроме того, иногда распределения могут быть двухвершинными или даже многовершинными, так как на некоторых отрезках градиента вид может быть подавлен более сильными конкурентами.
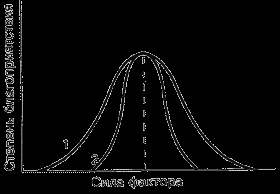
Рис. 5. Варианты отношения видов к факторам среды (по Шилову, 1998). 1 – эврибионтный вид, 2 – стенобионтный вид.
Разные виды имеют различные амплитуды распределения по градиентам факторов среды. Виды с узкой амплитудой называются стенобионтными, с широкой – эврибионтными (рис. 5). Так стенотермные и эвритермные организмы – это виды соответственно неутойчивые и устойчивые к колебаниям температуры. К эвритермным относятся многие насекомые (муравьи, жуки ксилофаги и др.). Пример стенотермности в животном мире – веслоногий рачок (Copilla mirabile), который не выдерживает изменений температуры за пределами 23–29°C (Шилов, 1998). Стенотермными являются деревья тропического леса, которые выдерживают колебания температуры в интервале 5–8°C. Классический пример эвритермности – Лиственница Гмелина (Larix gmelinii), которая в Якутии выдерживает колебания температуры от плюс 30 до минус 70 градусов. Большинство видов деревьев средней полосы Европы выдерживает колебания температуры в пределах 60 градусов. Эта эвритермность во многом связана со способностью деревьев переживать холодное время года в состоянии покоя.
Большой опыт изучения отношения видов к разным факторам среды накоплен ботаниками. Они определяли диапазоны распределения видов по отношению к увлажнению, богатству почвы элементами минерального питания, интенсивности пастбищной нагрузки и т.д. с целью последующей оценки характера экотопов по составу видов растительных сообществ.
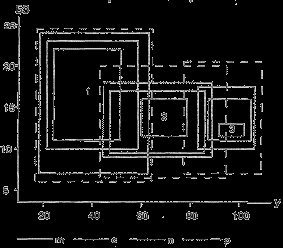
Рис. 6. Экологические ареалы трех видов растений по отношению к факторам увлажнения (У) и богатства почвы элементами минерального питания (БЗ, по Раменскому и др., 1956). Градации проективного покрытия: m – более 8%, c – 8–2,5%, n – 2,4–3 %, p – 0,2–0,1 %.
На рис. 6 показаны экологические ареалы трех видов растений разной экологии, связанных с сухими местообитаниями – типчака (Festuca valesiaca, 1), условиями умеренного увлажнения – овсяницы луговой (Festuca pratensis, 2) и переувлажненными почвами – осоки лисьей (Carex vulpina, 3). Рисунок иллюстрирует общеизвестные экологические закономерности:
– при уменьшении обилия расширяется экологическая амплитуда вида;
– кривые распределения видов по градиентам факторов среды могут налагаться друг на друга при невысоком обилии, но оказываются разомкнутыми при высоком обилии.
Контрольные вопросы
1. На какие градации делится градиент фактора среды между пределами толерантности вида?
2. Какие формы может иметь кривая распределения вида на градиенте экологического фактора, кроме колоколовидной?
3. Приведите примеры стенобионтных и эврибионтных видов.
^
3.2. Принцип индивидуальности экологии видов
Каждый вид индивидуально распределяется по экологическим факторам, кривые распределений разных видов перекрываются, но их оптимумы различаются.
По этой причине при изменении условий среды в пространстве (например,от сухой вершины холма к влажному логу) или во времени (при пересыхании озера, при усилении выпаса, при зарастании скал) состав экосистем изменяется постепенно. Известный российский эколог Л.Г. Раменский сформулировал этот закон образно: «Виды – это не рота солдат, марширующих в ногу».
Индивидуальность экологии видов – это то свойство, которое «крайне неудобно» для практического использования данных о связи видов и условий среды. По этой причине для удобства использования в практике (т.е. из прагматических соображений) знание об индивидуальных особенностях видов упрощается (редуцируется) до системы экологических групп. Такие группы объединяют не тождественные по экологии виды, а виды с близким отношением к одному или нескольким факторам среды.
Для выделения экологических групп градиент среды разбивается на некоторое число классов, и к одной экологической группе относятся виды, которые имеют зоны оптимума в этом классе. Количество классов зависит от задачи, которая была поставлена при выделении экологических групп, и полноты знаний о распределении видов по градиенту экологического фактора, и потому экологические группы видов могут быть выделены в более крупном и более мелком масштабе.
Так например,по отношению к фактору увлажнения почвы все виды растений можно разбить на три группы:
1. ксерофиты – виды сухих местообитаний;
2. мезофиты – виды нормально увлажненных местообитаний;
3. гидрофиты – виды переувлажненных местообитаний.
Однако этот ряд может быть разбит на 5 градаций за счет двух дополнительных переходных групп и тогда примет следующий вид:
1. ксерофиты,
2. ксеромезофиты,
3. мезофиты,
4. мезогидрофиты,
5. гидрофиты.
При наличии более обширного материала о распределении видов вдоль градиентов среды этот ряд может быть увеличен до 7 групп за счет дополнительных переходных групп между мезофитами и гидрофитами. Кроме того, наряду с гидрофитами (типичными водными растениями), можно выделить группу гигрофитов – растений переувлажненных почв, которые покрыты водой лишь часть вегетационного сезона или не заливаются вообще, а увлажнены только за счет приближенных к поверхности грунтовых вод.
Задача выделения экологических групп имеет множество аналогий в практической жизни. К примеру, форма и размер стопы человека индивидуальны, тем не менее, из соображений удобства обувь шьется по системе размеров. Поскольку разделение любой совокупности индивидуальных объектов на группы противоречит их природе, периодически возникают ситуации, когда трудно определить принадлежность объекта к группе: один размер обуви еще мал, а следующий – уже велик. Условность отнесения видов к экологическим группам еще больше. Решить вопрос о том, к какой группе относится вид – ксеромезофитам или мезоксерофитам, гигромезофитам или мезогигрофитам – часто бывает очень сложно.
Важным следствием принципа индивидуальности экологии видов является постепенность изменения состава растительных сообществ и экосистем вдоль градиентов среды. Такие постепенные изменения называются континуумом (непрерывностью). По этой причине конкретные сообщества и экосистемы выделяются также условно, как и экологические группы видов.
Концепция континуума была сформулирована в начале ХХ в. независимо двумя учеными – россиянином Л.Г. Раменским и американцем Г. Глизоном (H. Gleason). Во второй половине ХХ в. наибольший вклад в ее развитие внесли Р. Уиттекер (R. Whittaker), Дж. Кертис (J. Curtis), Р. Макинтош (R. McIntosh) и М. Остин (M. Austin).
Различают континуумы двух типов: экоклин и экотон. Экоклин – это абсолютный континуум, внутри которого на градиенте не выделяется зон быстрого и медленного изменений видового состава сообществ. Этот тип континуума преобладает в тех случаях, когда изменение состава сообществ происходит без смены жизненной формы растений, т.е. меняется травяная или лесная растительность. Пример экоклина приведен на рис. 7, из него очевидно, что смена растительных сообществ на градиенте происходит постепенно и провести границы сообществ, соответствующих разным условиям засоления почвы, можно только условно.

Рис. 7. Экоклин травяной растительности поймы нижней Волги по градиенту общего засоления (список видов сокращен, по Golub, Mirkin, 1986). 1 – Phalaroides arundinacea, 2 – Bolboschoenus borodinii, 6 – Argusia sibirica, 7 – Atriplex litoralis, 8 – Tripolium vulgare.
Экотон – это тип континуума, при котором на градиенте формируются более или менее однородные сообщества, связанные зоной быстрого и видимого на глаз перехода. Типичный пример экотона – растительность опушки (рис. 8), т.е. зоны контакта леса и злаковника (луга или степи).
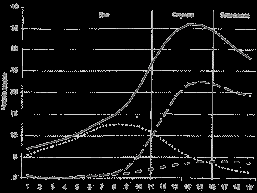
Рис. 8. Экотопный эффект в растительности лесной опушки (по Кучеровой, 2001). Число видов: 1 – общее, 2 – злаковников, 3 – лесных, 4 – опушечных.
Аналогичные континуумы типов экоклин и экотон проявляются и в характере изменения гетеротрофной биоты, в первую очередь насекомых и почвенных животных.
Контрольные вопросы
1. Объясните содержание принципа индивидуальности экологии вида.
2. Что такое экологические группы видов, с чем связана их условность?
3. Расскажите об истории развития и содержании концепции континуума.
4. Чем отличаются экоклин и экотон?
^
3.3. Принцип лимитирующих факторов
Суть этого принципа, связанного с именем Ю. Либиха, заключается в том, что наиболее важным для распределения вида является тот фактор, значения которого находятся в минимуме или в максимуме.
Как уже отмечалось, наличие в составе комплексного градиента лимитирующего фактора делает этот градиент ведущим, т.е. в большей степени, чем другие, определяющим состав и продуктивность экосистем и состояние популяций формирующих их видов. Так в зоне тундры основу ведущего комплексного градиента составляет количество тепла, так как влаги там достаточно, а обеспеченность элементами минерального питания также зависит от тепла: чем теплее субстраты, тем активнее идет в них процесс минерализации органического вещества и меньше накапливается неразложившихся остатков растений.
В зоне тайги лимитирующим фактором является обеспеченность почв питательными элементами. Почвы, формирующиеся на карбонатных породах, которые богаты кальцием и другими минеральными элементами, позволяют формироваться очень продуктивным сообществам. Однако в условиях тайги, особенно в южной ее части, возрастает роль второго ведущего комплексного градиента, связанного с увлажнением.
В лесостепной и степной зонах ведущий комплексный градиент формируют экологические факторы, связанные с лимитирующим фактором увлажнения, так как почвы в этих зонах (черноземы) богаты питательными элементами. Особенно остро влияние лимитирующего фактора проявляется в годы засух. На этот комплексный градиент может накладываться влияние выпаса (как отмечалось, выпас – это также комплексный градиент), а в южной части степной зоны – комплексного градиента засоления почвы.
В водных экосистемах для большинства входящих в их состав организмов лимитирующими факторами являются содержание кислорода и фосфора, а для растений, кроме того, – свет.
Для многих животных в условиях умеренного климата лимитирующим фактором является глубина снежного покрова. Свободное перемещение по глубокому снегу свойственно сравнительно небольшому числу видов, имеющих «лыжи» (заяц беляк, заяц русак, белая куропатка) или отличающихся «длинноногостью» (лось). Глубокий снег является препятствием для перемещения волка и кабана. Этот фактор определяет границы коневодства с круглогодичным содержанием животных на пастбищах. При глубоком снеге (в период зимнего выпаса – тебеневки) лошади не могут использовать корм.
Контрольные вопросы
1. Кто является автором принципа лимитирующих факторов, какие факторы называются лимитирующими?
2. Приведите примеры лимитирующих факторов в наземных экосистемах разных природных зон.
3. Какие факторы являются лимитирующими в водных экосистемах?
Темы докладов на семинарском занятии
1. Для чего и как выделяют экологические группы видов?
2. Концепция континуума: история и значение для экологии.
3. Роль лимитирующих факторов в определении состава биоты экосистем.