
Звіт про науково-дослідницьку роботу морфофункціональні особливості перебудови скелета та внутрішніх органів в умовах порушеного гомеостаза
| Вид материала | Документы |
- Реферат звіт про науково-дослідницьку роботу:, 74.48kb.
- Реферат звіт про науково-дослідну роботу, 23.08kb.
- Реферат звіт про науково-дослідну роботу, 29.85kb.
- Реферат звіт про науково-дослідну роботу, 18.54kb.
- Реферат звіт про науково-дослідну роботу заключний, 39.02kb.
- З в І т Миргородського міського голови про роботу міської ради та її виконавчих органів, 765.99kb.
- Ся з метою інтеграції наукової та навчальної діяльності в системі вищої освіти,, 67.58kb.
- На науково-дослідницьку роботу Чупілки, 213.36kb.
- Розділ особливості діяльності органів внутрішніх справ, пов’язані з проблемою незаконного, 866.96kb.
- Міністерство освіти І науки України Кременчуцький державний політехнічний університет, 663.7kb.
2. 3 Дослідження печінки в умовах дії малих доз радіації та солей важких металів.
Аналізуючи результати дослідження впливу іонізуючого випромінювання в малих дозах (0,1 Гр; 0,2 Гр; 0,3 Гр) на печінку тварин, виявлені зміни на органному, клітинному і субклітинному рівнях, ступінь яких залежить від отриманої дози опромінення. Відмічено достовірне збільшення відносної маси печінки в групах R1, R2 і R3 на 19,7%, 26,6% і 34,4% відповідно. Морфометрично встановлено збільшення розмірів печінки: довжини і ширини відповідно групам на 4,7% і 2,3%; 6,4% і 4,0% та 9,6 і 8,8%.
Іонізуюче випромінювання викликає судинні розлади в органі. Відмічено повнокрів’я судин, наявність престазів і стазів, розширення навколокапілярних просторів і синусоїдів. Зі збільшенням дози випромінювання ці розлади поглиблювалися. Визначався набряк судинних стінок, гіперплазія ендотелію, набряк перипортальної сполучної тканини, лімфогістіоцитарна інфільтрація сполучної тканини навколо печінкових судин.
Морфометричні показники печінки, що вивчалися, змінювалися зі збільшенням отриманої тваринами дози. Відповідно до груп піддослідних тварин цієї експериментальної серії R1, R2 і R3 відносний об’єм ушкоджених гепатоцитів склав 22,4%±3,2%; 29,2±2,7% і 43,7±2,4%. Кількість гепатоцитів на 100 п.з. збільшилася на 13,4% і 14,9% у перших двох групах, а у тварин третьої зменшилася на 10%; кількість двоядерних гепатоцитів збільшилася на 11,6%, 21,2% і 15,0% відповідно (табл.1).
Таблиця 1
Дані морфометрії печінки щурів за дії іонізуючого випромінювання
-
Показники
Інтактні щурі 4 міс.
Доза опромінення
0,1Гр
0,2Гр
0,3Гр
Відносна маса печінки (%)
3,24±0,18
3,87±0,2
3,97±0,4
4,35±0,8
Довжина печінки (мм)
50,08±1,9
52,43±1,6*
53,14±2,3*
54,9±2,7*
Ширина печінки (мм)
42,9±1,9
44,40±1,2*
45,29±2,03*
46,7±2,2*
Товщина печінки (мм)
8,30±1,12
8,43±0,9*
8,64±2,07*
9,27±1,8*
Кількість гепатоцитів на 100 п.з
1306,1±39,7
1481,74±18,3
1500,72±12,6
1175,49±14,12
Об’єм ушкоджених гепатоцитів (%)
0
22,4±3,2
29,2±2,7
43,7±2,4
Ядерно-цитоплазматичне відношення (у.о.)
16,6±1,8
17,01±1,13
19,74±2,4
19,98±1,3
Кількість двоядерних гепатоцитів (%)
18,3±2,6
20,42±0,7
22,17±0,8
21,04±1,02
* р>0,05, у всіх інших випадках р<0,05
Зміни гістоструктури печінки тварин знаходилися в лінійній залежності від дози випромінювання. У групах R1 R2 ці зміни були помірні.
При максимальній дозі радіації у наших дослідженнях (0,3 Гр) відмічена дискомплексація печінкових балок, розвивалася жирова і вакуольна дистрофія значної частини гепатоцитів. Ультраструктура гепатоцитів щурів цієї групи набула змін, що переходять межу фізіологічної компенсації. Відмічено різке набухання мітохондрій, зменшення кількості крист, що свідчить про порушення біоенергетики печінкових клітин. Цистерни гранулярного ендоплазматичного ретикулуму розширені, зменшена кількість зв’язаних і вільних рибосом і полісом. Комплекс Гольджі редукований. Такі ж зміни спостерігалися і в ендотеліоцитах. Внутрішньоклітинні дистрофічні процеси з елементами деструкції розвивалися в купферівських клітинах. Відмічені деструкція крист мітохондрій, поява осередків лізису зовнішньої мембрани, розширення цистерн ендоплазматичної сітки, вогнищева деструкція мембран.
Встановлена зміна хімічного складу печінки при опроміненні малими дозами іонізуючого випромінювання. Зі збільшенням терміну дії опромінення і його величини зростає втрата печінкою води на 6,4% у групі R1, на 10,7% - у групі R2 і на 12,5% - у групі R3 (Р0,05). Зневоднення спричиняє виведення із печінки мікроелементів. Достовірно знижується вміст міді відповідно групам R1-R3 на 9,7%, 12,7% і 16,2% (Р0,05); цинку – на 3,5%, 6,9% і 9,8% (Р0,05); марганцю – на 12,5%, 18,7%, 22,3% і хрому – на 4,25%, 7,9%, 11,4% (Р<0,05). Зниження свинцю незначне – на 2,7%, 4,2% і 5,4% відповідно групам. Загальна мінералізація печінки зростає з 29,7% (R1) до 44,3% (R3), що, можливо, пов’язано з відкладанням у міжклітинному просторі кальцію та інших мікроелементів.
Солі важких металів спричиняють у печінці більш суттєві зміни, ніж радіація. Визначалася дискомплексація печінкових балок, яка прогресувала зі збільшенням терміну навантаження до 3-х місяців, в часточках з’являлися дрібні вогнища некрозу. У порівнянні з дією іонізуючого випромінювання поглиблюються дистрофічні процеси в гепатоцитах. Більшість з них з ознаками жирової, зернистої, а в деяких клітинах і балонної дистрофії. Цитоплазма втрачає базофільність, відмічається дискаріоз, багато без’ядерних клітин. Кількість двоядерних гепатоцитів і цитоплазматичного відношення у групах С2 і С3 знизилась на 3,9% і 2,5% та 16% і 21,1% відповідно.
Тривала дія солей важких металів (2-3 міс.) спричиняє значні зміни на субклітинному рівні. Структурно це визначається дрібновогнищевою деструкцією мембран ендоплазматичної сітки, зовнішніх мембран і крист мітохондрій, ядерної мембрани. Це прояви серйозних порушень біоенергетичних процесів у клітині, що спричиняє зниження синтетичної і репараційної активності метаболізму органел (55, 56). Розвиток катаболічних процесів структурно підтверджується редукцією пластинчастого цитоплазматичного комплексу Гольджі і появою в ділянці його локалізації великої кількості вторинних лізосом.
Дослідження хімічного складу печінки визначило підвищення кількості отриманих із питною водою мікроелементів. Кількість неорганічних речовин збільшилася на 58,6%, 64,6% і 71,0%, а вологість зменшилася на 10,9%, 17,1% і 16,3% відповідно групам С1, С2 і С3.
Комбінована дія солей важких металів і опромінення викликають у печінці більш глибокі зміни у порівнянні з дією тільки одного несприятливого чинника, що є наслідком накладання негативної дії двох факторів.
Зміни гістоструктури печінки у піддослідних тварин цих груп залежали від тривалості навантаження солями важких металів. Розвивалися дистрофічні зміни в гепатоцитах від дрібних ліпідних включень у тварин групи R1+С1, помірної зернистої і жирової дистрофії у печінці щурів групи R1+С2 до появи клітин з грубозернистою, вакуолізованою, іноді з ацидофільним забарвленням цитоплазмою, з пікнотичними ядрами, з вогнищами круглоклітинної інфільтрації і некрозу у групі R1+С3. Не- зважаючи на зростання дистрофічних і деструктивних процесів, виявляються виражені адаптаційно-компенсаторні процеси, що вказує на високий регенеративний потенціал печінки. Збільшується кількість гепатоцитів на одиницю площі на 8,4% у групі R1+С1, на 22,5% - у групі R1+С2. Відповідно у цих групах збільшується ядерно-цитоплазматичне відношення на 17,4%, 8,1%, а також кількість двоядерних гепатоцитів.
Із зростанням терміну вживання солей збільшується конденсація хроматину по периферії ядра, змінюються оболонки останнього. Зростає набряк мітохондрій і вакуолізація їх крист. Розширюються цистерни ендоплазматичної сітки. Знижується кількість вільних рибосом, полісом і глікогенових гранул. Комплекс Гольджі помірно редукується. Зі збільшенням терміну вживання солей важких металів до 3 місяців дистрофічні процеси переростають у деструктивні. У деяких гепатоцитах зустрічаються безструктурні зони. Комплекс Гольджі редукується.
При збільшенні дози іонізуючого випромінювання до 0,2 Гр за умови дії солей важких металів в тих же концентраціях і термінах, що і в попередніх групах піддослідних тварин зміни гістоструктури печінки прогресують. Більшість гепатоцитів з ознаками зернистої і жирової дистрофії. Зустрічаються гепатоцити зі спустошеною цитоплазмою і без ядер. Посилюються деструктивні процеси – у деяких часточках утворюються безструктурні зони та поодинокі інфільтрати.
На субклітинному рівні визначалися дистрофічні зміни органел. Зменшувалася їх кількість у цитоплазмі гепатоцитів. Мітохондрії набувають майже округлої форми, зменшується число крист, мембрани мітохондрій частково лізуються. Зростає деструкція ендоплазматичного ретикулуму з утворенням електронно-прозорих вакуолей. Різко знижується вміст глікогену. Кількість рибосом і полісом зменшується.
Отже, комбінація двох екологічних чинників – опромінення і навантаження металами викликає більш значні зміни в печінкових клітинах, ніж вплив одного несприятливого фактора окремо. Із збільшенням дози радіації і терміну споживання сполук важких металів ці зміни зростають і переходять із фази адаптаційно-репараційної у деструктивно-некротичну, про що свідчать результати третьої серії експерименту (R3+С1-С3).
При вивченні гістоструктури органа встановлено розростання сполучної тканини. Зустрічалися ділянки, в яких паренхіма заміщувалася сполучною тканиною. Втрачалося радіальне розташування печінкових пластинок і межі часточок. У більшості гепатоцитів розвивалася зерниста і жирова дистрофія. Контури гепатоцитів втрачали чіткість, цитоплазма у багатьох спустошена. Більшість ядер у стадії каріопікнозу, багато без’ядерних клітин.
Дистрофічні зміни клітинних органел розвивалися швидше і переходили у деструктивну фазу. В гепатоцитах групи R3+С1 значно зменшувалася кількість крист мітохондрій, знижувалася електронна щільність їх матриксу. Цистерни гранулярного ендоплазматичного ретикулуму вакуолізувалися. На його мембранах зменшувалася кількість рибосом і полісом, що свідчить про зниження інтенсивності синтезу білків як для репаративних потреб клітини, так і для потреб організму. В кінці другого місяця експерименту (R3+С2) в гепатоцитах з’являлася дрібновогнищева деструкція внутрішньоклітинних мембран. Поглиблення деструктивних процесів підтверджується редукцією пластинчастого цитоплазматичного комплексу Гольджі та появою у цитоплазмі великої кількості вторинних лізосом і ліпідних включень. В кінці експерименту катоболічні процеси у гепатоцитах переважали над синтетичними, що структурно проявлялося деструкцією значної кількості мітохондрій і їх крист, фрагментацією мембран гранулярного ендоплазматичного ретикулуму і редукцією комплексу Гольджі.
Для цієї експериментальної групи характерний підвищений вміст у печінці тих металів, що надходили в організм з питною водою, але він був нижчий, ніж у щурів другої експериментальної серії. Вміст марганцю, який входить до складу коферменту лужної фосфатази, вищий від контрольної групи на 7,1% (R3+С1), 13,8% (R3+С2) і 18,5% (R3+С3). Кількість цинку, який входить до складу карбоангідрази, дегідропептидази і інших ферментів, що забезпечують вуглеводний обмін, збільшується відповідно на 14,9%, 20,9% і 37,6%. Вміст міді, яка міститься в цитохромі, є збільшеним відповідно на 2,4%, 6,5% і 16,4%. Вміст хрому та свинцю збільшується відповідно на 4,7% і 7,4% (R3+С1), 6,8% та 16,2% (R3+С2) і на 11,5% і 22,7% (R3+С3). Відбувається подальше зниження у печінці води. Всі дані статистично достовірні (Р<0,05). Таким чином, комбінована дія екологічних чинників викликає деструктивні явища в печінці.
2. 4 Дослідження довгих кісток в умовах дії малих доз радіації та солей важких металів.
Аналізуючи комбінований вплив металів, що містяться в воді та ґрунті Ямпільського району (цинк та мідь) в кількості , що перевищують ПДК на 60-80% та низьких доз радіації, які еквівалентні рівню 0,1, 0,2 та 0,3 Гр, помітно поступове порушення росту та формоутворення довгих кісток скелету.
Максимальна довжина кісток зменшується пропорційно дозі випромінення з максимальною різницею через тиждень - відповідно 8,67% (р<0,05), 13,02% (р<0,05) та 16,29% (р<0,01). В подальшому відбувається зменшення різниці з контролем, але в групах тварин, що опромінювалися в дозах 0,2 та 0,3 Гр, вона залишається достовірною протягом всього терміну спостереження. Ширина проксимального та дистального епіфізів також зменшується зі збільшенням дози випромінення з максимальною різницею через тиждень після експерименту, яка відповідно складає 3,56% - 8,0% (р<0,05) та 4,98% - 7,94% (р<0,05) (0,1 Гр), 10,82% - 12,65% (р<0,05) та 9,73% - 12,68% (р<0,01) (0,2 Гр) і 13,29% - 14,39% (р<0,01) та 13,99% - 16,21% (р<0,01) (0,3Гр).
Аналіз поперечних розмірів середини діафізу вказує на протилежну тенденцію. Так через добу відбувається збільшення ширини та передньо-заднього розміру в залежності від дози випромінення відповідно на 5,51% - 8,11% (р<0,05) та 6,01% - 7,42% (р<0,05) (0,1 Гр), на 9,1% - 11,75% (р<0,05) та 9,16% - 10,89% (р<0,01) (0,2 Гр) і на 14,67% - 16,97% (р<0,01)та 13,06% - 14,79% (р<0,01) (0,3 Гр). Через тиждень різниця з контролем збільшується на 1,5 - 2%, та поступово має тенденцію до зменшення, але в групах тварин, що опромінювались в дозах 0,2 та 0,3 Гр не повертається до рівня інтактних тварин.
Гістологічно відбувається поступове порушення як структури ростової пластини так і компактної речовини діафізу.
Так при опромінені в дозі 0,1 Гр структура епіфізарного хряща за В.Г. Ковешніковим не відрізняється від інтактних тварин протягом всього терміну спостереження, але зі збільшенням дози до 0,2 Гр відмічається накопичення сполучної тканини в проліферативній та дефінітивній зонах, зменшується кількість клітин, що формують "монетні стовпчики" неправильної форми зі зруйнованими оболонками та без фігур мітозу. Помітно розширена зона деструкції, що вмішує значну кількість сполучної речовини. Через тиждень відбувається деяке погіршення структури та майже повне її відновлення через місяць спостереження.
В умовах споживання солей цинку та міді і опроміненні в дозі 0,3 Гр важко розрізнити межі зон із-за накопичення сполучної речовини, різкого зменшення кількості хондроцитів та порушення їх орієнтації. Більшість клітин сплющеної форми без фігур мітозу, багато з них вакуолізовані. Через тиждень епіфізарний хрящ повністю втрачає звичну будову, клітини формують конгломерати, що оточені проміжною речовиною. Відновлення хряща відбувається повільно і через місяць після експерименту ростова зона представлена хондроцитами, що формують невиражені стовпчики. Багато з клітин вакуолізовані та напівзруйновані. Межі зон визначаються нечітко, що пов'язане з проникненням клітин в сусідні зони.
Морфометрично відбувається поступове звуження ширини епіфізарного хряща в залежності від дози опромінення, з максимальною різницею через тиждень. Звуження відбувається в основному за рахунок проліферативної та дефінітивної зон, різниця з контролем яких через тиждень складає відповідно 9,94% - 14,72% (р<0,05) та 7,95% - 9,71% (р<0,05) (0,1 Гр), 16,97% - 21,42% (р<0,01) та 12,89% - 16,81% (р<0,05) (0,2 Гр) і 29,54% - 35,27% (р<0,01) та 22,76% - 27,75% (р<0,01) (0,3 Гр). В групі тварин, що опромінювалась в дозі 0,1 Гр, через три тижні ці показники недостовірно відрізняються від контролю, в інших - поступове зменшення різниці не зрівнюється з показниками інтактних тварин.
Зона деструкції реагує на несприятливі чинники поступовим розширенням. В групі R1+С1, достовірна різниця з контролем спостерігається тільки через добу та 1 тиждень. В той же час в групах R2+С1 та R3+С1 вона є достовірною протягом всього терміну спостереження. Зона індиферентного хряща майже не реагує на вплив несприятливих чинників.
Діафіз, як більш стала структура, в групі R1+С1 не реагує на несприятливі чинники Ямпільського району. При зростанні дози радіації до 0,2 Гр з`являються лінії склеювання, мозаїчні ділянки забарвлення. В остеонному шарі відмічається поява первинних остеонів, що достовірно підтверджується зменшенням їх діаметру та збільшенням ширини гаверсових каналів. На межі з ендостальним шаром з`являються лакуни резорбції, що свідчить про порушення перебудови кістки. Зменшується кількість остеобластів, що слабо сприймають барвники. Перфоруючі канали пронизують товщу діафізу, порушуючи його компактність. З третього тижня помітне відновлення морфоструктури діафізу: зникають лінії склеювання, порожнини резорбції, мозаїчні ділянки звапнення зустрічаються на окремих препаратах стегнових кісток.
Найбільші зміни спостерігаються при опроміненні в дозі 0,3 Гр. В діафізі межі зон розрізняються нечітко. Остеонний шар різко звужений і представлений в основному первинними остеонами. Ділянки мозаїчного забарвлення займають всю площину компакти. Порожнини резорбції, заповнені сполучною тканиною, мають значні розміри. Майже не виділяються остеоцити, їх лакуни зливаються, утворюючи неправильних розмірів порожнини. На деяких препаратах помітно заміщення компактної кістки грубоволокнистою, що особливо характерно для великогомілкових кісток. Лінії склеювання, що свідчать про затримку росту кістки, значної товщини. Внутрішні генеральні пластинки мають звивистий характер, місцями відсутні. Остеобласти периостального шару не забарвлюються, фігури мітозів відсутні. Через місяць помітно незначне покрашення будови діафізу. Лінії склеювання майже не змінюються. Зменшуються порожнини резорбції, але ще займають значні площі. Первинні остеони майже не заміщуються вторинними, багато хаотично розміщених уламків. На межі з кістковим мозком залишаються ділянки грубоволокнистої тканини.
Морфометричні показники діафізу також залежать від дози випромінення. Так поступове звуження остеонного шару прямо пропорційне дозі випромінення. В групі R1+С1 достовірна різниця спостерігається в строки 1 день та 1 тиждень. Зі збільшенням дози радіації збільшується різниця з контролем: в групі R3+С1 вона складає 22,19% - 25,62% (р<0,01) через тиждень спостереження, поступово знижується але навіть через місяць дорівнює 7,31% - 10,02% (р<0,05). Розширення зон внутрішніх та зовнішніх генеральних пластинок також сягає значних цифр в групах R2+С1 та R3+С1 та є достовірним протягом всього експерименту.
Зміни хімічного складу кісток характеризуються збільшенням вмісту вологи та гідрофільних елементів калію і натрію, зменшенням мінеральної насиченості кістки, в основному за рахунок кальцію і накопиченням металів, що надходили зовні (цинк та мідь). При чому виражені зміни помітні вже в групі R1+С1 та значно зростають зі збільшенням дози випромінення.
Найбільше страждає рівень кальцію, який одним з перших реагує на ушкодження, що в залежності від групи зменшений через добу на 9,11% - 11,85% (р<0,05) (R1+С1), на 14,99% - 18,51% (р<0,05) (R2+С1) та на 23,88% - 26,85% (р<0,01) (R3+С1). Через тиждень його вміст зменшується ще на 2-3% та поступово має тенденцію до збільшення, але достовірно не відрізняється від контролю тільки в групі R1+С1 через три та чотири тижні після експерименту.
Вміст міді та цинку зростає в групі R1+С1 максимально через тиждень відповідно на 8,49% - 10,45% (р<0,05) та 7,45% - 9,43% (р<0,05), в групі R2+С1 - на 12,83% - 16,37% (р<0,01) та 11,55% - 12,31% (р<0,05), в групі R3+С1 - на 16,83% - 18,99% (р<0,01) та 15,33% - 14,38% (р<0,01). Ці данні підтверджують деякі дослідження, щодо накопичення екзогенних металів в кістках [57, 58], але наші цифри значно вищі, вірогідно із-за додаткового впливу радіації, що сприяє включенню міді та цинку в хімічний склад кістки.
Середино-Будський район характеризується підвищеним вмістом в воді та ґрунті солей марганцю, свинцю та міді та підвищеним рівнем радіації, що в різних місцях еквівалентна дозам 0,1, 0,2 та 0,3 Гр.
Місячний вплив на тварин зазначених солей металів та опромінення в дозі 0,1 Гр викликає затримку росту кісток, яке дещо більша ніж в попередньому районі в середньому на 2-3%. Повернення до показників контролю в даній групі тварин для більшості показників відбувається тільки через місяць спостереження. При збільшенні дози випромінення до 0,2 Гр відбувається подальша затримка росту. Так довжина кістки менше за контроль в першу добу на 12,15% - 13,72% (р<0,05), через тиждень - на 12,96% - 14,96% (р<0,01) та поступово повертається до рівня інтактних тварин, але залишається достовірно меншою навіть через місяць. Інші остеометричні показники також перевищують контроль на 2-4%, ніж при дії солей Ямпільського району.
В групі тварин R3+С2, яку опромінювали в дозі 0,3 Гр, різниця довжини кісток складає в бік зменшення через тиждень - 16,77% - 17,96% (р<0,01), ширина проксимального та дистального епіфізів менша в той же термін на 15,63% - 17,44% (р<0,01) та 14,39% - 17,91% (р<0,01). Ширина та передньо-задній розміри діафізу перевищують контроль максимум на 16,93% - 19,11% (р<0,01) та 15,26% - 18,27% (р<0,01), що більше ніж в серії R3+С1 на 2-3%. Всі перелічені показники даної групи достовірно відрізняються від контролю протягом всього терміну спостереження, хоча мають тенденцію до нормалізації.
Епіфізарний хрящ, що забезпечує ріст кістки в довжину, у даних тварин зазнає змін вже при дозі випромінення у 0,1 Гр. Зона росту має типову будову, але помітно деяке збільшення кількості сполучної тканини в тяжах поміж хондроцитів. В проліферативній зоні з'являються клітини нетипової форми - полігональні та різко сплющені. Зменшується кількість мітозів. Клітини цієї зони дещо меншого розміру. Збільшується кількість сполучної тканини й поміж клітин дефінітивного хряща, що представлений типовими хондроцитами, деякі з яких з пікнотичним ядром. Збільшується кількість клітин зони деструкції, що свідчить про затримку процесу утворення кісткового матриксу. Починаючи з третього тижня відбувається нормалізація структури хряща, зменшується кількість сполучної тканини і атипових клітин і через 4 тижні пластинка росту має будову характерну для інтактних тварин.
При опроміненні тварин в дозі 0,2 Гр епіфізарний хрящ різко потоншується, відмічається нерівність контурів та згладженість меж між зонами. Велика кількість сполучної речовини вдається поміж хондроцитів в колонках та поміж колонками, інколи утворюючи окремі конгломерати клітин. Кількість клітин, що утворюють зону проліферації, різко зменшена, окремі клітини зі зруйнованою оболонкою, фігури мітозу відмічаються тільки в шарах, що прилягають до індиферентного хряща. Відмічається поява вакуолей. Зона дефінітивного хряща звужена, інколи представлена 1-2 рядками клітин. Велика кількість сполучної речовини руйнує стовбчасту будову зони та орієнтацію її колонок. Клітини різко збільшені, неправильної форми, багато з них зі зруйнованими ядрами. Зона деструкції різко розширена, що вказує на порушення процесів кісткоутворення. Великі за розміром клітини зони оточені сполучною речовиною, зустрічається безліч уламків клітин та вогнища крововиливів.
Через тиждень збільшується кількість сполучної речовини, руйнуючи стовпчасту будову зон. Фігури мітозу поодинокі. Збільшується вміст атипових клітин. Дефінітивний хрящ майже не виявляється і зливається з зоною деструкції. В наступні строки (2, 3, 4 тижні) поступово зменшується кількість проміжної речовини, збільшується вміст хондроцитів з фігурами мітозу, але багато з них атипової форми. Зона проліферуючого хряща дещо порушена із-за конгломератів клітин. Дефінітивний хрящ чітко визначається тільки через місяць і вміщує значну кількість зруйнованих клітин.
В групі R3+C2 з боку епіфізарного хряща відмічено прогресивне зменшення кількості клітин, що формують проліферативну зону. Майже зникає межа між дефінітивним та проліферуючим хрящем, клітини здебільшого не формують звичних "монетних" стовпчиків, а розміщені окремими групами, що оточені великою кількістю сполучної речовини. Фігури мітозу поодинокі та зустрічаються на окремих препаратах. Відбувається вростання судин в ростову зону. Клітини проліферуючого хряща набряклі та вакуолізовані, що свідчить про значне порушення їх функціонування. Зустрічається значна кількість без`ядерних та зруйнованих клітин. Зона деструкції значно розширена, з незначним вмістом клітин та збільшенням сполучної речовини. Через тиждень межі зон розрізнити неможливо, значно зменшується кількість клітин, фігури мітозів не визначаються. Відновлення структури хряща розпочинається тільки на третій тиждень. Збільшується кількість клітин, що формують зони хряща, але хондроцити розміщені хаотично і майже не утворюють стовпчики.
Морфометрично відбувається прогресуюче звуження ростової зони за рахунок проліферативного та дефінітивного хрящів, що більш виражене ніж у тварин Ямпільського району. Звуження зон проліферуючого та дефінітивного хряща перевищує цифри попередньої групи тварин на 2-4%. Зона деструкції має тенденцію до розширення залежно від дози випромінення. Максимальна різниця отримана через тиждень після експерименту і дорівнює відповідно 7,96% - 8,23% (р<0,05) (R1+C2), 14,53% - 16,17% (р<0,05) (R2+C2) та 19,48% - 24,30% (р<0,01) (R3+C2). В подальшому різниця з контролем зменшується але в групах R2+C2 та R3+C2 достовірно відрізняється від контролю навіть через місяць.
Для комбінації солей Середино-Будського району є характерним поява змін в компактній речовині діафізу вже при сумісній дії радіації, що еквівалентна дозі в 0,1 Гр, чого не зустрічається у інших дослідженнях. З першої доби спостереження відмічено появу поодиноких ліній склеювання на межі периостальної та остеонної зон. Кількість остеобластів біля періосту зменшена. Звужений остеонний шар зміщений в бік періосту розширеною зоною внутрішніх генеральних пластинок. В остеонному шарі спостерігається поодинокі порожнини остеокластичної резорбції, зменшення забарвлювання ядер остеоцитів, розширення каналів остеонів. Інколи виявляються первинні остеони з широким гаверсовим каналом.
Через тиждень збільшується кількість первинних остеонів, з'являються додаткові лінії склеювання, зменшується кількість остеобластів. Починаючи з другого тижня, спостерігається швидке поліпшення будови компакти, зменшуються, і через три тижні повністю зникають, лінії склеювання; через три тижні первинні остеони змінюються вторинними, хоча їх діаметр дещо менший за контроль. Збільшується і навіть перевищує контроль, кількість секретуючих остеобластів, що вказує на активізацію процесів кісткоутворення.
При опроміненні тварин в дозі 0,2 Гр в діафізі переважають процеси резорбції над кісткоутворенням, помітна затримка периостального росту. Багаточисельні лінії склеювання на межі періосту та остеонного шару зливаються, остеобласти майже не забарвлюються. Октеокластичні порожнини резорбції межують з гладкоклітинними, що заповненні сполучною чи молодою кістковою тканиною. Структура остеонного шару значно порушена в першу чергу звивистими пронизуючими канальцями, зонами мозаїчного забарвлення та уламками вторинних остеонів. Значно збільшується кількість первинних остеонів з тонкими кістковими пластинками та широким каналом, що розташовані в різних площинах. Остеоцитарні лакуни місцями без остеоцитів, зливаються, утворюючи порожнини неправильної форми. Цитоплазма остеоцитів слабо забарвлена. Ендостальний шар розширений з порожнинами резорбції та ділянками остеоіду.
Через тиждень збільшується кількість ліній склеювання, вогнищ резорбції, що виповнені сполучною тканиною. Кількість первинних остеонів значно зростає. Через місяць відбувається поновлення структури компакти. Але залишаються незначні за розмірами порожнини резорбції, лінії склеювання добре виражені. Дещо збільшується кількість остеоцитів та остеобластів, але багато з них незабарвлені, що свідчить про зменшення їх активності.
Найбільші зміни в діафізі помітні при додатковому опроміненні в дозі 0,3 Гр: відмічається збільшення площі грубоволокнистої кісткової тканини на межі з кістковомозковою порожниною, майже відсутні вторинні остеони. Остеонний шар являє собою тонкий прошарок кісткової речовини з порожнинами резорбції, ділянками мозаїчного забарвлення, що зміщена в бік періосту. На деяких препаратах великогомілкових кісток остеонна зона не вирізняється з масиву кісткової речовини. Перфоруючі канали пронизують всю товщу кістки. Зовнішні та внутрішні генеральні пластинки повністю не охоплюють діафіз, мають багаточисельні розриви та місцями майже відсутні.
Через тиждень помітно погіршення будови кістки, про що свідчать зникнення зональності, повна відсутність вторинних остеонів, великі масиви грубоволокнистої кістки. Відновлення структури відбувається повільно, через два та три тижні майже непомітне, що вказує на глибокі порушення функціонування кістки. Лише через чотири тижні відмічається зменшення площин грубоволокнистої тканини. На деяких препаратах плечової кістки виділяються лише поодинокі вторинні остеони, чітко видно межі зон.
Морфометричні показники наглядно відображають зростаючі зміни в компактній речовині діафізу. Так ширина остеонного шару через добу зменшена в зрівнянні з контролем на 10,91% - 12,93% (р<0,05) (R1+C2), на 14,89% - 19,37% (р<0,01) (R2+C2) та на 21,96% - 24,94% (р<0,01) (R3+C2), зростає на 2-3% через тиждень та поступово наближається до рівню інтактних тварин, але через місяць різниця дорівнює 2,24% - 3,22% (р>0,05) (R1+C2), 4,68% - 5,98% (р>0,05) (R2+C2) та 7,95% - 11,66% (р<0,05) (R3+C2). Розширення шарів зовнішніх та внутрішніх генеральних пластинок перевищує показники Ямпільського району на 3-4%, що вказує на більш токсичну дію солей міді, марганцю та свинцю. Поява первинних остеонів характеризується звуженням їх діаметру та розширенням діаметру гаверсових каналів, що також перевищують данні з попередньої групи дослідження.
З боку хімічного складу виявлене подальше збільшення вологості кісток, що перевищує дані попередніх груп на 2-4%. Продовжують зростання рівні калію та натрію, як гідрофільних елементів. Зменшення вмісту мінеральних речовин відбувається за рахунок кальцію, вміст якого відповідно групам менше за контроль через тиждень на 13,76% - 14,51% (р<0,05) (R1+C2), на 18,97% - 22,61% (р<0,01) (R2+C2) та на 28,91% - 30,27% (р<0,01) (R3+C2).
Метали, що в надмірній кількості надходили в організм тварин, накопичуються в кістках і їх рівень залежить від дози випромінення. Так вміст марганцю перевищує контроль в групі R1+C2 в першу добу на 6,48% - 7,93%, (р<0,05) в групі R2+C2 - на 10,15% - 12,42% (р<0,05), в групі R3+C2 - на 11,93% - 15,26% (р<0,01). В подальшому відбувається швидка втрата марганцю кісткою, і через місяць після закінчення експерименту його рівень менше за контроль відповідно на 5,99% - 7,54% (р<0,05) (R1+C2), на 6,24% - 8,9% (р<0,05) (R2+C2) та на 9,35% - 11,32% (р<0,05) (R3+C2). Вміст міді та свинцю зростає пропорційно дозі випромінення максимально через тиждень після закінчення експерименту та поступово зменшується, починаючи з другого тижня, але в групах R2+C2 та R3+C2 різниця з контролем є достовірною і через місяць.
Вивчення комбінованої дії солей цинку, хрому та свинцю, що містяться в Шосткінському районі та низьких доз іонізуючої радіації в дозах 0,1, 0,2 та 0,3 Гр, показало найбільш токсичну дії металів даного регіону.
В групі тварин R1+C3 всі остеометричні показники достовірно відрізняються від контролю та перевищують попередню групу на 3-4%, при чому через чотири тижні різниця з інтактними тваринами є достовірною (за деякими виключеннями). При збільшені дози випромінення до 0,2 Гр, різниця з контролем значно зростає. Так в групі R2+C3 затримка повздовжнього росту стегнових кісток складає в перший день 14,91% (р<0,01), через тиждень - 15,79% (р<0,01) і через місяць залишається меншою за контроль на 6,21% (р<0,05). Ширина проксимального та дистального епіфізів менша за показники інтактних тварин максимально через тиждень - відповідно на 13,95% (р<0,01)та 14,76% (р<0,01), і через місяць становить 6,73% (р<0,05) та 7,08% (р<0,05). Поперечні розміри перевищують попередню групу в середньому на 4-5% і залишаються достовірними протягом всього терміну спостереження. Збільшення радіаційного навантаження до 0,3 Гр. призводить до максимального порушення росту кістки з найбільшою різницею в великогомілкових кістках.
Порушення повздовжнього росту відбувається за рахунок епіфізарного хряща, будова та морфометрія якого мають найбільші зміни під впливом екології Шосткінського району. При опромінення щурів в дозі 0,1 Гр границі ростової зони нерівні, місцями глибоко вдаються в перекладки губчастої речовини епіфіза. Різко збільшується кількість сполучної речовини, що місцями відокремлює хондроцити в ізогенні групи, руйнуючи стовбчасту структуру зони. Кількість мітозів зменшується, збільшується вміст клітин атипової форми (полігональні та сплющені). Кількість клітин, що формують стовпчики зменшується до 13-17. Зона дефінітивного хряща представлена клітинами без фігур мітозу, багато з яких зі зруйнованою оболонкою та дистрофічними змінами в ядрі, багато клітин сплющеної форми, що оточені великою кількістю сполучної речовини. Зона деструкції представлена конгломератом напівзруйнованих клітин зі значним вмістом сполучної речовини. Зменшення клітинного складу хряща, збільшення сполучної речовини через тиждень після експерименту змінюється процесами поступового відновлення ростової зони. Через два та три тижні помітно збільшення клітин проліферуючої зони, деяке зменшення кількості сполучної речовини та атипових хондроцитів. В зоні дефінітивного хряща зменшується кількість зруйнованих клітин та проміжної речовини, поліпшується будова зони деструкції. Через місяць відбувається майже повне відновлення структури хряща, але зустрічаються поодинокі хондроцити сплющеної форми без фігур мітозу, в зоні деструкції - збільшення вмісту проміжної речовини.
Морфометрично підтверджується поступове звуження як загальної ширини хряща, так і окремих зон, за винятком шару деструкції, який значно розширюється. (Рис.3)
Р
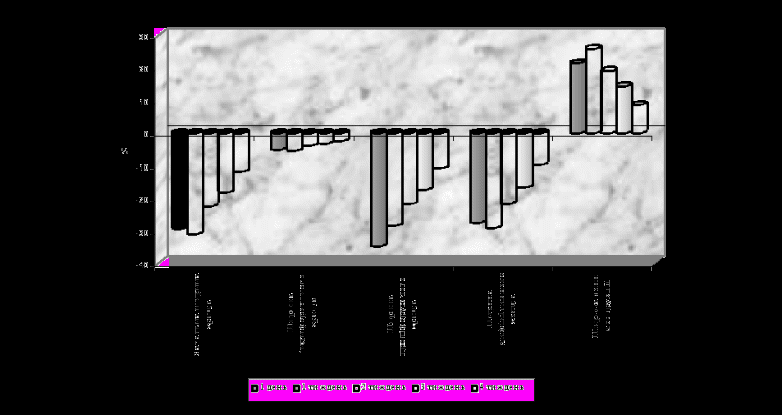 ис.3 Процентне співвідношення показників морфометрії епіфізарного хряща плечових кісток щурів групи R3+C3
ис.3 Процентне співвідношення показників морфометрії епіфізарного хряща плечових кісток щурів групи R3+C3У тварин, яких опромінювали в дозі 0,2 Гр, проліферативний хрящ різко звужений, клітини формують стовпчики, які орієнтовані в різних площинах, різко зменшена кількість хондроцитів. Клітини неправильної (сплющеної та полігональної) форми, багато з них із напівзруйнованим ядром та гігантськими вакуолями. Сполучна речовина відділяє клітини в ізогенні групи та порушує стовпчастість зони. Межа із дефінітивним хрящем майже не розрізняється.
Найбільші зміни помітні в групі R3+С3, що отримувала опромінення в дозі 0,3 Гр. Епіфізарний хрящ характеризується повною втратою зональності, межі зон відсутні, що особливо виражене в великогомілкових кістках. Навіть без морфометрії помітно значне звуження хряща. Складається враження, що ростова зона майже повністю заміщена сполучною речовиною з незначним вмістом клітин, що містяться в її товщі. Хондроцити не утворюють колонок. Клітини сильно набряклі, фігури мітозів не зустрічаються. Уламки хондроцитів зустрічаються по всій площині хряща. Зони уламків хондроцитів значні за розмірами та місцями зливаються.
Діафіз тварин, що зазнали впливу чинників Шосткінського району, з перших термінів спостереження зазнає значних змін. Так при вживанні солей міді, цинку та свинцю і опромінені в дозі 0,1 Гр відмічається збільшення товщини та кількості ліній склеювання в периостальному шарі. Сама зона нерівна, місцями на межі з остеонним шаром помітні ділянки губчастої речовини. Кількість остеобластів зменшена, їх ядра нечітко забарвлені. На межі з остеонним шаром та в його глибині відмічається поява багатоядерних клітин - остеокластів, що утворюють невеликі за розміром порожнини резорбції. Перфоруючі канали збільшуються в розмірі, інколи зливаються, порушуючи структуру остеонного шару, сприяючи тим самим втраті компактності кістки. Збільшується кількість первинних остеонів, що особливо помітно в великогомілковій кістці. Інколи відмічається хаотичне розміщення вставних пластинок. Втрата іонів кальцію та нерівномірність процесів мінералізації призводить до появи мозаїчних ділянок забарвлення. Внутрішні генеральні пластини місцями розірвані. Через тиждень відбувається різке погіршення будови компакти. Відбувається збільшення вогнищ резорбції та мозаїчних ділянок звапнення. Остеоцити місцями не забарвлюються, утворюючи "пустоти" в компакті. Через два тижні помітно зменшення площин резорбції, трохи поновлюється будова оточуючих пластинок. Через місяць відбувається збільшення кількості остеобластів та остеоцитів з чітким забарвленням, порожнини резорбції не відмічаються.
Збільшення дози випромінення до 0,2 Гр призводить до того, що більш розширений ендостальний шар відтісняє до періосту остеонну зону, яка втрачає свою звичну будову. В ній переважають первинні остеони еліпсоподібної форми з широкими каналами, порожнини гладкоклітинної та остеокластичної резорбції займають значну площу компактної речовини. Ділянки мозаїчного забарвлення великого розміру та неправильної форми. На межі периостального та остеонного шару відмічаються лінії склеювання хвилястої форми, що місцями зливаються. Остеобласти та остеоцити майже не забарвлюються, утворюючи видимість порожнин та розривів в компактній речовині. Біля кісткомозкового каналу зустрічається грубоволокниста кісткова тканина.
Але додаткове опромінення щурів в дозі 0,3 Гр та вживання солей Шосткінського району (цинк, хром та свинець) призводить до найбільших змін в компактній речовині. В діафізі домінують процеси резорбції, перебудова компактної речовини не відбувається. Великі за розмірами порожнини остеокластичної резорбції, що виповнені сполучною тканиною, займають всі шари компакти. Межі між зонами нечіткі, звивисті, пластинки розміщені в різних площинах, що руйнує компактність кістки. Між пластинами периостального та ендостального шарів виникають широкі розриви, які місцями виповнені грубоволокнистою кістковою тканиною. Остеонний шар представленим виключно первинними остеонами, що складаються з 2-3 концентрично розміщених кісткових пластинок з широким каналом здебільше овальної форми. Вставних пластинок мало, що вказує на різке уповільнення перебудови кістки. Ділянки мозаїчного забарвлення займають всю товщу компакти. Відновлення будови діафізу не спостерігалось в усі строки спостереження.
Морфометрія показала подальше звуження остеонного шару та розширення зон внутрішніх та зовнішніх генеральних пластинок в зрівнянні з тваринами Середино-Будського району. Різниця становить в середньому 3-6% і залишається достовірною протягом всього терміну спостереження. Збільшення кількості первинних остеонів характеризується звуженням їх діаметру та розширенням гаверсових каналів. Різниця з контролем в великогомілкових кістках через тиждень після закінчення експерименту становить відповідно 11,7% (р<0,01) та 12,92% (р<0,01) (R1+С3); 17,98% (р<0,05)та 18,39% (р<0,01) (R2+С3) і 18,51% (р<0,01) та 21,98% (р<0,01) (R3+С3). Через місяць різниця залишається статистично достовірною.
Зміни хімічного складу кісток тварин, що зазнали впливу чинників Шосткінського регіону, сягають значних цифр. Збільшення вмісту води через тиждень на 13,08% - 16,95% (р<0,01) (R1+С3), на 21,54% - 23,91% (р<0,01) (R2+С3) та на 28,31% - 30,87% (р<0,01) (R3+С3), супроводжується затримкою натрію та калію. Вміст останніх зростає в зрівнянні з показниками Середино-Будського району на 3-5%. Продовжує прогресивно зменшуватися рівень кальцію прямо пропорційно зі збільшенням дози випромінення. Значно зменшується в зрівнянні з контролем вміст міді, магнію та марганцю, досягаючи найбільшої різниці в великогомілкових кістках групи R3+С3 (через тиждень відповідно на 17,98% (р<0,01), 21,74% (р<0,01) та 21,93% (р<0,01)). Метали, що надходили зовні, депонуються в кістковому матриксі. Їх виведення відбувається дуже повільно, і навіть через місяць різниця з контролем є достовірною. Різниця з контролем складає в великогомілковій кістці для цинку - 11,15% (р<0,05), для свинцю - 11,03% (р<0,05).
Таким чином, опромінення в низьких дозах і вживання солей важких металів викликають в кістках скелета остеодепресивні зміни.