
Азотофіксація І продуктивність сої
| Вид материала | Документы |
- Реферат проект висвітлює питання теоретичного І агроекологічного обґрунтування розробки, 83.34kb.
- Використання різних доз азотних добрив І бактеріальних препаратів при вирощуванні сої, 527.45kb.
- Державний реєстр пестицидів І агрохімікатів, дозволених до використання в Україні, 2594.96kb.
- "Органогенні елементи", 93.03kb.
- Праця, кадри та соціальний розвиток колективу, 134.73kb.
- В. С. Бончик, кандидат технічних наук, доцент кафедри сільськогосподарських машин пдата,, 15.83kb.
- Реферат на тему, 165.84kb.
- Реферат на тему, 301.53kb.
- Центральна та східна Європа, 1276.46kb.
- Реферат на тему: Обє’кти аудиту Об'єкти аудиту, 56.55kb.
Азотофіксація і продуктивність сої.
. Роль мінерального і симбіотичного азоту в живленні сої.
До найбільш важливих біологічних процесів, які мають глобальну післядію для біосфери відносять фотосинтез і азотофіксацію. Від фіксації молекулярного азоту, яку здійснюють обмежене число мікорорганізмів, залежить існування життя на Землі в тій же пропорції, в якій воно залежить від фотосинтезу як джерела енергії. Якби азот, який виноситься з грунту, постійно не повертався знову в грунт, життя на планеті повільно припинилось (С.Ф. Измайлов, 1996).
Важливою особливістю сої являється здатність то ендосимбіозу з азотофіксуючими суббактеріями – ризобіями. Завдяки азотофіксації, яка проходить в сформованих в симбіозі з ризобіями бульбочками, соя може, в значній мірі, або навіть повністю задовольняти свою потребу в азоті (симбіотрофне живлення азотом). Це знижує залежність рослини від наявності азотних сполук в грунті і дозволяє вирощувати її при відсутності або при мінімальному використанні дорогих і екологічно небезпечних азотних добрив. В той же час бобові культури мають звичайну для інших рослин властивість до поглинання з грунту і асиміляції мінеральних і органічних сполук азоту (Н.А. Проворов, 1996).
Не дивлячись на значну кількість робіт, присвячених аналізу фізіологічно-біологічних механізмів фіксації молекулярного і асиміляції азоту бобовими, співвідношення симбіотичного і автотрофного азотного живлення не можу вважатись достатнього вивченим (В.Л. Кретович, 1994; Н.А. Проворов, 1996). У сої відносна ефективність використання фіксованого азоту і азоту з мінеральних добрив суттєво залежить від сорту і умов вирощування рослин і прибавки від інокуляції в багатьох випадках можу бути вище, ніж від внесення азоту (В.Ф. Патика, Н.З. Толкачев, В.И. Завирюхин, Н.А. Саенко, 1987; F.F. Dugue, М.С. Neves, A.A. Franko, 1985; R.J. Rennie, S. Dubetz, J.B. Dole, H.H. Muendel, 1982).
Таблиця №1.
Порівняльна характеристика сортів сої Х005 і Maple Presto за властивістю використовувати різні джерела азоту (R. Rennie, 1982)
| Параметри азотонакопичення | Х005 | Maple Presto |
| 1. Загальна кількість фіксованого азоту, кг/га | 115 | 82 |
| 2. Загальне накопичення азоту в надземній частині рослин, мг на 1 рослину.
| 1039 605 | 882 842 |
| 3. Доля інокульованого азоту в загальному азотному балансі рослин: а) засвоєного з атмосфери, %
б) засвоєного з добрив при внесенні 160 кг/га азоту, % | 60 53 19 | 53 38 31 |
Таким чином, одним з важливих зовнішніх факторів, які здійснюють вплив на утворення і розвиток кореневих бульбочок сої і їх азотофіксуючу активність, являється мінеральний азот. Його високий вміст в грунті затримує появу бульбочок і знижує інтенсивність азотофіксації. Невеликі дози азоту можуть здійснювати стимулюючу дію (Л.Ю. Верниченко, Ю.М. Миллер, 1983; А. Eagleshat, S. Hassonta, R. Seegers, 1983).
Середні і високі дози зв'язаного азоту знижують ефективність функціонування симбіотичної системи і не завжди сприяють росту врожаю, а в деяких випадках ведуть до його зниження (Б.А. Ягодин, М.В. Вильям, Ю.П. Сазонов, 1984; D. McNeil, T. Zarue, 1984). Дія невисоких стартових доз азотнних добрив на урожай сої також залежить від її сорту, штамів в Rhizobium, умов вирощування (А. Russhel, P, Vose, E. Matsui, 1982; Л.М. Афанасьев, Л.М. Доросинский, А.П. Кожемяков, 1983; В.Ф. Патика, И.З. Толкачев, В.И. Заверюхин, И.П. Саенко, 1987).
Причини таких різних думок залишаються незрозумілими, а погляди про доцільність використання стартових доз азоту в практиці рослинництва мають протиріччя (I. Freire, 1984; F. Warembourg, M. Feerhandex, 1985).
Одним з підходів до вирішення питання оптимізації азотного живлення бобових являються дослідження про взаємозв'язки процесів симбіотичної азотофіксації і асиміляції мінерального азоту в забезпеченні сої азотом і кількісна оцінка цих процесів по їх внеску в загальний азотний фонд. В цьому напрямку досліджень оцінюється внесок різних джерел азоту в кінцевий врожай, але не приділяється достатньої уваги їх ролі в формуванні азотного статусу рослин в процесі росту і розвитку, особливо на ранніх стадіях (Л.Ю. Верничення, Ю.М. Миллер, 1983; А. Eaglesham, S. Suegers, 1983, J. Silbury, D. Catchpoole, W.Wallace, 1986).
Соя характеризується відносно повільними темпами накопичення сухої речовини і азоту на ранній стадіях онтогенезу і високою інтенсивністю цих процесів в період плодоутворення (Т.Я. Жизневская, В.Б. Ильясова, 1979; А.Т. Грицун, 1981). Мінеральний азот для сої має суттєву роль в період вегетативного росту. Починаючи з цвітіння джерелом азотного живлення стає азотофіксація. Високі темпи азотофіксації на початку репродуктивної фази підтримуються за рахунок посилення активності одиниці маси бульбочок, а пізніше – за рахунок збільшення маси. В період від початку плодоутворення до наливання зерна в рослини сої поступило 55-60% від загальної кількості азоту, фіксованого за вегетацію. Тому ріст бобів і наливання зерна здійснювались, головним чином, шляхом прямого, шляхом прямого використання фіксованого азоту, а не за рахунок реутилізації раніше накопиченого азоту і зниження його вмісту у вегетативних органах (Г.Н. Троицкая, А.Г. Гидимов, С.Ф. Измайлов, 1993).
Враховуючи ці дослідження можна зробити висновок, що дивлячись на високу енергоємність процесу симбіотичної азотофіксації, величезну потребу генеративних органів в елементах живлення і конкуренцію їх з бульбочками за фотосинтезами, соя має властивість підтримувати активне функціонування фіксуючої азот симбіотичної системи навіть в період активного плодоутворення. Підвищена потреба бобів в азоті являється важливим фактором, який визначає на рівні цілої рослини високі темпи азотофіксації в репродуктивний період (J.Jimsande, 1998), на основі показників розвитку азотофіксуючої активності в бульбочках сої проходить до висновку, що потреба в азоті і швидкість фотосинтезу визначають в кінці-кінців загальну активність азотофіксації.
На основі аналізу мінеральних даних можна зробити висновок, що при ранньому утворенні бульбочок і високоефективному симбіозі соя формує підвищений врожай в основному за рахунок симбіотичного азоту. Кількість азоту, яка необхідна для підтримання росту рослин до включення в процес азотофіксації невелика і може бути забезпеченою грунтовими запасами. Не виключена роль стартових доз азотних добрив, особливо на бідних грунтах, для страхування рослин від можливої нестачі азоту на випадок затримки появи бульбочкових і повільного їх розвитку при несприятливих умовах.
В останні роки в селекції бульбочкових бактерій, які використовуються для інокуляції сої, ведеться пошук не тільки високоактивних і конкурентоспроможних штамів, але і штамів з високою відносно ефективністю азотофіксації, які мають властивість до рециклювання молекулярного водню в бульбочках. Це забезпечує рослинам мінімалізацією енергетичних витрат на одиницю фіксованого азоту (Бонарцева Г.А., Мышкина В.А., Мишустин Е.Н, 1983; Hanus F.J, Albrecht S.H, Zablotowicz R.M, 1981; Miller R.W, Sirois J.C., 1982).
Для оцінки відносної ефективності азотофіксації (ВЕА) симбіозу Rhizobium – проводили не тільки визначення нітрогенезної активності ацетиленовим методом, але паралельно визначали кількість екзогенного водню за методикою Sehnbert K.R., Evans H,J, (1978): ВЕА=1 - Нг/ЕгНг. З формули видно, що відносна ефективність азотофіксації вище при меншому виділенні екзогенного водню на одиницю відновленого ацетилену.
Садиков Б.Ф., Зуева Л.Д., Воронялова Н.А. (1985) визначили можливість використання показників нітрогенезної активності і відносної ефективності азотофіксації у сої в симбіозі з бульбочковими бактеріями для відбору перспективних штамів і оцінки впливу невисоких доз азотних добрив на симбіотичну азотофіксацію. Виявилось, що споживання азоту врожаєм зерна корелювало з нітрогенезною активністю або відносною активністю азотофіксації. Найбільший коефіцієнт кореляції був між показниками, що характеризують винос азоту і відношення нітрогенезної активності до відносної ефективність азотофіксації (табл.1)
Таблиця №2.
Кореляція між виносом азоту врожаєм сої, інокульованої різними штамами бульбочкових бактерій з азотофіксацією і відносної ефективністю азотофіксації в симбіозі (Б.Ф. Садиков, Л.Д. Зуева, А.У. Каппушев, И.А. Воронкова, 1985)
| Штам | Нітрогенезна активність (НА), мкл С2Н2/росли./г. | Відносна ефективність азотофіксації (ВЕА) 1-Н2г:С2Н4 | НА : ВЕА, мкл/росл./г | N, кг/га |
| Контроль | - | - | - | 91,6 |
| N70 | - | - | - | 97,5 |
| 6345 | 8,2 | 0,92 | 7,5 | 107,9 |
| 629а | 6,0 | 0,6 | 3,6 | 100,8 |
| 647а | 8,7 | 0,69 | 5,9 | 101,6 |
| 645аа | 13,4 | 0,63 | 8,4 | 103,0 |
| 639б | 14,6 | 0,92 | 13,4 | 106,7 |
| 648а | 13,1 | 0,69 | 9,0 | 103,7 |
| 645б | 12,6 | 0,44 | 5,6 | 107,4 |
| 644б | 13,1 | 0,46 | 6,0 | 107,8 |
| 381 | 13,3 | 0,90 | 12,0 | 117,1 |
| 87 | 12,4 | 0,53 | 6,7 | 97,7 |
| Середнє | 11,5 | 0,68 | 7,8 | 105,4 |
| Коефіцієнт кореляції | 0,40 | 0,57 | 0,63 | - |
Результатами іонометричного визначення вмісту мінеральних форм азоту N-NH4 i N-NO3 в коренях і листках сої при внесення в грунт 18 і 35 кг\га N у вигляді аміачної селітри подані в табл.№2. Враховуючи те, що наявність азоту аміачної форми в рослинах пов’язане з азотофіксацією і споживанням азоту з грунту, а вміст N-NO3 виключно з мінерального живлення, то зниження відношення N-NH4 : N-NO3 вказує на зменшення долі симбіотичного азоту в живленні сої при внесенні азотних добрив. Ще більші відмінності у співвідношенні форм азоту були характерними для листків сої в порівнянні з коренями (табл.№2)
Таблиця №3
Співвідношення мінеральних форм азоту в коренях і листках сої при зміні долі симбіотичного і мінерального живлення (сорт Чиппева, початок цвітіння)
| Доза мінерального азоту, кг/га | Вміст азоту, мг/кг сухої речовини | |||||||
| Грунт | Листки | Корені | ||||||
| NH4 | NO3 | NH4 | NO3 | NH4 ––––– NO3 | NH4 | NO3 | NH4 ––––– NO3 | |
Котроль | 5,9 | 5,3 | 964 | 309 | 3,12 | 570 | 652 | 1,01 |
| N18 | 6,5 | 5,3 | 1068 | 355 | 3,0 | 676 | 661 | 1,02 |
| N35 | 6,4 | 5,2 | 903 | 822 | 1,10 | 576 | 669 | 0,86 |
Нашими дослідженнями симбіотичної азотофіксації сої в залежності від внесення азотних добрив встановлено, що кількість бульбочок на одну рослину в значній мірі залежала від вмісту азоту в грунті (табл.№3). Спостерігалась тенденція зменшення кількості бульбочок і їх маси з однієї рослини при збільшенні дози азотних добрив. Вміст нітратного азоту в грунті без додаткового внесення добрив був високим, тому навіть невисока доза азотних добрив приводила до зниження азотофіксації.
Таблиця №4
Вплив азотних добрив на кількість і масу бульбочок у сої та вміст мінеральних форм азоту в грунті (1997р.)
| Показники | Фази розвитку сої | ||||||||
| Галузіння | Цвітіння | Зелена стиглість бобів | |||||||
| Без добрив | N30 | N60 | Без добрив | N30 | N60 | Без добрив | N30 | N60 | |
| 1. Число бульбочок на 1 рослину, шт | 3,0 | 3,5 | 4,0 | 7,2 | 6,3 | 6,1 | 12,8 | 10,3 | 8,1 |
| 2. Маса бульбочок на 1 рослину, мг | 23 | 28 | 34 | 91 | 63 | 50 | 287 | 201 | 153 |
| 3. Вміст в грунті мінерального азоту (мг/кг грунту) N – NO3 | 10,8 | 17,5 | 20,2 | 9,4 | 19,7 | 21,1 | 4,1 | 6,7 | 7,2 |
| N – NH4 | 6,8 | 8,6 | 10,3 | 14,8 | 15,7 | 15,9 | 8,0 | 9,2 | 9,7 |
3.2 Інтенсивність освітлення посівів: азотофіксація.
Фотосинтез – процес засвоєння зеленими рослинами не тільки енергії і вуглекислого газу, але й в значній мірі такого важливого компоненту, як азот. У більшості випадків субстрактами живлення рослин азотом являються іони мінеральних солей (NO3, NH4). Соя, як бобова рослина, здатна засвоювати молекулярний азот (N2) з повітря за допомогою азотофіксуючих бактерій. Питання взаємозв'язку процесу фотосинтезу бобових рослин і фіксації атмосферного азоту бактріями в кореневих бульбочках давно являються предметом спеціальних досліджень. Засвоєння вуглецю і азоту вимагає участі енергії фотосинтетичного походження. Серед прямих продуктів фотосинтезу знаходяться як азотисті так і безазотисті сполуки, тому засвоєння цих елементів являється важливою основою живлення рослин, і звичайно, їх продуктивності (А.А. Ничипорович, 1971, Т.Ф. Андреева, 1982).
Тільки при нормальному співвідношенні цих процесів проходить оптимальний ріст і розвиток рослин, забезпечується їх висока продуктивність.
Аналіз мінеральних джерел показує, що не у всіх видів бобових рослин процес азотофіксації забезпечує достатньою кількістю азоту нормальний ріст і розвиток. Як відомо з попереднього розділу, багато дослідників рекомендують в посівах сої внесення "стартових" доз мінерального азоту. Нестача азоту особливо проявляється у сої на початку вегетації, так як бульбочки формуються дуже повільно і достигають лише після цвітіння рослин (D. Phillips, 1980; L.Williams, T. De Jong, D. Phillips, 1981).
Протягом вегетативного росту соя в польових умовах фіксує лише 10% азоту (від сумарної азотофіксації), 90% засвоюється в репродуктивний період (R.Hatdy, U.Havelka, 1975). В той же час потреба в азоті у рослин сої велика як у вегетативну фазу, так і в період цвітіння – достигання бобів, які містять багато білків.
В дослідженнях Нгуен Тхи Чи, Т.Ф. Андреевой, Л.Е. Строгонова (1983) сою вирощували при інтенсивності 60; 120 вт/м2. Ефект дії світла на накопичення загальної біомаси, маси бульбочок і фіксації азоту знаходяться в більшій залежності від віку рослин (табл.№1).
Таблиця №5.
Вплив інтенсивності освітлення сої на фіксацію атмосферного азоту.
| Фаза розвитку рослини | Інтенсивність світла, вт/м2 | Суха маса, г | Потенціальний фотосинтез, мг СО2/дм2 за 1 год | Активність азотофіксації 1 рослини, Нмоль С2Н4/хв | |
| рослини | бульбочок | ||||
| Бутонізація | 60 | 1,25 | 0,02 | 34,9 | 5,5 |
| 120 | 1.84 | 0,04 | 34,0 | 12,0 | |
| Цвітіння | 60 | 2,87 | 0,04 | 50,0 | 47,9 |
| 120 | 3,43 | 0,13 | 48,0 | 114,9 | |
| Утворення бобів | 60 | 6,18 | 0,13 | 58,0 | 47,5 |
| 120 | 7,62 | 0,27 | 57,2 | 44,9 | |
Приведені дані показують, що при підвищенній інтенсивності освітлення значно посилюється ріст бульбочок при відсутній зміні питомої активності азотофіксації. Але загальна фіксація азоту на рослину збільшується.
Позитивну дію освітлення високої інтенсивності на ріст бульбочок пов'язують не тільки з підвищенням інтенсивності фотосинтезу, а також з посиленням відтоку органічних речовин в кореневу систему і бульбочки (S. Sheikholelam, K.Fishbeck, D.Phillips, 1980).
За даними F.Minchin, J.Pate (1973), тільки 26% вуглецю, засвоєного в процесі фотосинтезу, залишається в надземній частині, 42% - трансформується до коренів і 32% надходить безпосередньо в бульбочки.
Для вияснення залежності між фотосинтезом і азотним обміном у сої цікаві результати одержані Т.Ч. Нгуеном, Т.Ф. Андреевой, Л.Е. Строгановой (1983). Рослини, заражені Phizobint, вирощували при 0,5 і 1 мМ азоту (табл. №2). Протягом 20 днів рослини росли при інтенсивності освітлення 80 Вт/м2, потім їх перемістили на 15 днів на світло високої інтенсивності (200 Вт/м2)
Таблиця №6.
Вплив азоту та інтенсивності освітлення на фотосинтез та азотофіксацію сої
| Фаза розвитку рослини | Інтенсив-ність світла, вт/м2 | Норма азоту, мМ | Суха маса, г | Площа листків, дм2/рослин | Потен-ціальний фотосин-тез, мг СО2/дм2 за 1 год | Активність азотофіксації 1 рослини, Нмоль С2Н4/хв | |
| росли-ни | бульбо-чок | ||||||
| Бутонізація | 80 | 0,5 | 1,04 | 0,04 | 1,75 | 9,8 | 49,5 |
| 1,0 | 1,38 | 0,02 | 2,30 | 12,0 | 23,9 | ||
| Утворення бобів | 80 | 0,5 | 3,21 | 0,09 | 3,32 | 12,5 | 29,2 |
| 1,0 | 5,05 | 0,06 | 5.89 | 15,0 | 33,8 | ||
| 200 | 0,5 | 3,84 | 0,16 | 3,77 | 14,8 | 29,8 | |
| 1,0 | 5,92 | 0,10 | 6,16 | 16,0 | 61,5 | ||
В даних умовах освітлення кількість фіксованого рослиною атмосферного азоту було недостатнім при такій забезпеченості мінеральним азотом, рослини відчували нестачу в азотному живленні. Це підтверджується незначним збільшенням сухої біомаси рослин, зниженням азоту в листках при вирощуванні при інтенсивному освітленні. Нестача мінерального азоту в грунті обмежує фотосинтетичну активність рослин. Фактори, які підвищують фотосинтез (світло, концентрація СО2, температура) в умовах азотного "голоду" не здійснюють позитивного впливу на інтенсивність фотосинтезу (L.Williams, D.Phillips, 1980). Разом з тим було доведено на рослинах сої, що фотосинтез в умовах нестачі азоту в грунті можна підвищувати шляхом інокуляції активним штамом Rhizobium (R. Maiet, W.Brill, 1978). В результаті підвищення фотосинтетичної діяльності рослин збільшується приріст їх біомаси, ріст бульбочок і фіксація азоту в розрахунку на одну рослину.
Таким чином, інтенсивність освітлення здійснює суттєвий вплив на формування бульбочок на рослинах сої. Різні умови освітлення не приводять до зміни питомої активності азотофіксації. Посилення росту бульбочок являється основною причиною збільшення азотофіксації рослин. У вегетативний період росту і розвитку сої, а також при утворенні бобів у умовах дефіциту азоту рослини формують неповноцінний фотосинтетичний апарат з низькою інтенсивністю фотосинтезу, яку не вдається підвищити за допомогою кращої освітленості. Стає очевидним, що поліпшення азотного живлення у вказані періоди поліпшує активність фотосинтетичного апарату і забезпечує більш високу азотофіксацію рослин. Основним завданням підвищення продуктивності рослин сої являється поліпшення азотного живлення шляхом активізації процесу фіксації молекулярного азоту за рахунок використання активних штамів Rhizobium, або внесення невеликих доз азотних добрив.
В практиці рослинництва часто застосовують загущені посіви, а також змішані посіви сої із злаковими культурами, які нерідко відрізняються інтенсивним ростом і займають верхній ярус травостою. В цих випадках часто погіршуються умови освітлення, що приводить до значних змін ростових процесів, накопичення біомаси і звичайно на продуктивність посівів.
Аналіз літературних джерел показує, що затінення рослин часто приводить до утворення в кореневих бульбочках бобових метаболічних змін, які схожі із старінням (I. Soronen, 1970, T.Ching, S. Headtke, S.Russel, 1975).. При нестачі світла знижується вміст. Спостерігається зниження загального вмісту аскорбінової кислоти в кореневих бульбочках, що спостерігається при нормальному вмісту гемоглобіну в бульбочках сої, що свідчить про значні зміни в функціонуванні клітин рослини-господаря (К. Swaraj, O. Gard, 1976).
Вивченню дії затемнення різної тривалості на відносну ефективність азотофіксації, виділення вуглекислоти і води кореневими бульбочками присв'яені дослідження К. Сквардж, П.Н. Дуброво, С.В. Шищенко (1985). При тривалому затемненні знижується нітрогенезна активність на 60-80% (табл.№1). Паралельно спостерігається зниження дихання бульбочок, що пов'язано з зменшенням кількості синтезованих продуктів, які б могли бути у вигляді субстратів дихання. Однією з причин різкого зниження нітрогенезної активності при затіненні може бути зниження доступності АТФ, так як процес азотофіксації проходить при значних витратах енергії.
Соя, на відміну від інших зернобобових рослин має високу ступінь зворотних змін в кореневих бульбочках при поліпшенні світлового режиму. Експериментальні дані свідчать про відновлення азотофіксації на високому рівні, на відміну від порушення умов водного дефіциту, коли після проведення поливу азотофіксуюча активність бульбочок не відновлюється до зворотнього рівня (S.Edie, D. Phillips, 1983).
Таблиця №7.
Вплив тривалості теплового періоду на ацетилен відновлюючи активність, виділення СО2 і Н2 бульбочками.
| В 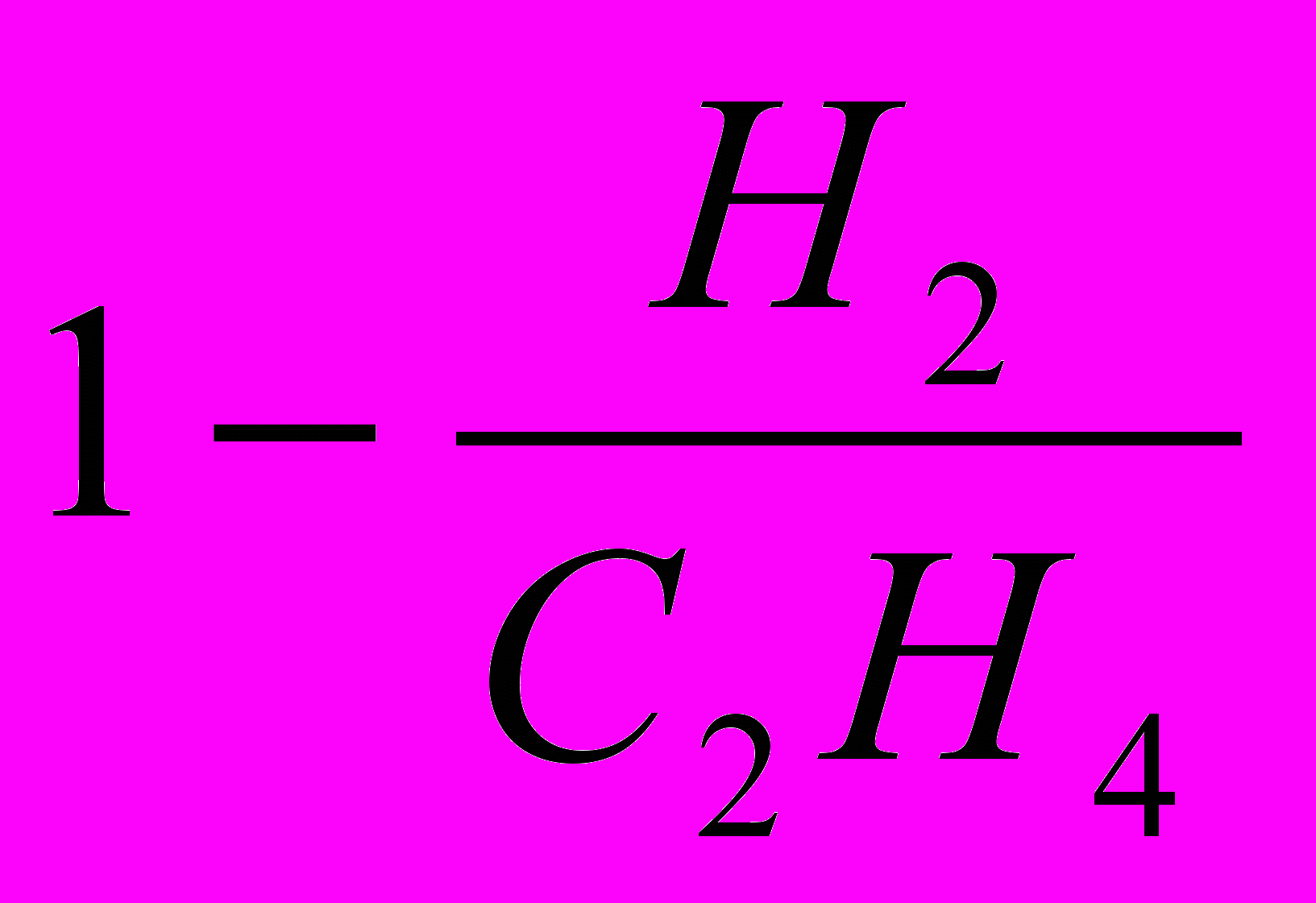 аріант аріант | Ацетилен- відновлюча активність (виділення С2Н2) | Дихання (виділення СО2) | Виділення Н2 | Енергетична ємкість азотофіксації, СО2/С2Н4 | Відносна ефективність азотофіксації,  |
| | |||||
| Світловий період 16 год на добу | 8,06 | 101,5 | 4,94 | 12,6 | 0,39 |
| Темнота: 18 год. | 3,94 | 53,5 | 1,80 | 13,6 | 0,54 |
| 42 год. | 2,76 | 32,5 | 1,19 | 11,8 | 0,57 |
| 66 год. | 1,63 | 23,4 | 1,60 | 14,3 | 0,02 |
| 90 год. | 0,78 | 22,7 | 0,78 | 29,2 | 0,00 |
Причиною падіння азотофіксуючої активності в бульбочках сої при затіненні являється погіршенні постачання бульбочок фотоасимілятами.
5.3.3. Водний режим грунту і азотофіксація.
Водний дефіцит. Бобові рослини дуже негативно сприймають як дефіцит вологи, так і надмірне зволоження в прикореневій зоні. В літературі зустрічається багато даних про чутливість кореневих бульбочок, а разом з тим і процесу симбіотичної азотофіксації до водного дефіциту (S. Edie, D. Phillips, 1983; K.Cvaradg, C.B. Шищенко, Т.И. Козлова, 1984). Нестача вологи в грунті може визивати як незворотні, так і зворотні пошкодження симбіотичної азотофіксуючої системи, які спричиняють пошкодження структури бульбочок в результаті механічного ефекту десикації (J. Sprent, 1972). В результаті посухи може припинитись надходження продуктів фотосинтезу в бульбочки при підсиханні рослин.
Прогресуюча витрата азотофіксуючої активності в бульбочках сої при підсиханні їх поверхні пов'язана із зниження швидкості дифузії кисню в середину бульбочок (C. Pankhuts, J.Spient, 1975).
При нестачі води, в першу чергу, гинуть рослини, як ростуть на симбіотично-зв’язаному азоті, у порівнянні з бобовими, які одержують мінеральний азот у вигляді добрив (H.Zahtan, J.Sprent, 1983).
Скварадж К, Шищенко С.В., Козлова Г.И (1984) для вияснення причин зниження азотофіксуючої активності бульбочок сої при різних умовах водного дефіциту провели серію лабораторних експериментів в піщаній культурі. Встановили, що протягом перших двох днів після припинення поливу насиченість піщаного субстрату знизилась на 38%, але вміст води в бульбочках знизився лише на 1% (табл. №8.). Наступне зниження вмісту води (3 дні після припинення поливу) привело до різкого зменшення вмісту води в бульбочках. Через день після відновлення поливу (після 2 днів без поливу) дефіцит насичення бульбочок водою відновився до рівня контролю складаючи лише 6,6%
Таблиця№8.
Характеристика водного режиму грунту сої в умовах водного дефіциту і наступного поливу
| Варіант | Вологість піску, % від ПВ | Вміст води в бульбоч-ках, % | Дефіцит насичення бульбочок водою, % | Суха речовина, % | |
| в бульбочках | в листках | ||||
Контроль | 51,8 | 72,6 | 3,5 | 27,4 | 25,3 |
| 2 дні без поливу | 13,1 | 71,5 | 16,0 | 28,5 | 25,6 |
| 3 дні без поливу | 2,0 | 59,9 | 44,4 | 40,1 | 41,0 |
| 2 дні без поливу + 1 день відновлення поливу | 32,9 | 72,3 | 6,6 | 27,6 | 27,3 |
Дія посухи в першу чергу відобразилась на бульбочках, ніж на листках. Вміст води першого повністю розгорнутого листка через 2 дні після припинення поливу був практично таким як і на контролі. Протягом перших двох днів дефіциту вологи не було відмічено ніяких ознак засихання листків сої. На 3-й день після припинення поливу вміст води в листках суттєво змінився, а фотосинтез повністю був подавлений. Зниження симбіотичної азотофіксації при несприятливих умовах сої може бути реакцією рослини для пристосування до даних кліматичних умов, що підвищує виживаність рослин і їх симбіотичної системи. Однією з причин зниження нітрогенезної активності і дихання бульбочок при водному дефіциті є не пряма дія дефіциту вологи на фотосинтез. Зниження інтенсивності фотосинтезу в цих умовах спостерігається, головним чином, за рахунок падіння тургору листків і закривання чечевичок.
Порівнюючи вплив водного дефіциту на листковий апарат і на бульбочки, можна зробити висновок, що азотофіксуюча система бульбочок сої сильніше реагувала на нестачу вологи, ніж надземна маса сої.
Перезволоження грунту. Азотофіксуючі бульбочки потребують в достатній кількості кисень. Але в польових умовах вміст кисню в ризосфері може бути знижений завдяки фотопоглинанню коренями, грунтовими аеробними мікроорганізмами і обмеженому газовому обміні. Особливо значне зниження вмісту кисню в грунті спостерігається при перезволоженні грунту і затопленні її водою. Це призводить до зниження азотофіксуючої активності бульбочок, яка залежить від багатьох причин. Це – родові, видові і соротві відмінності рослин, вид штаму Rhizobium, фази розвитку рослин, які знаходять на перезволожених грунтах, тривалість дії затоплення, температура зовнішнього середовища (A.Gallacher, J. Sprent, 1978).
Соя і кормові боби значно стійкіші до затоплення кореневої системи сої затримують фізіологічні процеси в бульбочках, активність фотосинтезу в листках і переміщення асимілянтів з листків в корені і бульбочки (В.Е. Косьмакова, Е.Г. Зверева, 1973). Анаеробні умови впливають безпосередньо на дихальну систему клітин бульбочок, знижають утворення АТФ, необхідної для процесу азотофіксації. При значній нестачі кисню в оточуючому середовищі клітини бульбочок переходять на анаеробний метаболізм, в результаті чого збільшується утворення етанолу і вуглекислого газу (J. I. Sptent, A. Gallacher, 1976).
Тривале перезволоження викликає в бульбочках морфологічні і анатомічні зміни, які носять часто адаптивний характер. Рослини в таких умовах пристосовуються до даних умов шляхом утворення поверхневих коренів. Ці корені розміщуються вище поверхні перезволоженого грунту. На таких коренях утворюються нові бульбочки, маса яких перевищує масу бульбочок сої, які виросли при нормальному зволоженні (М. Hunter, P.Jabtun, L. Byth, 1980; R. Trocason, R. Zawn, D. Byth, 1981). На перезволожених грунтах в бульбочках сої спостерігається розвиток поверхнево-розміщених, нещільно зложених видовжених клітин, які не затримують вільне переміщення газів в середину бульбочки і в зворотньому напрямі (C. Pakhurst, J.Sprent, 1975).
В дослідженнях Андрєєвої І.Н., Скварадж К.Г., Козлової Г.І.(1987) виявлений взаємозв’язок структурних і функціональних змін в бульбочках сої в умовах перезволоження грунту. Перебування кореневої системи сої в умовах затінення знижувало нітрогенезну активність бульбочок (табл.9).
Таблиця №9.
Вплив перозволоження кореневої системи на нітрогінезну активність і виділення СО 2 бульбочками сої(мкмоль/г сирої маси бульбочок за 1год).
| Варіант | Нітрогінезна активність (виділення С2Н4) | Виділення СО2 | ||
| 30 хв. | 60 хв. | 30 хв | 60 хв | |
Контроль (нормальне зволоження) | - | 8,0 | - | 34,1 |
| Затемнення : 1 доба | 5,1 | 7,7 | 24,0 | 23,4 |
| 2 доби | 3,6 | 4,1 | 31,6 | 35,0 |
| 4 доби | 2,7 | 3,5 | 28,4 | 29,0 |
Відновлення нітрогенезної активності після 2 добового затоплення було можливим, так як бульбочки верхнього ярусу не були пошкодженні і продовжували нормально функціонувати після відновлення аеробних умов. Після 4-добового затоплення зберігається незначна можливість до відновлення нітрогенезної активності. Наявність незначного рівня азотофіксуючої активності після 4-добового затоплення пояснюється збереженням симбіотичної системи частин бульбочок верхнього ярусу. В бульбочках нижнього і середнього ярусів проходить розпад бактероїдів і деградація органел.
5.3.4. Вплив гербіцидів на азотофіксацію сої.
При інтенсивній технології вирощування сої обов'язковим заходом являється грунтовим заходом являється використання грунтових і страхових гербіцидів, які являються високоактивними сполуками і здійснюють рівносторонню фізіологічну дію на процеси метаболізму рослин і, звичайно, на мікросимбіонта. Тому гербіциди прямо пов'язані з азотофіксацією. Круглов Ю.В, Пороменская Л.Н. (1986) відмічають негативний ефект гербіцидів на симбіотичні взаємовідносини в бобово-ризобіальній системі.
На посівах сої найбільш широко використоують тефлан, який має широкий спектр дії на бур'яни (Майер-Баде Г, 1972). В таблиці №10 представлені врожайні дані сої, одержанні в польових дослідах з використанням гербіциду трефлан у дозі 2 і 3 кг/га д.р. по фону інокульованих і неокульованих рослин. Збільшення врожаю насіння, свідчить про ефективність препаратів бульбочкових бактерій в різних регіонах. В той же час у всіх дослідах трефлан негативно діяв на азотофіксацію. Аналогічні дані були одержані у вегетаційних дослідах у НДІ сільськогосподарської мікробіолоїї (Росія), Ю.В. Круглов, А.М. Параменская, Н.И. Чоботарь (1984). Трефлан негативно діє на утворення бульбочок, особливо в початкові фази розвитку сої, знижує біохімічну активність і як наслідок – низька симбіотична активність фіксації азоту (Н.И. Чеботарь, 1979).
Результати оцінки стану симбіотичної системи у сої при обробці гербіцидами приведені в талиці №11. Вони свідчать про негативну дію гербіцидів на активність симбіотичної азотофіксації у сої. Необхідно підкреслити, що негативна дія цих гербіцидів на соєво-ризобіальний симбіоз не завжди супроводжувалась зниженням врожаю. Поясненням являється те, що посіви залишаються без бур'янів і значно поліпшується мінеральне живлення рослин. В той же час стає зрозумілим, що доля біологічного азоту в урожаї при цьому значно зменшується, а значить, цінність бобової культури як азотофіксатора суттєво знижується.
Таблиця №11.
Стан симбіотичної системи і продуктивності сої при нітрогенізації і використанні гербіцидів (Пароменская Л.Н., Кожемякова А.Л., Чеботирь Н.И., Лисовский Л.А., 1987).
| Варіанти | Активність нітрогенезу | Урожайність зерна | ||
| НмольС2Н2 / 1 рослину | % від контролю | Ц\га | % від контролю | |
| Контроль (без гербіциду) | 22,2 | - | 17,8 | - |
| Трифлан (1кг/га) | 11,7 | 52,7 | 18,5 | 104 |
| Сенкор (0,3кг/га) | 19,1 | 86,0 | 18,9 | 106 |
| Базагран (1,5 кг/га) | 16,0 | 72,0 | 13,3 | 78 |
| Дуал (2,5 кг/га) | 12,5 | 56,3 | 18,3 | 103 |
Таким чином, обробка посівів сої гербіцидами може суттєво знизити ефективність використання бульбочкових бактерій. Тому необхідні правильний підбір гербіцидів не токсичних у відношенні бобово-ризобіального симбіозу і розробка заходів, які знижують негативну дію їх використання.