
Агроэкологическое обоснование эффективности ландшафтных систем земледелия в центральном черноземье
| Вид материала | Автореферат диссертации |
- Агроэкологическое обоснование систем применения удобрений в севооборотах на дерново-подзолистых, 1334.17kb.
- Программа дисциплины «история и методология систем земледелия», 143.31kb.
- Тема Современные технологии орошаемого земледелия и полива, 186.16kb.
- Н. Г. Осенний 2012 г. Расписание, 25.09kb.
- 3 Исправьте все стилистические, грамматические, орфографические и пунктуационные ошибки,, 360.88kb.
- Школу экологического органического живого земледелия ведет знаменитый агроном, 36.09kb.
- Состояние ландшафтных систем и их охрана в зоне белореченского химзавода, 772.75kb.
- Технологии земледелия и растениеводства: региональный опыт, 495.68kb.
- Аннотация рабочей программы дисциплины «Земледелие» наименование дисциплины для направления, 36.44kb.
- Ое хозяйство с помощью мелиорации и поливного земледелия, началом строительства в России, 115.22kb.
Биоэнергетический потенциал агроландшафтов и эффективность использования природных и антропогенных ресурсов. Возрастающие затраты на производство единицы сельскохозяйственной продукции обусловливают задачу ресурсо- и энергосбережения, а также оценку коэффициента полезного действия (КПД) использования природных и антропогенных ресурсов при различных вариантах развития сельскохозяйственного производства. Несмотря на важность экономических показателей результативности производства, наиболее объективный анализ эффективности, характеризующий интенсивность процесса связывания энергии агроэкосистемами – энергетический. Он позволяет измерить в сопоставимых единицах затраты живого и прошлого труда, ресурсов различного происхождения, не зависит от политики цен и т.д.
Количественно оценить уровень использования природных и антропогенных ресурсов, прогнозировать направленность развития агроэкосистем позволяет методика ресурсно-экологической оценки на биоэнергетической основе (Володин и др., 1999). В соответствии с этой методикой нами рассчитаны биоэнергетические и эколого-экономические показатели (табл. 7).
Таблица 7 – Биоэнергетический потенциал и показатели ресурсно-экологической оценки агроландшафтов
| Показатель | Агроландшафт | |
| без ЛСЗ | с ЛСЗ | |
| Биоэнергетический потенциал территории, ГДж/га | 2544,1 | 2878,2 |
| Биоэнергетические показатели Коэффициент использования энергии ФАРвп Коэффициент использования органического вещества почвы Коэффициент использования совокупной антропогенной энергии | 4,4·10-3 2,9·10-2 6,0 | 6,3·10-3 3,9·10-2 8,0 |
| Эколого-экономические показатели Показатель производительности агроэкосистемы на единицу денежных затрат, МДж-день / руб. Показатель производительности агроэкосистемы на единицу трудовых затрат, МДж-день / чел.-час. | 6,0·10-2 52,3 | 7,9·10-2 59,6 |
Из таблицы видно, что все биоэнергетические и эколого-экономические показатели агроландшафтов с освоенной ландшафтной системой земледелия выше, чем без нее. Превышения составляют от 14,0 % (производительность на единицу трудовых затрат) до 43,2 % (коэффициент использования энергии ФАР). Таким образом, преобразование агроэкосистемы на ландшафтной основе повышает эффективность использования природных и антропогенных ресурсов, приводит к значительному энерго- и ресурсосбережению.
Освоение ландшафтных систем земледелия привело к увеличению коэффициента использования ФАР и ассимиляционной способности агроэкосистем. Повышение ассимиляционного потенциала преобразованных на ландшафтной основе агроэкосистем позволяет рассматривать их, в отличие от традиционных, не как источник СО2 в атмосфере, а в качестве резервуара депонированного углерода, емкость которого может достигать 0,71 т/га в год. Поскольку результирующая величина накопления гумуса почвами Красногвардейского района от 0,02 % (абс.) содержания гумуса в год в целом по району до 0,05 % (абс.) и более в отдельных случаях определяет ежегодное депонирование углерода пахотным слоем почвы в объеме 0,26-0,64 т/га. Вклад защитных насаждений агроландшафтов района оценивается в 0,07 т/га в год.
Таким образом, ландшафтные системы земледелия явились своего рода пусковым механизмом цепной реакции, приводящей к положительным результатам в природных и производственных процессах.
4 Взаимоотношения между компонентами энтомоценоза в антропогенно сформированных агроландшафтах
Основные объекты и плотность популяций вредных и полезных видов насекомых в системе ландшафтного земледелия. Ввзаимоотношения полезных и вредных насекомых рассматривались нами в целом в агроландшафте, на территории всего Красногвардейского полигона с анализом сообществ насекомых, как в лесополосах, так и в травостое культур.
Доминирующее положение в энтомоценозе вредных видов по численности особей занимали трипсы, относительное обилие которых достигало 93 % (рис. 1). Причем установлено увеличение доли объекта в энтомокомплексе. На это указывают показатели плотности трипсов в многолетней динамике. Если в 2007 г. численность вида составила 37,3 экз./м2, то в 2008 г. – уже 59,0, а в 2009 г. – 96,7 экз./м2 в целом по территории полигона. Очевидно, это предопределено чрезмерно возросшей долей зерновых колосовых в посевах. Эти объекты чаще других могут создавать необходимость дополнительного антропогенного вмешательства. Остальные виды вредных насекомых (злаковые мухи, тли, галлицы, цикадки, настоящие щитники и клопы-слепняки, пьявицы, пилильщики, долгоносики и другие) практически за время исследований не достигали экономически опасного уровня.
Из условно преобладающих видов злаковые тли имели наибольшее значение в заселенности фитоценозов полигона. Их доля от общей численности фитофагов достигала 8 % (2008 г.). В среднем за 2007-2009 гг. плотность тлей в целом по всей исследуемой территории составила 2,7 экз./м2. Что особенно важно, плотность популяции в зерновых ценозах составляла в максимуме – 10 экз./м2 – это низкий показатель для данного вида и он существенно ниже экономического порога вредоносности. Очевидно, на территории полигона сложились экологически не совсем благоприятные условия для тлей.
Практически во всех энтомологических сборах выявлялись злаковые мухи. Несмотря на то, что в отдельных учетах отмечалась плотность 14 экз. /м2, в целом же численность их была небольшой: в среднем за 3 года обследований на всей площади полигона – 1,7 экз. /м2, на полях злаковых культур – 3,5 экз./м2. Следует отметит, что плотность злаковых мух со временем снизилась с 3,3 экз./м2 в 2007 году до 0,9 экз./м2 в 2009 году. Экономический порог вредоносности (3-5 экз./м2) был превзойден злаковыми мухами лишь в 2007 году.

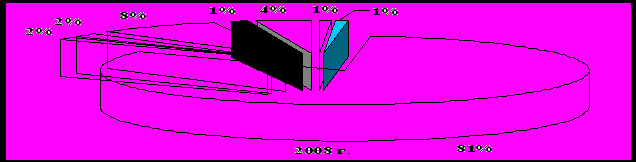


Рисунок 1 – Состав комплекса фитофагов на Красногвардейском полигоне
Для цикадок относительно высокая численность наблюдалась только в 2007 г. и составила 1,5 экз. /м2 при относительном обилии в комплексе фитофагов 3 %. По отдельным учетам была зафиксирована максимальная численность – 14 экз./м2. В целом же за три года для всей изучаемой территории их плотность составила – 0,7 экз./м2. Похожая ситуация складывалась и в отношении галлиц. Лишь в 2007 г. их доля среди комплекса вредных насекомых достигла 4 %, что соответствовало плотности 1,0 экз./м2 в среднем для всего участка, которая в последующие годы снизилась в 33-133 раза. Отдельные учеты фиксировали максимальную плотность равную 23 экз./м2. Моль зерновая имела небольшое увеличение численности в 2008 году, что составило всего 2 % от численности фитофагов – это соответствовало плотности 1,1 экз./м2. В 2007 и 2009 гг. плотность этого вида была ниже в 17-153 раза.
Третья группа малочисленных фитофагов (хлебный клопик, полосатая хлебная блошка, пилильщики, зеленый листовой долгоносик и вредная черепашка), плотность которых изменялась в пределах 0,2-0,3 экз./м2, практически, не имели никакого хозяйственного значения.
Таким образом, за годы исследований энтомокомплекса вредных видов территории полигона только трипсы проявили себя как наиболее многочисленные. Возможно, ощутимый вред могли нанести злаковые мухи в 2007 году. Для подавляющего большинства насекомых этой группы характерны такие плотности популяций, которые не позволяют отнести их к экономически значимым. Во многом это связано с контролем их численности в системе лесных полос со стороны природных врагов.
Среди энтомофагов самыми многочисленными были пауки – их относительное обилие составляло 49-55 % сообщества полезных видов (рис. 2) при их средней численности 3,8 экз./м2, в отдельных учетах фиксировали до 34 экз./м2. Они встречались во всех точках учета на полигоне. Второе место по значимости в комплексе энтомофагов занимали перепончатокрылые паразиты. Их доля в период исследований составляла от 30 до 36 %. Годовое изменение численности видов имела положительную тенденцию – от 1,4 до 2,9 экз./м2 и в среднем составила 2,3 экз./м2.
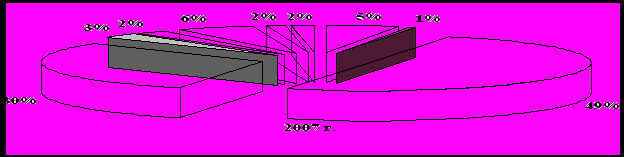
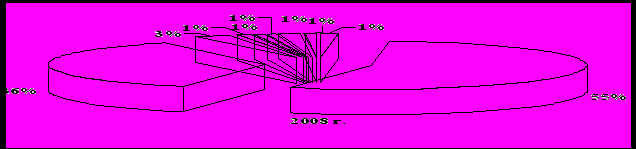
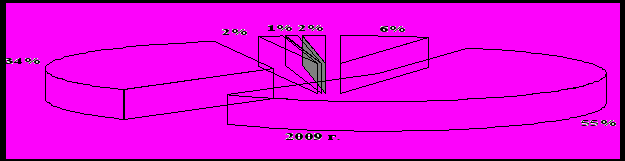

Рисунок 2 – Основой состав энтомофагов на Красногвардейском полигоне
Широко распространенными, играющими существенную роль в регулировании численности фитофагов, были также мягкотелки и божьи коровки, как наиболее активные и подвижные энтомофаги. Относительное обилие этих групп видов в разные годы достигало 6 %. На уровне 1-3 % варьировала доля среди полезных членистоногих таких хищников как мухи-журчалки и златоглазки. Реже, в единичных экземплярах встречались такие энтомофаги как хищные клопы-охотники набис и ориус, верблюдки, жужжалы, муха ктырь, полосатый трипс и стафилиниды. Однако их значение не следует недооценивать.
Соотношение популяций фитофагов и энтомофагов. Анализ распределения энтомофауны по элементам ландшафта выявил следующие особенности. В поле фитофагов в количественном отношении было больше, чем в лесной полосе в среднем в 7 раз, что вполне объяснимо пищевыми предпочтениями этой группы насекомых. Тогда как энтомофагов, наоборот, в лесной полосе было больше, чем в межполосном пространстве. Однако, разница в данном случае – 1,7 раз – была не столь контрастной, как в отношении фитофагов.
Следует отметить, что наблюдается пространственная дифференциация энтомокомплекса не только между лесными полосами и межполосным пространством, но и среди компонентов рассматриваемых фитоценозов. Например, заселенность фитофагами и энтомофагами опушек лесных полос значительно – на 31-48 % выше, чем их внутренней части. Очевидно, для фитофагов это связано с наличием большей массы травостоя (3,92 т/га) на опушках по сравнению с внутренней частью (1,64 т/га) лесных полос. Второй немаловажный фактор влияния на заселенность опушек фитофагами определяется сельскохозяйственными культурами на полях, прилегающих к лесным полосам.
Что касается агроценозов, то здесь ситуация не столь однозначна. Если рассматривать всю совокупность фитофагов, то их плотность в центре полей выше, чем в 15-метровых краевых зонах в среднем на 29-35 %. Такое распределение вредных насекомых напрямую связано с плотностью самых массовых вредителей в изучаемой экосистеме – трипсов. Другие фитофаги такие, как тли, цикадки, хлебный клопик, в отдельные годы злаковые мухи и пилильщики, предпочитают селиться в приопушечной зоне полей. В отношении остальных насекомых вследствие их малочисленности определенных закономерностей в распространении установить не удалось. Плотность энтомофагов в среднем в центре межполосных пространств была ниже, чем в прилегающих к лесным полосам зонах на 57-223 %.
Как показывают наши исследования, на распространение энтомофауны существенное значение оказывает экспозиция склона. Причем и для фитофагов, и для энтомофагов направление влияния совпадает, то есть численность этих групп насекомых на южном склоне превосходит таковую на северном. Для фитофагов эта разница в лесных полосах составляет 65 %, в агроценозах – 24 %. Энтомофагов в лесных полосах южного склона по сравнению с северным больше на 95 %, в межполосном пространстве – на 126 %.
Обращает на себя внимание ситуация складывающаяся с заселением энтомофагов приопушечных зон агроценозов южного и северного склонов – она зеркальна, т.е. на южном склоне энтомофаги предпочитают краевую зону с северной стороны лесной полосы, а на северном склоне – наоборот. Коэффициент корреляции (r) между численностью вредных и полезных насекомых на этих частях полей равен 0,99. Высокое его значение свидетельствует о тесной связи в трофической цепи «хищник – жертва».
Результаты трехлетних исследований позволяют также утверждать, что энтомоценозы агроландшафтов – это довольно мобильные сообщества и являются трансполевыми комплексами. Об этом свидетельствует высокие значения коэффициента корреляции r = 0,98-0,99 между вредными и полезными видами насекомых как в лесных полосах, так и на межполосном пространстве. Но если размещение фитофагов на полях в большинстве случаев зависело от вида возделываемой культуры, то энтомофаги распределялись по посевам культур равномернее фитофагов. Это может свидетельствовать о том, что пространство полигона для полезных видов является единым.
Большинство паразитов и хищников, выявленных нами в результате энтомологических сборов – многоядны. По этой причине, а также вследствие различного отношения насекомых к агроэкологическим условиям оценивался суммарный эффект всех взаимодействующих видов.
Обращает на себя внимание то, что соотношение фитофаг : энтомофаг намного, в среднем в 12 раз, шире в межполосном пространстве, чем в лесополосе (табл. 8). Так, в лесной полосе это соотношение 2,7 : 1, а на поле – 32,2 : 1. Эта тенденция верна как для южного склона (в 12 раз), так и для северного (в 18 раз). Причем сравнение данных по склонам разных экспозиций показывает, что это соотношение для лесополос на южном склоне в 1,2 раза меньше, чем на северном. Такие территориальные отличия наблюдаются и для агроценозов культур, но уже с разницей в 1,8 раз. Это позволяет сделать вывод, что на склонах южной экспозиции складываются более благоприятные условия для естественного регулирования численности вредных организмов.
Таблица 8 – Соотношение фитофагов и энтомофагов в зависимости от структурных элементов агроландшафта (в среднем за 2007-2009 гг.)
| Элементы агроландшафта | Южный склон | Северный склон | Водосбор | |
| Лесные полосы | 1. южная опушка | 3,1 | 2,9 | 2,8 |
| 2. внутри | 2,8 | 2,8 | 2,8 | |
| 3. северная опушка | 2,2 | 3,7 | 2,5 | |
| Среднее | 2,7 | 3,2 | 2,7 | |
| Агро-ценозы | 1. 15 м от южной границы поля | 21,7 | 94,1 | 21,2 |
| 2. центр поля | 57,4 | 110,4 | 61,7 | |
| 3.15 м от северн. границы поля | 28,8 | 37,1 | 29,1 | |
| Среднее | 31,8 | 57,0 | 32,2 | |
Анализ соотношения вредных и полезных видов по разным частям поля показывает, что оно больше в центре поля, чем в шлейфовых зонах в среднем в 2,1-2,9 раз. Причем напряженность отношений в энтомокомплексе центральной части агроценозов южного склона выше, чем северного. Прессинг со стороны энтомофагов в центре полей на склоне северной экспозиции ослабевает в 1,9 раз.
Оценивая соотношение фитофагов и энтомофагов в приопушечных зонах полей южного склона и вблизи южной опушки лесных полос на северном склоне, можно заключить, что уровень эффективности энтомофагов в них довольно высок. Очевидно, что около лесных полос создаются лучшие условия для естественного контроля численности популяций вредителей. Это может служить основой для корректировки химических обработок. Для некоторых видов фитофагов даже при численности вредителей близкой к традиционно используемым экономическим порогам вредоносности может быть нецелесообразным краевое опрыскивание посевов. То есть ландшафтные системы земледелия при всех своих положительных эффектах могут обеспечивать еще и определенное повышение экологической безопасности при проведении защитных мероприятий на основе использования химических средств защиты растений.
Полученная количественная оценка взаимоотношений в энтомокомплексе обследованной территории является прямым следствием влияния активности трипсов – самых мелких, но многочисленных среди фитофагов – для которых существует необходимость применения дополнительных защитных мероприятий. Максимальное соотношение между вредными (без учета трипсов) и полезными насекомыми составляет в среднем – 4,8:1. Это свидетельствует в пользу того, что на территории полигона между вредными видами, кроме трипсов, и их естественными врагами сложились равновесные взаимоотношения.
5 ВИДОВОЕ РАЗНООБРАЗИЕ РАСТИТЕЛЬНОСТИ В ЛАНДШАФТНЫХ СИСТЕМАХ ЗЕМЛЕДЕЛИЯ
До сих пор не существует единого мнения, в какой степени видовое разнообразие определяет устойчивость экосистемы. Однако большинство исследователей показывают положительную связь между разнообразием сообществ и стабильностью их экологических функций. При ведении интенсивного сельскохозяйственного производства, при котором выращивается ограниченное количество сельскохозяйственных культур, имеющих коммерческую ценность, мы стоим перед необходимостью решения острой проблемы сохранения агроландшафтов в целом и сохранения биоразнообразия в частности.
Видовое разнообразие растительности антропогенно сформированных агроландшафтов. Для изучения влияния ЛСЗ на биоразнообразие растительности преобразованных территорий был выбран модельный комплекс – Красногвардейский стационар, включающий практически все элементы экологического каркаса ландшафтных систем: разновозрастные лесные полосы (1947 г. и 1983-1985 гг. закладки) и межполосное пространство, занятое полями севооборотов, а также участки овражно-балочных комплексов и сплошного облесения, являющихся основным резерватом видов, которые потенциально могут внедряться на участках с интенсивным ведением сельскохозяйственного производства (поля, лесополосы, участки залужения и др.).
До начала освоения ЛСЗ видовое разнообразие растительности стационара соответствовало уровню видового разнообразия пашни с прилегающей к ней территорией пастбищных угодий с ограниченным набором сельскохозяйственных культур и сопутствующих им видов травянистой растительности. Посадка контурных лесополос, мероприятия по сплошному облесению участков, наиболее опасных в эрозионном отношении, привели к значительному изменению видового разнообразия растительности стационара. Отмечена ее дифференциация в зависимости от ландшафтных условий и сформированных сообществ.
В ходе исследований флористического разнообразия было выявлено 264 вида из 173 родов и 59 семейств. Сравнительный анализ изучаемых сообществ показал, что ведущие места в ранжировке по количеству видов занимают семейства Fabaceae (30), Asteraceae (29), Poaceae (24), Lamiaceae (18), Rosaceae (14), Brassicaceae (13), Apiaceae (13), Caryophyllaceae (11). На их долю приходится 57,3 % от общего числа видов. Остальные семейства имеют небольшое число видов, а 25 из них представлены одним видом. В целом набор ведущих семейств, особенно первых десяти, совпадает, в основном, с таковым в Центральном Черноземье (Камышев, 1978). Черты своеобразия систематической структуры исследуемой флоры проявляются в заметно более высоком положении в спектре семейства Fabaceae и более низком семейства Brassicaceae.
Структура флоры и типы растительности агроландшафта. Одним из важнейших показателей растительного покрова является состав и соотношение разных жизненных форм растений в фитоценозах. Самой многочисленной из жизненных форм стационара является травянистая флора (235 вида), включающая одно-двулетники – 28 % от общего числа видов и многолетники (61 %). Травянистая растительность относится к 46 семействам однодольных и двудольных растений. Наиболее многочисленными по видовому составу семействами у однодольных оказались мятликовые (24 вида), у двудольных – бобовые (30) и астровые (29). Доля древесной и кустарниковой растительности составляет 6 и 5 %, соответственно.
Специфику растительного покрова определяют 9 основных фитоценотических групп, которые представлены типично степными видами – их 57, лугово-степными – 45 видов, лесными – 40, сорно-сегетальными и сорно-рудеральными видами – 69, луговыми – 25, опушечно-лесными – 23, адвинтивными – 4 вида и 1 вид следует отнести к кальцефильной группе. На долю сорных растений приходится 26,1 %. Сравнительно большое число видов сорных растений является показателем существенной антропогенной трансформации ландшафта.
При анализе видового состава сообществ с точки зрения отношения растений к влаге выделены две основные группы – мезофиты и ксерофиты, а также промежуточные между ними категории. Доля участия мезофитов (включая ксеромезофиты) достаточно велика – 67 %. Очевидно это является следствием влияния относительно влажного мезоклимата, сформированного лесной формацией данной территории.
Лесополосы и примыкающие к ним агроценозы являются единым биоценотическим комплексом, где существует определенная система пространственно-временных отношений между флорой, фауной и окружающей их средой. Немаловажное значение для стабилизации фитосанитарного состояния агроценозов имеет наличие энтомофильных растений. Среди изучаемой флоры энтомофильные растения преобладают – 218 видов, или 82,6 % от общей численности видов.
Ядовитые растения составляют около 5,3 % от общего видового состава. Некоторые из них являются лекарственными. В целом лекарственных растений насчитывается 165 видов – это более 60 % всего видового состава.
На исследуемой территории особое внимание уделялось выявлению во флористическом составе редких, исчезающих и эндемичных видов флоры. Из установленных нами видов растений на Красногвардейском полигоне одинадцать входят в Красную книгу Белгородской области: тюльпан Биберштейна, осока низкая, любка двулистная, дремлик морозниковый, первоцвет весенний, лен желтый, лен украинский, лен многолетний, триния многостебельная, синюха голубая, синяк русский или Румянка. А также четыре вида кандидата на включение в Красную книгу Белгородской области: лук медвежий, или черемша, шиповник Юндзилла, ракитник австрийский и колокольчик персиколистный.
В общее достаточно высокое разнообразие изучаемого стационара вносят вклад сообщества, формирующие различные элементы агроландшафта (пашня, естественные кормовые угодья, залежь, защитные лесные насаждения, лесные массивы). Однако участие этих растительных сообществ неравнозначно.
Наибольшее видовое разнообразие отмечено в овражно-балочном комплексе (182 вида), прилегающем вплотную к участку байрачного леса, и в лесной полосе закладки 1947 года (133 вида). Остальные компоненты данного агроландшафта имеют значительно меньшее видовое богатство и располагаются в следующем порядке по мере его убывания: залежь (105 видов), молодые лесные полосы (90) и поля севооборотов (83 вида).
Вследствие отмечаемого сходства растительных сообществ молодых лесных полос (особенно их доминантных видов), очевидно вызванного их сходным возрастом (1983-1985 гг. закладки), было принято решение учитывать их под одним названием «молодые лесные полосы». Поля севооборотов, где происходит перемещение комплексов организмов вслед за движением по территории соответствующей сельскохозяйственной культуры, мы рассматриваем, как единое сообщество и их флористические списки объединяем в один.
Особый интерес для агроэкосистем представляет анализ процессов, происходящих на участках с наибольшей динамикой почвенно-растительной системы.
По соотношению эколого-ценотических групп видов в составе растительного покрова возможно получение одной из оценок структурного разнообразия растительности (рис. 3).
В старовозрастной лесной полосе, также как и в молодых лесных насаждениях отмечается значительное превосходство лесной и опушечно-лесной флоры по видовому богатству и по степени ее участия в растительных сообществах по сравнению с фитоценозами поля и залежи. Если в лесных полосах доля этой фитоценотической группы составляет более 32 %, то для поля и залежи она равна 6 и 11 %, соответственно. Следует отметить, однако, что по количеству лесных видов растительности 63-летняя лесополоса в 2 раза превосходит молодые лесополосы.

Примечание – обозначение эколого-ценотических групп:
Рисунок 3 – Эколого-ценотические спектры видового богатства в сообществах
Красногвардейского полигона, %
Обращает на себя внимание и предсказуемое отличие этих типов фитоценозов по наличию сорного компонента, который преобладает в растительных сообществах поля (54 %) и залежи (41 %). Здесь доля сегетальной и рудеральной флоры в 1,5-2,0 раза выше, чем в лесополосах.
Степная эколого-ценотическая группа, включающая собственно степную, лугово-степную и луговую формации, довольно широко представлена во всех компонентах Красногвардейского полигона. Это связано с тем, что виды этой группы имеют широкий экологический оптимум обитания. Наименьшая ее доля в молодых насаждениях (33 %) и максимальная во флоре залежи (48 %). Однако в 63-летней лесной полосе большая часть этой группы приходится на луговые и лугово-степные виды, тогда как на залежи преобладают степные виды.
На рисунке 4 показано распределение видов по экологическим группам в различных сообществах.
А Б
В Г
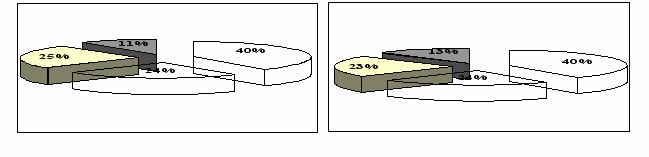

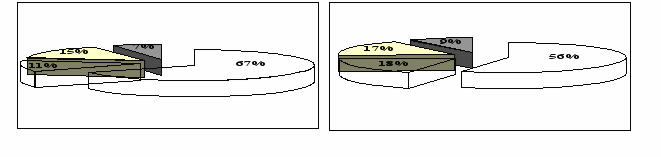
Рисунок 4 – Распределение видового богатства растительных сообществ Красногвардейского стационара по экологическим группам: А – старовозрастная лесополоса, Б – молодые лесополосы, В – поле, Г – залежь.
Определяющим фактором было влияние лесополос. В системе защитных насаждений увеличивается влажность прилегающих территорий (полей) и это отражается на экологической структуре флоры. Наибольшей мезофитностью обладала растительность старовозрастной лесополосы и молодых защитных насаждений. Под дубовой лесополосой доля мезофитных видов составляет две трети от общего их количества. В молодых лесополосах она была на 11 % меньше, хотя тоже значительна. Совокупное участие мезофитов и ксеромезофитов в этих двух флорокомплексах было примерно одинаковым и составляло 74-78 %. Наибольшая доля ксерофитов отмечалась на поле и залежных участках – 36 %. Однако и в межполосном пространстве мезофиты (40 %) совместно с ксеромезофитами (24 %), занимали большую долю в общем видовом разнообразии поля и залежи.
Таким образом, на данной территории отчетливо отразилось влияние лесной формации на формировании экологической структуры всей растительности, выразившееся в преобладании мезофитных, более влаголюбивых видов.
Изучение жизненных форм, их динамики исключительно важно для познания закономерностей приспособления популяций и организмов к тем или иным условиям среды. Распределение растений по жизненным формам может служить показателем направленности сукцессионных процессов в каждом сообществе и в пределах всего изучаемого полигона (рис. 5).
А
Б
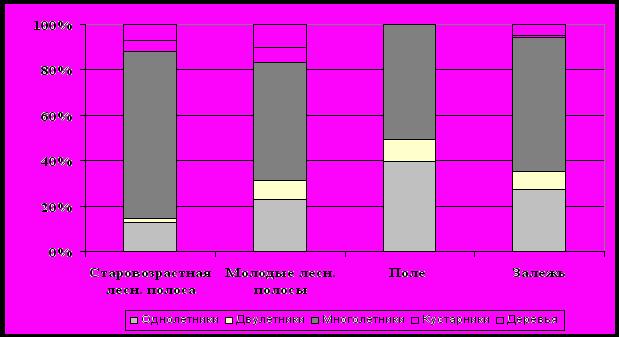
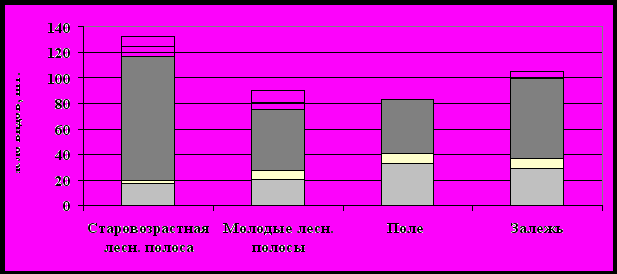
Рисунок 5 – Жизненные формы растений Красногвардейского полигона.
А – общее число видов по жизненным формам и сообществам.
Б – то же в процентах.
Анализ жизненных форм показал, что для всех типов ценозов характерно преобладание травянистой растительности – от 83 % в молодых лесных полосах до 100 % на поле. По видовому богатству она максимальная в 63-летней лесной полосе (117 видов). Из травянистой флоры самыми многовидовыми являются многолетники. Это типично для всех компонентов данного комплекса. Однако для растительных сообществ поля и залежи существенная роль принадлежит малолетникам. Доля одно- и двулетников на поле составляет 50 %, тогда как в старовозрастной лесной полосе только 15 %. Молодые лесополосы и залежь по участию малолетников занимают промежуточное положение – 31 и 36 %, соответственно. Наибольшим разнообразием древесной растительности, определяемым как видовым сортиментом высаженных пород, так и внедренных видов, естественно обладают защитные лесные насаждения.
В целом различия между изучаемыми компонентами Красногвардейского полигона довольно значительны, о чем свидетельствует коэффициент Жаккара, изменяющийся в пределах от 0,20 до 0,35. Это проявляется за счет расширения видового обилия, особенно в 63-летней лесополосе, увеличения лесных и опушечно-лесных видов в защитных насаждениях, а также сокращения в них видов сорной растительности. Снижение доли однолетних и двулетних растений в лесополосах существенно, особенно по сравнению с полем.
Трансформация травянистой флоры под влиянием древесных пород. До сих пор не существует четкого представления о количественных характеристиках биоценотических связей между компонентами агроландшафта, протекающих в нем процессов и явлений, их взаимовлияния и взаимодействия, определяющего, в том числе фитосанитарное состояние агроэкосистемы в целом.
С целью прогнозирования влияния лесных полос на засоренность прилегающих полей севооборотов, необходимо учитывать происходящие во времени процессы трансформации растительности, особенно наиболее динамичной травянистой. Растительный покров Красногвардейского полигона многообразен. Травостой различных составляющих его ценоэлементов отличается сложной неоднородной морфологической структурой. Фитоценотическая роль отдельных видов неравноценна.
Растительность поля представляет собой многокомпонентное сообщество, в которое помимо культурного растения в зависимости от его вида и технологии возделывания входят виды естественной растительности. Наши исследования произрастающих на полях севооборотов видов показали, что больше половины из них – это сорная флора. Малолетние растения обильно представлены именно в этом флорокомплексе – 50 % видового состава. Фитоценозы поля при исключении культурного компонента (это хорошо видно на плохо обрабатываемом поле пара) характеризуются случайным сочетанием растений, слабым их взаимовлиянием, отсутствием сомкнутого растительного покрова и хорошо подходят под описание, данное А.Г. Вороновым (1973), группировки начальной стадии восстановительной сукцессии. Здесь имеют преимущество так называемые эксплеренты, способные быстро захватывать освобождающуюся территорию.
Наши исследования показали, что в травянистом сообществе 7-8-летних контурных лесных полос в ЗАО «Должанское» Вейделевского района растительный покров в надземной части не сомкнут (проективное покрытие 75-80 %), однако растения располагаются значительно теснее – обычен групповой характер распределения растений: вокруг особи, давшей семена, развивается ее потомство. Такой фитоценоз характерен для бурьянистой стадии залежи.
Накопленный в прежние годы в пахотном слое банк семян позволяет сохранить представительство сегетальных и рудеральных видов на залежном участке Красногвардейского полигона. Тем не менее, характерным для растительного покрова залежи является сокращение доли сорной флоры до 41%, а доли однолетних и двулетних видов до 36 % по сравнению с полями севооборота. Травостой густой и высокий. Проективное покрытие – 100 %. Здесь наблюдается более сложная растительная группировка. Наряду со злаково-бобово-разнотравными сообществами нами выделяются разнотравно-злаковые растительные сообщества, в которых фон составляют пырей ползучий и мятлик луговой. Очевидно, это связано с бóльшим возрастом залежи – 12 лет.
В 25-27-летних контурных лесных полосах Красногвардейского полигона наблюдаются характерные признаки (все пространство лесополос занято разнотравно-пырейными, чисто пырейными и мятликовыми ассоциациями) для стадии длиннокорневищных и рыхлодерновинных злаков восстановительной сукцессии. Доминирующая роль в составе растительных сообществ остается на протяжении довольно длительного периода за двумя злаками – мятлик (доминант) и пырей (содоминант). Сходные сообщества мы отмечали при обследовании в соседних районах (Алексеевском, Валуйском, Вейделевском) лесных полос и другого породного состава (например, ясень, вяз), но того же или близкого возраста.
Особенностью 63-летней лесной полосы является большое разнообразие травянистых сообществ. Встречаются сообщества характерные как для ранних этапов сукцессии (пырейно-разнотравные, разнотравно-злаковые, пырейно-мятликовые), так и характерные для лесных климаксовых стадий. На видовой состав старовозрастной лесной полосы, несомненно, оказал влияние прилегающий к ней дубовый лес. Однако видовое разнообразие полосы несравнимо богаче, чем видовое богатство леса.
Кроме того, наши исследования показали, что в старовозрастных лесных полосах, особенно в тех, которые имеют значительную ширину и, самое главное, высокую степень сомкнутости крон, очень часто реализуется другой сценарий развития травяного покрова. В многорядных с сомкнутым пологом полосах различного породного состава травянистая флора отсутствует вообще или произрастает разрежено, единичными экземплярами.
Метод трансформации пространственных рядов во временные позволил рассматривать растительные сообщества Красногвардейского полигона (поля севооборотов, участки залежи, 25-27-летние лесные насаждения и 63-летняя дубовая лесополоса) как стадии единого динамического процесса – восстановительной сукцессии.
Именно агроценозы с механической обработкой почвы, которая удерживает их на первой стадии сукцессии, являются источником возобновления сорной флоры в агроландшафтах. Тогда как в лесных полосах через определенный период времени травяная флора проходит сукцессионное развитие и достигает климаксовой стадии, растительные сообщества которой не угрожают ухудшением фитосанитарного состояния агроценоза. Поскольку виды, входящие в эти сообщества характеризуются теневыносливостью, медленным ростом, низкой плодовитостью и не способны выживать в условиях поля. Это, очевидно, прогнозируемая перспектива развития флористического состава экологического каркаса ландшафтных систем земледелия. Контроль за численностью сорняков во многом зависит от культуры земледелия, а не от прилегающих лесных насаждений.