
Ю. И. Научение и память: системная перспектива
| Вид материала | Литература |
- Книга канадского автора. Учебник общей психологии с основами физиологии высшей нервной, 8180.96kb.
- Научающе-бихевиоральное направление в теории личности по б. Ф. Скиннеру, 676.83kb.
- Конкурс «Полярная перспектива 2011» Оконкурсе. Фотоконкурс «Полярная перспектива», 74.49kb.
- Лекция 5 Внутренняя память, 178.2kb.
- 1. Изучение поведения история и методы 13 Глава 1 Что такое поведение, 7795.15kb.
- Форма, часы, 52.45kb.
- Память память, 1160.48kb.
- Пояснительная записка к системе учебников «Перспектива» (для 1−4 классов общеобразовательных, 1411.93kb.
- Индивидуальное развитие памяти у людей, 357.08kb.
- Человеческого духа, 1692.85kb.
Александров Ю.И.
НАУЧЕНИЕ И ПАМЯТЬ: СИСТЕМНАЯ ПЕРСПЕКТИВА
Александров Ю.И. Научение и память: системная перспектива // Вторые симоновские чтения/Под ред. Шевлев И.А. и др. М.: Изд. РАН, 2004. C. 3-51.
Содержание
Введение
Детерминация активности нейрона
От концепции нейрона сумматора и проводника возбуждения к концепции его интегративной деятельности
Нейрон как «организм» в организме
Объединение нейронов в систему как способ достижения результата на уровне целого организма и отдельной клетки
«Действие» нейрона
Множественность «нейротрансмиттеров»
Научение как образование «следов» и как системогенез
Системно-селекционная концепция научения
Актуализация памяти при реализации поведения
Неонейрогенез
Консолидация памяти: от формирования и упрочения «следов» посредством повышения эффективности синапсов к системогенезу
Реконсолидация при реактивации памяти и при научении
Консолидация с системной точки зрения
В начале формирования памяти: экспрессия «ранних» генов как показатель рассогласования
От рассогласования через согласование к консолидации
Элиминация нейронов как «альтруистичный суицид»
Долговременная потенциация: полезный артефакт?
Выдвижение и селекция гипотез при научении
Индивидуальное развитие как последовательность системогенезов
Заключение
Благодарность
Литература
ВВЕДЕНИЕ
Проблема научения и памяти принадлежит к числу наиболее интенсивно разрабатываемых. Являясь главной для множества исследователей самого разного профиля: в молекулярной биологии, нейрофизиологии, психологии, она, однако, важна не только для них, но, пожалуй, для всех исследователей упомянутых и ряда других областей знания.
Это подчеркивал и П.В. Симонов, который называл проблему «механизмов замыкания условных связей» «центральной» [30, с.35].
Чему бы ни была посвящена конкретная работа, она имеет дело с индивидом, у которого есть память. Пока индивид жив, ее содержание модифицируется за счет приобретения новых форм поведения. Но не только. Хотя особенно глубокая реорганизация памяти имеет место, как правило, при формировании новых поведенческих актов, но и, как будет специально сказано ниже, реализация приобретенной ранее памяти есть также «повторение без повторения» [15]. Таким образом, само существование индивида есть непрерывная модификация, а точнее - непрерывное созидание памяти.
Как это следует из названия, в настоящем сообщении проблема научения и памяти будет рассматриваться с позиций системного подхода, развиваемого в течение многих лет в системной психофизиологии, базирующейся на теории функциональных систем. Позиции системной психофизиологии, будучи оригинальными, не остаются, однако, островком, изолированным от остальной науки. Содержание целого ряда теоретических и экспериментальных статей, появившихся за последнее время в авторитетных научных журналах, приводит к заключению, что налицо новая фаза движения науки от «стимульного» к «целевому» и «холистическому» детерминизму, к утверждению системности и подчеркиванию активности индивида. Это движение еще не является главным направлением (mainstream) развития науки, но получает все большую поддержку, «официализируется» [79,80,84,100,155,157].
Несмотря на отмеченную тенденцию, и в настоящее время сохраняются существенные различия между системной и традиционной парадигмами. Естественно, подход к экспериментальному изучению закономерностей научения и памяти и к трактовке полученного материала в значительной мере зависит от методологических установок исследователя. Эмпирическое явление превращается в факт, будучи интерпретированным в терминах теории [25,26,42]. (К сожалению, часто имплицитной и потому эклектичной.) Поэтому одно и то же явление может предстать для авторов, придерживающихся разных теорий, в качестве разных фактов.
Вот почему мне представляется важным показать, как выглядят закономерности научения и памяти с позиций системного подхода, и сравнить это взгляд с традиционным.
Из только что сказанного вытекает, что, прежде чем изложить взгляд на проблему научения и памяти с системных позиций, следует сначала описать тот концептуальный аппарат, который используется для интерпретации экспериментального материала в системной парадигме. Однако, принимая во внимание ограничения объема настоящего сообщения, а также то, что на протяжении ряда лет такие описания многократно приводились (см., например, [3,4,6,7,12,37,38]), я остановлюсь лишь на том, что считаю совершенно необходимым для последующего изложения логики системного анализа названной проблемы. Это - понимание нейрона не как проводника возбуждения, а как «организма» в организме.
ДЕТЕРМИНАЦИЯ АКТИВНОСТИ НЕЙРОНА
От концепции нейрона сумматора и проводника возбуждения к концепции его интегративной деятельности
Традиционно нейрон рассматривается как элемент, входящий в проводящую структуру большей или меньшей сложности, например, в рефлекторную дугу. Тогда функцией нейрона оказывается обеспечение проведения возбуждения. При этом в качестве стимула рассматривается пресинаптическая импульсация, а в качестве реакции –импульсация постсинаптического нейрона. То есть нейрон, как и организм, реагирует на стимулы.
Важнейшим событием в развитии теории функциональных систем стало определение системообразующего фактора - результата системы, под которым понимался полезный приспособительный эффект в соотношении организма и среды, достигаемый при реализации системы. Таким образом, в качестве детерминанты поведения в теории функциональных систем рассматривается не прошлое по отношению к поведению событие - стимул, а будущее – результат [12]. Однако теория функциональных систем включала наряду с понятием «результат», и понятие «пусковой стимул». Это сочетание отражало, как я полагаю, эклектичность классического варианта теории. Кажущаяся необходимость использования понятия «пусковой стимул» (чем-то ведь должен «запускаться» поведенческий акт, даже если его организация определяется будущим – результатом, на который этот акт направлен!) отпала при рассмотрении поведенческого акта не изолировано, а как компонента поведенческого континуума: последовательности поведенческих актов, совершаемых индивидом на протяжении его жизни. При этом оказывается, что следующий акт в континууме реализуется после достижения и оценки результата предыдущего акта. Упомянутая оценка - необходимая часть процессов организации следующего акта, которые, таким образом, могут быть рассмотрены как трансформационные или процессы перехода от одного акта к другому. Места для стимула, в том числе и пускового, в континууме нет [37].
Решающий шаг на пути формирования системного подхода к пониманию активности нейрона был сделан П.К.Анохиным [12] . В концепции интегративной деятельности нейрона, предложенной им вместо традиционной «суммационной» концепции, рассматривающей нейрон как сумматор и проводник возбуждения в рефлекторной дуге, генерация потенциалов действия нейроном рассматривалась как обеспечение достижения результата системы и следствие внутринейронных химических процессов. Однако, хотя в этой новой концепции и подчеркивалась роль межклеточных контактов не в проведении возбуждения, а в обмене метаболическими субстратами между клетками, активность нейрона в ней все еще рассматривалась как реакция, следующая в ответ на стимул - импульсацию пресинаптических нейронов.
Нейрон как «организм» в организме
Следующим шагом в развитии системных представлений о детерминации активности нейрона была формулировка В.Б. Швырковым [38] представления о том, что нейрон, как и любая живая клетка, реализует генетическую программу, нуждаясь в метаболитах, поступающих к нему от других клеток. Активность нейрона, как и поведение организма, является не реакцией на прошлое событие, а средством изменения соотношения со средой, «действием», которое обусловливает достижение результата. Последовательность событий в деятельности нейрона становится аналогичной той, которая характеризует активный целенаправленный организм, а его импульсация - аналогичной действию индивида.
Иначе говоря, активность нейрона, как и поведение организма, рассматривается не как реакция, а как средство изменения соотношения со средой, «действие», которое обусловливает устранение несоответствия между «потребностями» и микросредой, в частности, за счет изменения синаптического притока. Это изменение, если оно соответствует текущим метаболическим «потребностям» нейрона, приводит к достижению им «результата» и прекращению активности.
Предполагается, что рассогласование между «потребностями», определяемыми генетически, и реально поступающими метаболитами может иметь место как при генетически обусловленных изменениях метаболизма клетки, так и при изменении притока метаболитов от других клеток.
Таким образом, в рамках данных представлений, нейрон - не «кодирующий элемент», «проводник» или «сумматор», пусть даже очень «умный», а организм в организме, обеспечивающий свои «потребности» за счет метаболитов, поступающих от других элементов.
Представление о клетке как об организме в организме не является новой идеей, сформулированной впервые в системной психофизиологии. Так, более пятидесяти лет назад Ч. Шеррингтон [141, p.73] писал: «Утверждение, что из клеток, составляющих нас, каждая является индивидуальной эгоцентричной жизнью – не просто фраза. Это не просто удобный способ описания. Клетка как компонент тела – не только визуально ограниченный модуль, это отдельная жизнь, сосредоточенная на себе. Она живет собственной жизнью… Клетка – это отдельная жизнь, и наша жизнь, которая, в свою очередь, является отдельной жизнью, всецело состоит из жизней-клеток». Что действительно нового добавлено в описываемой концепции нейрона к этому утверждению – это приведение данного общетеоретического представления об «эгоистичной» клетке, в частности, о нейроне, в соответствие с представлением о системной детерминации ее активности, связываемой с рассогласованием между «потребностями» клетки и метаболическим притоком.
Роль большинства химических соединений, поступающих в «микросреду» клетки, сводится к изменению свойств и скорости синтеза имеющихся в ней белков или к инициации синтеза новых белков. На метаболизм нейрона влияют и нейромедиаторы, выделяемые из терминалей контактирующих с ним нейронов. Соединяясь со «своим» рецептором, медиатор не только изменяет проницаемость ионных каналов, но и оказывает влияние на внутриклеточные процессы, выступая в качестве индуктора внутриклеточных метаболических превращений (см., например, [34]). Однако, и изменение проницаемости ионных каналов, обычно связываемое лишь с модуляцией электрического потенциала нейрона, в свою очередь оказывает существенное влияние и на метаболизм клетки: трансмембранный транспорт метаболитов, поддержание уровня pH внутри клетки [136].
Объединение нейронов в систему как способ достижения результата на уровне целого организма и отдельной клетки
Рассмотрение нейрона как организма в организме соответствует представлениям о значительном сходстве между закономерностями обеспечения жизнедеятельности нейрона и одноклеточного организма [105]. Однако между ними существует и серьезное различие. Одноклеточный организм (как, впрочем, и многоклеточный) может обеспечить свои метаболические потребности за счет лишь собственной активности. Причем эта активность не обязательно вступает в отношения взамосогласования с активностями других организмов. Нейрон же обеспечивает «потребности» своего метаболизма объединяясь с другими элементами организма в функциональную систему. Их взаимосодействие, совместная активность приводит к достижению результата, новому соотношению целостного индивида и среды.
«Извне», на уровне целостного индивида, при анализе его внешнего поведения, результат описывается как определенное соотношение организма и внешней среды, которое прекращает действие, направленное на его достижение, и делает возможной реализацию следующего поведенческого акта. «Изнутри», на уровне отдельных нейронов, достижение результата выступает как удовлетворение метаболических «потребностей» нейронов и прекращает их импульсную активность.
Такой подход к пониманию активности нейронов предполагает применение для ее анализа «предрезультатных» гистограмм. Построение «предрезультатных» гистограмм позволяет обнаружить нейроны, активность которых закономерно увеличивается при реализации поведения, направленного на получение данного результата, и прекращается при его достижении. Рис. 1 иллюстрирует прекращение активаций нейронов цингулятной и антеролатреальной областей коры при достижении результатов инструментального поведения: захват кольца или контакт с педалью (данные экспериментов Аверкина Р.Г. и др. [51,52]).
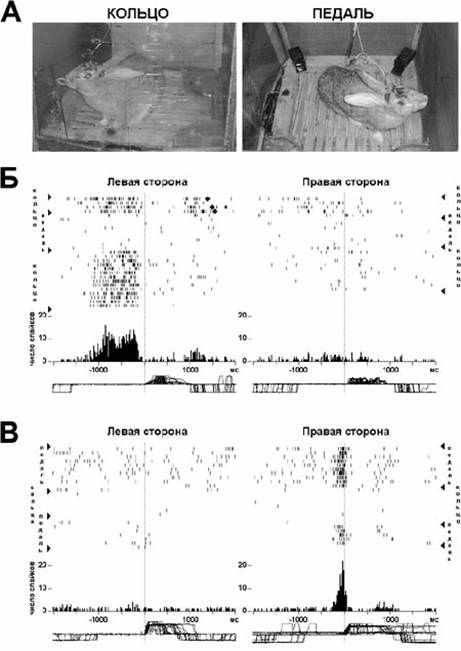
Рис. 1. Активации нейронов цингулятной и антеролатеральной моторной областей коры кроликов, прекращающиеся при достижении результатов инструментального поведения.
Экспериментальная клетка (А и Г), в которой инструментальное пищедобывательное поведение, реализуется животным попеременно способом потягивания за кольцо (А) или нажатием на педаль (Г). Клетка снабжена парой кормушек, автоматически подающихся при нажатии на соответствующие педали (расположенные у той же, что и кормушка, стенки клетки) или при потягивании за соответствующие кольца. Ниже - растры импульсации и гистограммы активности нейронов антеролатеральной (Б и Д) и цингулятной (В и Е) областей коры.
Б – нейрон антеролатеральной коры активировался при подходе, захвате и потягивании обоих колец. Д – нейрон антеролатеральной коры, активировался при контакте с правой, но не левой педалью. Активация при подходе к кольцу и его захвате не появлялась. В – нейрон цингулятной коры активировался при захвате левого, но не правого кольца. Активация при подходе к педалям и нажатии на них не появлялась. Е – нейрон цингулятной коры активировался как при подходе, захвате и потягивании левого кольца, так и при подходе и нажатии на левую педаль.
На В и Д растры и гистограммы построены относительно начала потягивания за кольцо и начала нажатия на педаль, соответственно; на Б и Е растры и гистограммы построены относительно завершения потягивания за кольцо и нажатия на педаль. Вертикальные линии, проходящие через все компоненты фрагментов Б-Е, обозначают момент, от которого производилось построение растров и гистограмм. На растрах вертикальные черточки обозначают отдельные импульсы нейрона, а горизонтальные ряды черточек – последовательность импульсов в отдельном цикле пищедобывательного поведения. Под растрами, кумулятивные гистограммы с шириной канала 20 мс ( для В и Д) и 50 мс (для Б и Е). Внизу –актограммы поведения для всех циклов пищедобывательного поведения, совершенных животным при регистрации импульсации данного нейрона. На актограмме смещение линий вверх соответствует потягиванию кольца или нажатию на педаль, вниз—опусканию морды животного в кормушку. На фрагменте В ромбиками указаны повторные потягивания. На фрагменте Е прямоугольником указано первое потягивания в случае, когда животное в пищедобывательном цикле реализовало несколько раз инструментальный акт.
В литературе имеются данные, которые позволяют связать частоту «предрезультатной» активности с величиной потребности, с одной стороны, и с появлением поведения, направленного на удовлетворение этой потребности, с другой. Показано, что у зависимых от кокаина животных, обученных нажимать на педаль для ведения себе кокаина (через канюлю, вживленную в яремную вену), частота активности нейронов, вовлекающихся в обеспечение инструментального кокаиндобывательного поведения, тем выше, чем ниже концентрация кокаина в организме [121]. Когда частота активности, постепенно нарастая, достигает определенного уровня, развертывается инструментальное поведение, достижение результата которого - введение кокаина, - проявляется в подавлении активности этих нейронов [63,121]. Затем, по прошествии некоторого времени, концентрация кокаина опять понижается, частота активности повышается и реализуется поведение добывания кокаина - цикл повторяется.
«Действие» нейрона
«Действие» нейрона, его импульсная активность, не только влияет на микросреду, но изменяет и сам импульсирующий нейрон. Уже давно было известно, что «следовые», постспайковые процессы (такие как изменение поляризации, ионной проницаемости) играют существенную роль в регуляции чувствительности нейрона к последующему притоку. Позже как на препаратах, так и на бодрствующих животных было показано, что потенциал действия, генерируемый нейроном, распространяется не только в «обычном» направлении - по аксону к другим клеткам, но и в обратном направлении - к дендритам данного нейрона (феномен «обратного распространения», «backpropagation») [60,85]. При этом его чувствительность к синаптическому притоку существенно модифицируется. При сочетании эффектов «обратного распространения» с пресинаптической импульсацией возникают изменения в дистальных дендритах, а также и в теле нейрона.
Имея в виду только что изложенное, активность нейрона, как и поведение индивида, можно рассмотреть как со стороны влияния на окружающую среду (микросреду), так и со стороны модификации активного агента, модификации, соответствующей ожидаемым параметрам эффекта этих влияний и являющейся непременной характеристикой активности. Тогда модификация нейрона вследствие его собственной активности может быть рассмотрена как показатель подготовки нейрона к будущему притоку, связанному с его активностью. Иначе говоря, эти данные свидетельствуют в пользу того, что, давая спайки, нейрон не только обеспечивает необходимый ему метаболический приток, но и подготавливается к его «утилизации» [4].
Множественность «нейротрансмиттеров»
В рамках традиционного подхода к пониманию нейрона как проводника возбуждения закономерно возникает вопрос, сформулированный Kandel E.: «Почему имеются разные нейротрансмиттеры, если лишь одного достаточно для того, чтобы опосредовать передачу всех электрических сигналов?» (см. в [27]). Сложность теоретических построений, необходимых для того, чтобы ответить на этот простой вопрос, впечатляет ([27] и другие статьи в том же номере журнала).
С позиций представления о системной детерминации активности нейронов нейротрансмиттер больше не рассматривается как стимул, действующий на нейрон (или отдельный его локус), а нейрон – не рассматривается как передатчик электрических сигналов. Вместо этого нейротрансмиттеры рассматриваются как вещества необходимые для метаболизма клетки, которые вносят вклад в удовлетворение ее «потребностей». Многообразие и сложность этих потребностей таково, что разнообразие медиаторов само по себе не кажется удивительным. Проблема же «множественности нейротрансмиттеров» [27], т.е. множественности передатчиков гомогенных электрических сигналов, превращается при этом в проблему определения специфики метаболических паттернов, связанных с обеспечением тех или иных «потребностей», и в проблему модификации паттернов при формировании специализации нейронов в процессе системогенеза (см. ниже).
Заметим, что в рамках представлений, отрицающих подход к нейрону как к проводнику возбуждения, сам термин «медиатор» или «нейротрансмиттер» (используемый в значении «биологически активное вещество, являющееся посредником в процессе передачи возбуждения через осуществление синаптического влияния») представляется неадекватным.
НАУЧЕНИЕ КАК ОБРАЗОВАНИЕ «СЛЕДОВ» И КАК СИСТЕМОГЕНЕЗ
Системно-селекционная концепция научения
Идея развития, наряду с идеей системности, относится к фундаментальным положениям теории функциональных систем. Обе они были воплощены в концепции системогенеза, которая сформулирована с привлечением обширного экспериментального материала, накопленного при исследовании формирования нервной системы и поведения. В этих исследованиях было обнаружено, что в процессе раннего онтогенеза избирательно и ускоренно созревают именно те элементы организма, имеющие самую разную локализацию, которые необходимы для достижения результатов систем, обеспечивающих выживание организма на самом раннем этапе индивидуального развития [12].
В настоящее время становится общепризнанным, что многие закономерности модификации функциональных и морфологических свойств нейронов, а также регуляции экспрессии генов, лежащие в основе научения у взрослых, сходны с теми, которые определяют процессы созревания, характеризующие ранние этапы онтогенеза [9,10]. Это дает авторам основание рассматривать научение как реактивацию процессов созревания, имеющих место в раннем онтогенезе.
В теории функциональных систем, наряду с признанием специфических характеристик ранних этапов индивидуального развития по сравнению с поздними [3,41], уже довольно давно [32,36] было обосновано представление о том, что системогенез имеет место не только в раннем онтогенезе, но и у взрослых. Формирование нового поведенческого акта в любом возрасте есть формирование новой системы - системогенез.
Позднее был сделан вывод о том, что принципиальным для понимания различий роли отдельных нейронов в обеспечении поведения является учет истории формирования поведения, т.е. истории последовательных системогенезов [3,5], а затем разработана системно-селекционная концепция научения ([147], см. также в [4,6,7,38]).
Системно-селекционной концепции созвучны современные идеи о «функциональной специализации», пришедшие на смену идеям «функциональной локализации», и о селективном (отбор из множества клеток мозга нейронов с определенными свойствами), а не инструктивном (изменение свойств, «инструктирование» клеток соответствующими сигналами) принципе, лежащем в основе формирования нейронных объединений на ранних и поздних стадиях онтогенеза [77]. Представление, сложившееся как в когнитивных науках, так и в биологии, о том, что индивид снабжен «многоцелевыми» врожденными элементарными «кирпичиками» - «примитивами», из которых в условиях инструктирующего влияния среды может быть сконструировано что угодно в процессе «ассоциативного обучения», подвергается разносторонней критике [126]. Так, например, Дж. Эдельманом доказательно обосновываются преимущества представления, согласно которому научение есть процесс селекции и прогрессивной специализации [77].
Принцип селекции по Эдельману может быть сведен к следующим положениям. В мозгу формируются группы нейронов, каждая из которых по-своему активируется при определенных изменениях внешней среды. Специфика группы обусловлена как генетическими, так и эпигенетическими модификациями, происшедшими независимо от упомянутых изменений. Селекция имеет место уже при созревании мозга в раннем онтогенезе, в процессе которого множество нейронов гибнет. Отобранные же клетки составляют первичный ассортимент. Вторичный ассортимент, формируется в результате селекции, происходящей при научении в процессе поведенческого взаимодействия со средой. Принятие положения о селекции как основе развития на всех его этапах устраняет дихотомию между созреванием и научением [134].
В рамках системно-селекционной концепции научения формирование новой системы рассматривается как формирование нового элемента индивидуального опыта в процессе научения. В основе формирования новых функциональных систем при научении лежит селекция нейронов из «резерва» (предположительно низко активных или «молчащих» клеток). Эти нейроны могут быть сопоставлены с первичным ассортиментом и обозначены как преспециализированные клетки.
Селекция нейронов зависит от их индивидуальных свойств, т.е. от особенностей их метаболических «потребностей». Отобранные клетки становятся специализированными относительно вновь формируемой системы. Эти нейроны могут быть сопоставлены с вторичным ассортиментом по Эдельману.
Специализация нейронов относительно вновь формируемых систем — системная специализация — постоянна. (Интересно, что сформированная «память» иммунных клеток также постоянна, они никогда не «забывают» [93].) Таким образом, новая система оказывается «добавкой» к ранее сформированным, «наслаиваясь» на них.
Положения о наличии в мозге животных разных видов большого числа «молчащих» клеток, об увеличении количества активных клеток при обучении, а также о том, что вновь сформированные специализации нейронов остаются постоянными (в эксперименте - в течение всего периода хронической регистрации: недели и даже месяцы), и что при научении происходит скорее вовлечение новых нейронов, чем переобучение старых, согласуются с данными, полученными в работах ряда лабораторий [59,63,99,152,156,160,162].