
Замалиева Фания Файзрахмановна
| Вид материала | Автореферат |
СодержаниеТеоретическое обоснование закономерностей в процессе нарастания |
- С. А. Замалиева Философские основания, 852.65kb.
- Республики Татарстан «Не стоит село без праведника…», 75.45kb.
- Хуснутдинова Фания Салимовна Село Старокутлумбетьево, 2011 год конспект, 79.64kb.
ОСВИТ - общая суммарная векторная интенсивность крылатых тлей за годы размножения семенного материала в открытом грунте, выраженная в кратных коэффициентах по отношению к критическому порогу; * - степень зараженности вирусами на стадии ССЭ; **- выделены положительные результаты, когда зараженность вирусами у СЭ составила не более 5%.
Мы проанализировали также коэффициенты роста вирусной зараженности у различных сортов в производственном размножении за период наблюдений с 2001 по 2006 годы (таблица 4). Нарастание вирусной зараженности от нуля до 1-2% проходит незаметно, так как мы пока неспособны обнаружить низкую степень зараженности, поэтому представленные в таблице коэффициенты роста определены после превышения указанного уровня степени зараженности. Из данных таблицы можно заметить, что в 2001 году коэффициенты роста повторной вирусной зараженности были высокими, что объясняется особыми условиями года, связанными с высокой векторной интенсивностью тлей и совпадением лета с активным ростом растений. В 2005 году высокий КРВЗ у сорта Леди Розетта связан с поздним сроком созревания сорта и растянутым летом многочисленных крылатых тлей. Можно отметить относительно более высокие КРВЗ у сортов Луговской и Юниор - 7,2 и 7,6, свидетельствующие о неустойчивости этих сортов к вирусному заражению. Остальные сорта имели следующие значения КРВЗ : Жуковский ранний – 1,7-3,8; Адретта – 1,2-2,5; Удача – 2,0; Розара – 2,0-6,0; Пранса – 3,0; Сантэ – 3,6; Невский – 1-3,7; Рождественский – 3,0-4,0; Лорх – 2,1-2,8; Чародей -1-6. Невысокие КРВЗ у большинства сортов после достижения 1-2% зараженности, свидетельствуют о том, что устойчивость этих сортов к вирусному заражению находится примерно на одном уровне с сортом Невский. Значит, высокую степень зараженности на конечном этапе получения суперэлиты (таблица 3) у сортов Жуковский ранний - 25%, Лорх - 16,6%, Розара, - 12,5%,Рождественский - 25% - можно объяснить только более быстрым ростом зараженности на предыдущих этапах процесса скрытого заражения (до достижения уровня 1-2%). Основным отличием этих сортов было то, что размножение большинства из них на стадии ППП, в отличие от Невского, проводилось на небольших площадях. То есть, здесь необходимо отметить, что площадь посадки на стадии ППП имеет определенное значение в процессе нарастания вирусной зараженности.
Таблица 4
Коэффициенты роста вирусной зараженности
у оздоровленных сортов картофеля за период с 2001 по 2006гг
| № | Годы | 2001 | 2002 | 2003 | 2004 | 2005 | 2006 |
| СВИТ | 2,56 КП | 0,9 КП | 4,3 КП | 0,9КП | 10,5 КП | 0,8 КП | |
| 1 | Жуковский ранний | - | 2,3 | 1,7 2,2 | - | - | 3,8 |
| 2 | Белоярский ранний | 14,5* | - | - | - | - | - |
| 3 | Адретта | - | 1,2 | 2,5 1,6 | - | - | - |
| 4 | Удача | - | - | - | - | - | 2,0 |
| 5 | Розара | - | - | - | - | 2,0 | 3,0 6,0 |
| 6 | Юниор | - | - | - | 7,6* | - | - |
| 7 | Пранса | - | - | - | 3,0 | - | - |
| 8 | Сантэ | - | 3,6 | - | - | - | - |
| 9 | Невский | - | 2,0 1,0 | 3,7 1,0 | - | - | 1,0 |
| 10 | Рождественский | - | - | 3,0 | - | 4,0 | - |
| 11 | Луговской | - | 7,2* | 2,8 | - | - | - |
| 12 | Лорх | - | 2,1 | 2,8 | - | - | - |
| 13 | Снегирь | 11,4* | - | - | - | - | - |
| 14 | Елизавета | 18,2* | - | - | - | - | - |
| 15 | Петербургский | 18,9* | - | - | - | - | - |
| 16 | Леди Розетта | - | - | - | - | 14,0* | - |
| 17 | Чародей | - | - | 1,0 | 6,0 | | |
СВИТ - суммарная векторная интенсивность крылатых тлей за вегетационный период, выраженная в кратных коэффициентах по отношению к критическому порогу;
*- повышенные значения коэффициентов роста вирусной зараженности.
Рассмотрим подробнее нарастание вирусной зараженности в репродукционном размножении от ППП до СЭ на примере сорта Невский, который занимал самые большие площади в производственном размножении на протяжении длительного времени. Из таблицы 5 видно, что циклы размножения оздоровленного сорта Невский, начатые в открытом грунте с 2001 и 2002 года заканчивались получением суперэлиты с высоким уровнем зараженности – в 2003г - 45,8%, в 2004 г – 20,8%, соответственно. Лишь циклы размножения, начатые в 2003 и 2004гг закончились получением суперэлиты с зараженностью на низком уровне (1,7-4,1%). На рисунке 1 представлены четыре трехлетних цикла размножения семенного картофеля сорта Невский – с 2001 по 2003гг, с 2002 по 2004гг, с 2003 по 2005гг, с 2004 по 2006гг. В каждом цикле можно увидеть динамику изменения вирусной зараженности оздоровленного семенного картофеля во время 3-хлетнего размножения от первого полевого поколения до суперсуперэлиты и далее до суперэлиты и сопуствующую ей векторную интенсивность тлей - по оси абсцисс показана численность переносчиков вирусов картофеля крылатых тлей в годы выращивания, выраженная в единицах, эквивалентных персиковой тле. Сопоставив все данные на рисунке, можно отметить три важных момента.
Во-первых, рост зараженности происходит у сорта Невский, чаще всего, в кратных коэффициентах, находящихся в интервале от 1 до 5. В годы с низкой численностью переносчиков (2002г, 2004г, 2006г) коэффициент роста вирусной зараженности небольшой - 1-2, в годы с высокой численностью переносчиков вирусов (2001г,2003г,2005г) коэффициент роста вирусной зараженности был выше - 3-5. Таким образом, рост вирусной зараженности сорта в течение вегетационного периода происходит с кратным коэффициентом, величина которого прямо пропорциональна векторной интенсивности крылатых тлей.
Таблица 5
Изменение вирусной инфицированности (ВИ) у сорта Невский
в производственном размножении по репродукциям с 2001 по 2006 годы
| Годы | Векторная интенсив-ность Экв.ед. | Репро-дукция | Площадь, га | Началь-ная заражен-ность, %±m | Конеч- ная заражен-ность, %±m | Коэффи-циент роста вирусной заражен-ности |
| 2001 | 130 | ППП | 0,8 | 5,2 ±2,2 | 20,8 ±4,1 | 4,0 |
| | | С-с-э | 2,6 | 18,0 ± 3,8 | 86,3 ±3,4 | 4,8 |
| | | С-э | 18,0 | 23,0 ± 4,2 | 88,6 ±3,2 | 3,8 |
| 2002 | 45 | ППП | 1,0 | 2,2 ± 1,5 | 4,1 ±2,0 | 2,0 |
| | | С-с-э | 7,7 | 12,5 ± 3,3 | 15,6 ±3,6 | 1,0 |
| | | С-э | 17,6 | 86,3 ± 3,4 | 100,0 | 1,2 |
| 2003 | 215 | ППП | 1,7 | 0,0 | 0,0 | неизвестен |
| | | С-с-э | 5,0 | 4,1 ± 2,0 | 16,6 ±3,7 | 4,0 |
| | | С-э | 41,0 | 12,5 ± 3,3 | 45,8 ±5,0 | 3,7 |
| 2004 | 45 | ППП | 2,4 | 0,0 | 0,0 | неизвестен |
| | | С-с-э | 16,6 | 0,0 | 0,0 | неизвестен |
| | | С-э | 36,4 | 16,6 ± 3,7 | 20,8 ±4,1 | 1,0 |
| 2005 | 525 | ППП | 1,0 | 0,0 | 0,0 | неизвестен |
| | | С-с-э | 20,0 | 0,0 | 1,7 ±1,3 | >1,7 |
| | | С-э | 43,0 | 0,0 | 4,1 ±2,0 | >4,1 |
| 2006 | 40 | ППП | 0,3 | 0,0 | 0,0 | неизвестен |
| | | С-с-э | 6,0 | 0,0 | 0,0 | неизвестен |
| | | С-э | 43,0 | 1,7 ± 1,3 | 1,7 ±1,3 | 1,0 |
Примечание: %±m – доля зараженности с соответствующей ошибкой.
Во-вторых, чем выше исходная зараженность первого полевого поколения, тем выше зараженность семенного картофеля в конце размножения. Так, в четырех циклах размножения исходная зараженность возрастает от 5,2 до 45,8%, от 2,2 до 16,6%, от 0 до 4,1% и от 0 до 1,7%. Здесь очевидна положительная зависимость между исходной и конечной степенью вирусной зараженности материала.
Третий момент, наиболее важный для семеноводства, заключается в том, что при низкой степени исходной зараженности в двух последних циклах размножения, степень зараженности возрастала незначительно даже при очень высокой численности крылатых тлей-переносчиков вирусов, которая наблюдалась в 2005 году. То есть, при низкой степени исходной зараженности материала положительная зависимость между нарастанием степени зараженности вирусами и численностью крылатых тлей исчезала.
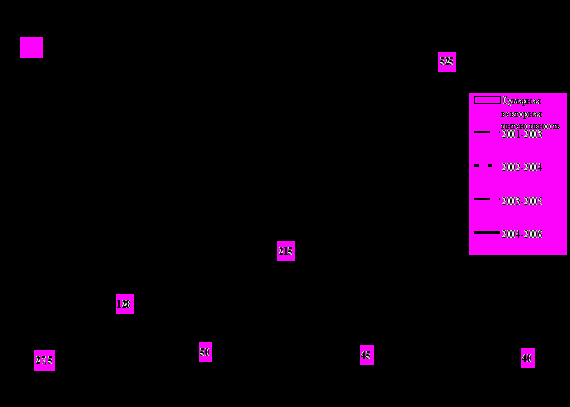
Рис. 1. Зависимость скорости реинфекции оздоровленного семенного картофеля от исходной зараженности и суммарной векторной интенсивности крылатых тлей.
Примечание: в 2000-2001гг указана средневзвешенная зараженность миниклубней на поле, где произрастал Невский – 5,2%, в 2001-2002гг указана зараженность после проведения тщательной браковки и снижения зараженности сорта до 12,5%.
Таким образом, на основании проведенных наблюдений в производственном размножении оздоровленного картофеля, мы выделили основные факторы и закономерности, от которых зависит интенсивность повторного заражения вирусами сортовой репродукции:
- Устойчивость сорта к заражению вирусами;
- Площадь посадки оздоровленного картофеля - особенно в первый год размножения в открытом грунте;
- Степень исходной вирусной зараженности оздоровленного картофеля;
- Суммарная векторная интенсивность крылатых тлей, динамика их лета, сроки наступления критического порога численности в текущем вегетационном периоде;
- Совпадение сроков массового лета тлей с фазами активного роста сортов картофеля;
- Кратный рост степени повторной вирусной зараженности в течение вегетационного периода, величина которого зависит, в основном, от устойчивости сорта картофеля и векторной интенсивности тлей;
7. Снижение зависимости нарастания повторной вирусной зараженности оздоровленного картофеля от численности тлей, происходящее при низкой степени исходной зараженности оздоровленного картофеля.
Теоретическое обоснование закономерностей в процессе нарастания
степени повторной зараженности вирусами оздоровленного семенного картофеля
Для объяснения всех полученных закономерностей необходимо теоретическое обоснование, объясняющее процессы распространения вирусов крылатыми тлями и роста степени повторной зараженности вирусами оздоровленного картофеля, происходящие в репродукционном размножении семенного материала. Рассмотрим отдельные этапы этого процесса с применением методов моделирования биологических процессов и математических расчетов.
Динамика нарастания степени повторной зараженности вирусами оздоровленного картофеля при его репродуцировании
Представим модель роста степени повторной зараженности вирусами сорта Невский с ежегодным коэффициентом, равным 4, на протяжении 12 лет (таблица 6, рисунок 2).
Таблица 6
Модель ежегодного 4-кратного роста повторной
зараженности вирусами оздоровленного картофеля в течение 12-летнего периода
-
Год
размножения
Степень зараженности посадок в период вегетации, %
Периоды
накопления вирусов
1
0,000006
Длительный
скрытый
2
0,000025
3
0,0001
4
0,0004
5
0,0015
6
0,006
7
0,025
8
0,1
9
0,4
10
1,6
Короткий обнаруживаемый
11
6,3
12
25